两栖动物骨龄学研究进展
2023-02-22李云磊陆宇燕李丕鹏
李云磊,原 涛,陆宇燕 ,李丕鹏
(1.沈阳师范大学两栖爬行动物研究所,沈阳,110034;2.潍坊市寒亭区第一中学,潍坊,261100)
在种群生态学研究中,准确判断种群年龄结构可以为评估种群生存现状、预测种群未来发展趋势、调节和管控野生动物种群提供科学依据,并为制定相应的保护利用和管理计划提供基础数据[1-3]。种群年龄结构是指不同年龄组别的个体数目在整个种群内的比例或配置情况,目前年龄结构特征主要分为增长型、稳定型和衰退型3 种类型[4],表示种群年龄分布特征最直接的方法是金字塔法,其中横轴表示分布的比例,纵轴表示年龄。若种群年龄分布特征呈金字塔型,则表示该种群处于增长状态,属于增长型;若种群年龄分布特征呈倒金字塔型,则表示该种群处于老龄化状态,属于衰退型;若种群年龄分布特征呈钟形,则表示该种群各年龄组分布相对均衡和稳定,属于稳定型。
全球两栖类物种数量呈急剧减少的趋势,甚至有些物种濒临灭绝[5-6]。2012年的统计数据显示,有2 469 种两栖类物种数量逐渐减少[7],自20 世纪70年代以来,保守估计有3.1%的两栖类物种已经灭绝[8],因此,对两栖类物种种群动态监测和分析尤为重要,动态信息的掌握对两栖动物生理、生态,甚至形态分类学的研究都具有重要意义[9]。
1 骨龄学基本概述
1.1 骨龄学方法
个体年龄是种群特征研究中重要的信息之一,但在野外调查和后续分析中,很难直接准确地判断动物个体的年龄。人们逐渐摸索出了借助于一些常规的生物学量度、解剖观察和特定的实验技术等间接推断动物的年龄。两栖类动物代谢功能的强弱会随环境温度的变化而变化[10],这种变化在骨骼生长过程中有明显的印记。通过骨组织或骨板上的特殊生长标记来确定个体年龄的方式已经在一些动物年龄研究中得到了应用,如龟类,通过在龟板上留下类似年轮的特殊标记线来推断龟类的年龄[11-12]。这种利用动物生长在骨组织上的特殊标记线来推断年龄的方法叫作骨龄学方法[13]。
马常夫[14]曾利用“十氢奈泡脊椎骨”的方法对两栖动物的年龄进行探究,将解剖出来的动物脊椎骨去除肌肉,自然干燥后放入十氢奈中处理一定时间后,在显微镜下可观察到椎骨关节凹盘处呈现的黑白相间的年龄环带,以计算个体的年龄。由于凹盘的左右侧壁向外倾斜,其上的环带易被观察到,但也有个体椎骨凹盘上的年龄环带不够清晰,因此对年龄的判断可能出现误差。另外,观察时需要将动物处死,不利于对物种的保护。
目前,在骨龄学方法研究中最为广泛运用并被广大研究者接受和认可的是利用两栖类动物的趾骨或指骨制作石蜡切片[15],HE 染色,观察骨切片上的生长抑制线(也称为年轮线,lines of arrested growth,LAG)数目,进而确定个体年龄。该方法操作简便且得出的结果相对准确[16-18],更重要的是对研究个体损伤有限,不危及生命,但是此法无法确定处于四季交替不明显、没有明显骨骼生长痕迹的动物个体的年龄[19]。
1.2 骨龄学原理
骨龄学方法确定年龄的主要依据是两栖动物生长发育的年周期变化,在环境温度适宜、食物和阳光充足的情况下,机体代谢功能很快得到提升,表现为个体生长(包括骨骼的生长)速度加快,即生长期。在环境温度过低或过高时,两栖类动物主动调节代谢水平,降低到一定水平后进入冬眠或夏眠状态,此时机体的生长速度极度减缓,称为生长的慢速期或停滞期[20]。在这种生长的周期性变化中,两栖动物长骨骨干中的有机物和矿物质的沉积也呈现明显的规律性,即快速生长期,骨质的快速生成导致有机物和矿物质沉积密度较低;而生长停滞期的骨质生长虽然缓慢,但有机物和矿物质沉积却显现出较高的密度。因此,显微观察长骨骨干部切片可清晰分辨出第1 年冬眠期及其前后生长缓慢阶段形成的骨环(染色较深)和第2 年生活期形成的骨环(染色较浅)之间形成的同心环状分界线,这个环状分界线被称为LAG[21],依 据LAG 数目可 间接判 断出个体的年龄。
1.3 骨龄学LAG识别
最早研究幼蛙和成年蛙长骨组织结构的Kastschenko 指出,两栖动物和哺乳动物在长骨组织学结构上存在着明显的差异性[22],两栖动物长骨骨质缺少像哺乳类那样独立的、完整的骨单位[23],会呈现出同心圆状的层层推进的生长方式,这正是骨龄学研究的结构基础。
对LAG 进行识别时,需要对骨骼切片的结构有一定的了解(图1),首先在显微镜下观察长骨切面时,骨髓腔的外侧有一着色较浅的环状骨质结构为内骨板,内骨板离心生长,从骨髓腔处推进式向外生长[25-26],通常在经历了第2 个冬眠期后才可观察到(1龄蛙的长骨组织切片上未观察到内骨板结构),之后每年都会在原有内骨板的外侧形成1 层新的内骨板层[22]。
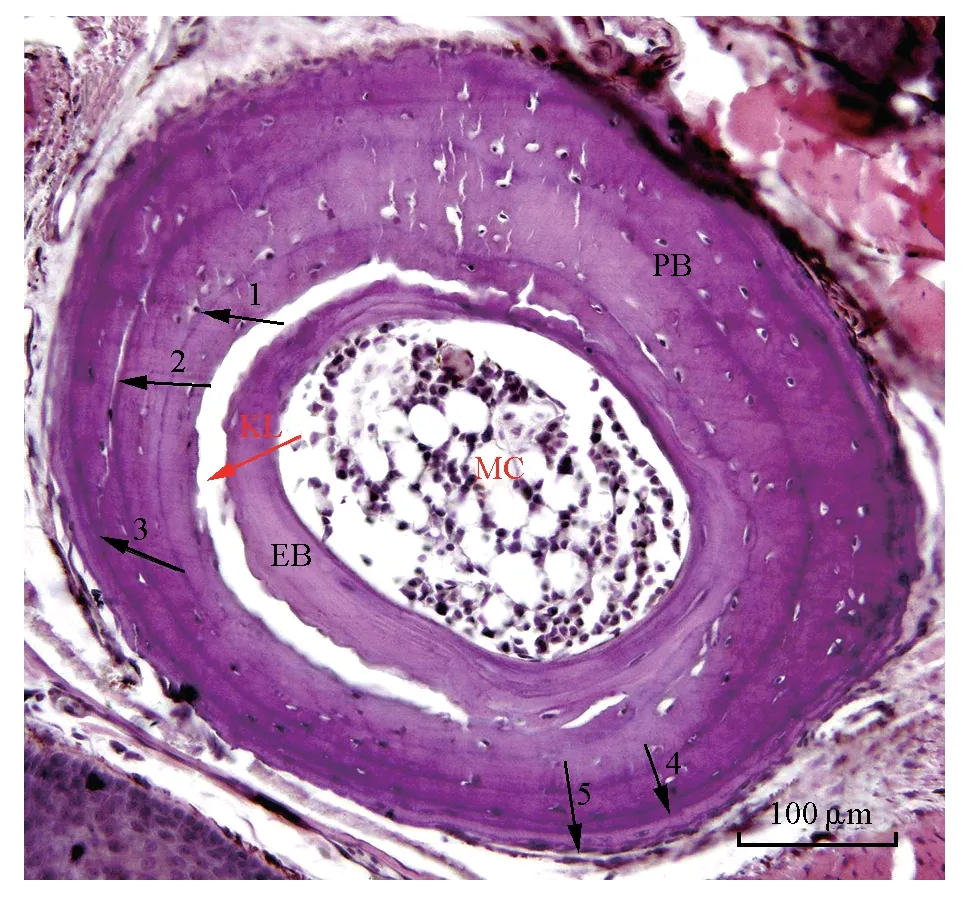
图1 西藏蟾蜍5龄雄性趾骨切片[24]Fig.1 The phalangeal bone of 5th instar male of Bufo tibetanus
在内骨板与外骨板之间有2 条染色特征不同的环状线条,靠近内骨板外侧的1 条着色较深且较粗的线条以及其外侧靠近外骨板内侧着色较浅且较细的线条。前者为重吸收线,又称回复线,其高度矿化,能够被苏木精深染色,同时可作为内骨板与外骨板的分界线,在估计动物的年龄时不可用,即没有年代学价值[27];后者是以首次发现此结构的Kastschenko的名字命名[28],称Kastschenko 线(KL 线),其组织学结构为1 层均质的软骨组织。在观察长骨切片时常可见内、外骨板间出现部分分离的现象,这是由于内、外骨板组织学结构特性不同,在制片过程中化学和物理因素的收缩程度差异导致在KL 线处出现撕裂现象,这种现象并不影响内、外骨板结构的完整性,不妨碍对外骨板上LAG 的计数[29-30]。Kastschenko 认为KL 线的软骨组织结构可能是幼体软骨组织的部分残余,该结构在成体动物骨骼发育中的作用还尚不清楚。也有学者认为KL 与内骨板的形成有关[31]。在KL线附近常常会发生重吸收现象[18],该现象通常发生在近外骨板的区域,有可能会损伤外骨板最内圈的LAG1,致使动物个体年龄有被低估的可能[32-33]。重吸收现象仅出现在最内圈LAG1 局部,并未发现将最内圈LAG1 全部吸收的现象,只要切片质量较好,并仔细观察,对于LAG 计数的影响可以忽略[34]。Castanet 等[35]认为外骨板骨质的重吸收程度与个体的性成熟有关,性成熟越晚重吸收程度越严重,但是对博斯欧螈(Triturus boscai)的研究显示,重吸收的程度与物种的性成熟年龄几乎没有必要的联系,个体1 龄之前生长期的时间长短才是重吸收程度的决定性因素[36]。
在KL 区域内可能存在着与骨质形成有关的成骨细胞和与骨质重吸收有关的破骨细胞。成骨细胞作用于内骨板的离心生长;破骨细胞则作用于外骨板内侧的重吸收。在最外层以骨髓腔为中心呈现辐射状生长的为外骨板,它的生长使长骨骨壁逐年变粗[22]。外骨板由致密的骨组织构成,显微观察长骨横切片时,可以清晰地看到外骨板上经HE染色后同心排列的LAG[37],它是鉴定年龄的唯一标记。确定KL 后,可以辅助确定LAG1 的位置[38],从第1 条LAG开始到外骨板最外缘所有的LAG 数目,即是实验动物的年龄[39]。
1.4 骨龄学LAG提示信息
在观察中发现,每相邻2 条LAG 之间骨组织的颜色与宽度均不同[40],宽度反映了两栖动物对应年份的生长情况。2 条LAG 之间距离较宽,提示对应年份环境条件(温度、食物等)良好,适宜个体生长,骨质积累状态良好。而环境条件相对较差时,个体的生长速度明显降低,2 条抑制线之间的宽度就较窄[41]。研究证明两栖类个体在进入繁殖期前有一个限制性条件,即个体必须到达本物种的最小体长[42],完成变态发育后,均有几年的快速生长期,因而在高海拔寒冷区域生活的两栖类较温暖区域的两栖类物种需要更长的时间达到最小繁殖体长[43],这种现象在长骨切面有良好的印记,完成变态发育后1~4 年(不同物种有所不同)主要为营养生长期,此时2 条LAG 线间的距离较宽。一旦观察到2 条LAG 线间的宽度明显小于其内侧2 条LAG 线间的宽度时,则预示着此个体达到了性成熟年龄[41]。究其原因,是由于两栖类动物个体在营养生长和生殖生长之间的能量投入发生了显著变化,在前期为了尽早达到性成熟的最小体长,两栖类动物将大部分能量用于生长和发育,在达到目标后转而将更多的能量投入到生殖腺及其相关器官的结构和功能的构建和完善中,为进入繁殖期做好充分的准备[44-45]。当然也存在一些特殊情况,如个体达到性成熟后,来年的环境条件优渥,机体在保证生殖投入的前提下,仍能保持较好的生长速度,就会出现性成熟后1 年的2 条LAG 线间宽度大于性成熟当年所形成的LAG 线宽度的情况[46]。
2 骨龄学方法的运用
对两栖动物骨龄学的研究可以追溯到20 世纪40 年代,两栖动物第1 次骨龄学技术的成功尝试是以颅骨为试验材料完成的[13],1940—1968 年,有学者利用不同的骨块作为试验材料探究了两栖动物的年龄问题。Senning[13]采用副蝶骨测定了泥螈(Necturus maculosus)的个体年龄,Schroeder 等[47]采用翼状骨对牛蛙(Lithobatus catesbeianus)的年龄进行分析和研究,在后续的研究中分别有学者利用肱骨、股骨、胫腓骨和椎骨等作为试验材料探究动物的年龄[48],直到Smirina[18]的试验证明动物趾/指骨同样能良好地呈现出LAG 的结构特点,指出该试验的取材方式可以更好地避免对动物个体造成破坏性伤害,此后利用趾/指骨测定动物年龄的方法逐步被认可并广泛使用。
2003 年,Pancharatna 等[48]对印度大头蛙(Limnonectes limnocharis)的肱骨、股骨、胫腓骨、桡尺骨和趾骨5 种长骨的LAG 对比分析,结果显示趾/指骨的LAG 数量基本与其他长骨的LAG 数量保持一致,进一步证明趾/指骨作为两栖动物骨龄学研究材料的可行性。2008年,美国内政部(U.S.Department of the Interior)和美国地质调查局(U.S.Geological Survey)组织专家编写使用骨龄学方法研究无尾两栖动物年龄的方法指南[49],旨在对骨龄学试验技术方法建立一个标准,促进研究工作的规范化。陈贵英等[50]在2008 年提出,根据骨龄学方法利用动物指骨或趾骨除可鉴定动物年龄外,还可兼顾种群生态学研究中的个体标记和分子生物学研究中的组织取样,在必要的取材需求下,尽量降低对个体的创伤以达到对两栖动物资源的保护。对近年来的研究结果统计,显示现存的8 146种两栖动物中,只有266种(3.3%)进行了骨龄学研究,其中蝾螈56 种、无尾两栖类208种,温带物种多于热带物种[51]。由此可见,两栖类种群的年龄结构信息仍存在严重不足,对全面掌握两栖类种群现状、发展信息及物种保护极为不利。
3 骨龄学LAG计数
骨龄学方法虽然快速、简便,但在年龄的确定过程中依旧容易出现误差,需要掌握形态学方面的知识来避免年龄鉴定过程中出现的错误。
3.1 选取观察部位
Rozenblut 等[22]通过对3 种欧洲水蛙(Rana lessonae,R.ridibunda,R.esculenta)长骨分化与发育的研究,发现在蝌蚪40~44 期时,软骨原始细胞最外层的细胞分化成外骨膜,开始逐渐形成外骨板(外骨膜骨);从41 期开始,长骨骨干中央肥大的软骨细胞退化,骨髓腔逐渐形成;到46 期,骨髓腔向两端扩大至靠近骨骺处,最终形成1 个沙漏型的长管状。因此在做长骨切片观察LAG 时,首选长骨近中部髓腔较小的部位切片,以便观察到较为完整的外骨板结构,为准确计数LAG 提供保障[22]。尤其在长骨骨干中部,长度为整块骨长度的25%~35%处,可得到LAG的最大数值[52]。
3.2 附加线
附加线即所谓的假线,假线的形成是由于个体受伤或遇到短暂的气候剧烈变化或食物缺乏而在长骨切面上的反映,一般假线仅在局部出现,未见其形成闭合环状结构,且线条结构不清晰[53]。它虽然代表动物在某个生长期的停滞生长却不代表年周期的生长停滞[54],计数时很容易与LAG区分。
3.3 双重线
有些两栖动物以夏眠的方式度过酷热缺水的夏季,此时在长骨切面上将出现2 条清晰且非常靠近的类似LAG 的结构,称之为“双重线”[54]。在计数时,通常把这种情况记为1条LAG。
3.4 外侧LAG判断
在观察一些老年个体的LAG 时,由于生长缓慢致使2 个年周期的LAG 距离极为接近,若同时出现假线将更加难以区分[39],此时需要有高质量的组织学切片,在高倍镜下还可较清晰地辨认每条LAG,对计数结果影响不大[55]。如果是在生长期后期或冬眠前期捕获的个体,可观察到在外骨板的最外侧有一浅染的骨质环,此环是当年生长期产生的骨质,由于没有冬眠期产生的致密骨质的“衬托”,其尚未形成清晰的LAG,因此,计数骨龄时需要将其计算在内,若忽略此环则会低估个体年龄。
4 结论
出于对两栖动物保护的迫切需要,骨龄学领域的研究越来越活跃,除了对生长停滞具有明显周期性变化的温带两栖动物,如大蟾蜍(Bufo bufo)[56]、美洲林蛙(Rana sylvatica)[57]和欧洲林蛙(R.temporaria)[42]等开展研究外,对代谢功能变化不明显的热带和亚热带两栖动物年龄的研究也逐渐增多[58-59],一部分研究表明骨龄学方法可以应用于没有明显四季变化的热带和亚热带物种,但也有一部分热带两栖动物骨骼没有明显的生长痕迹。在未来的研究中,挖掘出更多骨龄学研究信息,是值得期待和努力的。
