干旱胁迫下腐植酸对燕麦光合特性和内源激素的影响
2023-02-20朱珊珊米俊珍赵宝平武俊英刘景辉
朱珊珊,米俊珍,赵宝平,武俊英,王 莹,刘景辉
(内蒙古农业大学 农学院,内蒙古 呼和浩特 010019)
燕麦是我国北方重要的粮饲兼用作物,具有丰富的营养价值和食用价值,消费市场需求量较大,且呈现逐年增加趋势[1-3]。我国燕麦主要分布在干旱半干旱地区,这些地区自然条件恶劣,在燕麦生育期间干旱少雨,导致燕麦产量低下,生产力远不能满足消费需求[4]。干旱胁迫会导致植物正常生理代谢过程受到抑制,引起植物叶片气孔导度和蒸腾速率降低,导致植物减少蒸腾,以改善自身水分状态[5]。内源激素作为调控作物生长发育的物质在干旱条件下起着非常重要的作用[6]。植物受到干旱胁迫时,体内的生长素(IAA)、赤霉素(GA)和 细胞分裂素(CTK)含量会降低,而脱落酸(ABA)含量会升高,从而调控生理生化过程,以适应干旱[7]。
腐植酸(HA)是一类从泥碳、褐煤和风化煤中提取的能促进植物生长、增强作物抗逆性的有机物质,在农业生产中具有较大的开发和应用前景[8]。有研究指出,干旱胁迫下HA能改善作物叶片光合性能、控制其激素水平、增强其抗旱能力,从而提高作物产量[9-10]。在干旱条件下,HA能够抑制作物叶片气孔开度、增加气体交换速率、降低蒸腾速率,从而改善作物光合能力,增强抗旱性[11-12]。重度干旱胁迫下,喷施HA可调控燕麦叶片内源激素含量,缓解干旱胁迫伤害[13]。Yang等[14]研究显示,在水分亏缺条件下,施用黄腐酸可增加玉米叶片脱落酸含量,降低叶片蒸腾速率,从而提高瞬时水分利用效率,增强植株抗旱性。尽管腐植酸在燕麦抗旱性方面已经有一些研究,但是关于腐植酸对干旱胁迫下燕麦内源激素水平及其与光合特性关系的研究还比较少见。因此,本试验研究了干旱胁迫下腐植酸对燕麦生长指标、叶片光合参数和内源激素含量的影响,并探究了光合参数与内源激素含量间的关系,以期进一步明确腐植酸提高燕麦抗旱性的作用机制,为腐植酸的应用及燕麦的抗旱种植提供理论指导。
1 材料与方法
1.1 试验材料
试验材料为‘燕科2号’,由内蒙古农牧业科学院提供。腐植酸,购于内蒙古永业农丰生物技术有限责任公司。
1.2 试验设计
盆栽试验于2019年4月至8月在内蒙古农业大学燕麦产业研究中心温室进行。试验选用54个直径为25 cm、高18 cm的塑料盆,内装混合土5 kg(河沙和泥炭土体积比1∶1),混合土中N+P+K>10 g/kg,有机质含量>50 g/kg,pH值为5.5~8.5,含水量为9%,播种前施入磷酸二铵(N 18%,P2O546%)2 g/盆。塑料盆采用随机区组排列,设置3个水分条件:正常供水(田间持水量的75%,M1)、中度干旱胁迫(田间持水量的45%,M2)、重度干旱胁迫(田间持水量的30%,M3);2个喷施处理:喷施腐植酸500倍稀释液(HA)和等量清水(CK)。每个处理种植3盆,每盆30株,重复3次,三叶期定植为20株。2019年4月14日播种,8月21日收获。从拔节期(5月18日)开始至成熟期,采用称重差值法进行控水处理,在拔节期、抽穗期、灌浆期的上午09:00-11:00喷施腐植酸500倍稀释液和清水,植株叶片均匀喷施至叶面湿透但不滴水为止,各时期喷施7 d后取燕麦旗叶,投入液氮速冻后,放入-80 ℃冰箱冷藏,用于内源激素含量的测定。
1.3 测定项目及方法
1.3.1 生长指标测定 在拔节期、抽穗期、灌浆期,固定10株燕麦,用卷尺测量株高及每片叶的叶长、叶宽。采用长宽系数法[15]计算每片叶的面积(叶面积=叶长×叶宽×0.73),然后计算单株叶面积。
1.3.2 光合参数的测定 在拔节期、抽穗期、灌浆期,用便携式光合仪(CIRAS-3)在晴朗无风的上午09:00-11:30,测定旗叶气体交换参数,包括净光合速率(Pn)、蒸腾速率(Tr)、胞间CO2浓度(Ci)、气孔导度(Gs)。每个处理测定3株,结果取平均值。
1.3.3 内源激素含量的测定 燕麦旗叶ABA、玉米素核苷(ZR)、IAA、GA含量由中国农业大学作物化学控制研究中心采用酶联免疫吸附法(ELISA)测定。每个处理3次重复,结果取平均值。
1.4 数据统计分析
试验数据采用Microsoft Excel 2010进行处理,用SPSS 22.0软件进行显著性和相关性分析。
2 结果与分析
2.1 腐植酸对干旱胁迫下燕麦生长的影响
由图1可见,随着生育期的推移,燕麦的株高和单株叶面积呈上升趋势,而干旱胁迫导致燕麦株高、单株叶面积均显著降低。与CK相比,M1水分条件下,喷施腐植酸后燕麦的株高和单株叶面积均有所增加,但差异未达到显著水平;M2水分条件下,喷施腐植酸后燕麦株高在抽穗期和灌浆期差异达到显著水平,分别增加了8.40%和7.15%,单株叶面积在拔节期、抽穗期和灌浆期均显著提高,增幅分别为12.78%,6.96%和6.67%;M3水分条件下,喷施腐植酸后燕麦拔节期、抽穗期、灌浆期的株高分别显著增加了9.39%,9.49%和10.27%;单株叶面积分别显著增加了13.05%,8.79%和8.28%。以上结果表明,干旱会抑制燕麦的生长发育,喷施腐植酸可有效缓解干旱胁迫对燕麦生长发育的影响,尤其在M3水分条件下效果更佳。
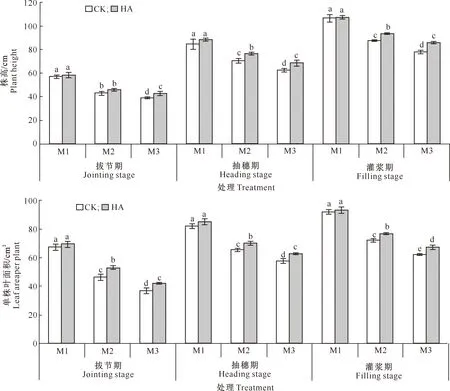
图柱上标不同小写字母表示同一生育时期的不同处理间差异显著(P<0.05)
2.2 腐植酸对干旱胁迫下燕麦旗叶光合特性的影响
由表1可以看出,燕麦旗叶Pn、Ci、Gs和Tr均随着生育期的推进呈先升高后降低的趋势。干旱胁迫导致燕麦旗叶Pn、Gs、Tr显著降低,Ci显著升高。M1水分条件下,喷施腐植酸后燕麦旗叶光合参数与CK处理相比无显著差异;M2水分条件下,喷施腐植酸后3个生育时期燕麦旗叶Pn、Gs和Tr平均较CK分别增加了14.15%,17.65%和13.73%,而Ci平均较CK降低8.19%,其中Pn在拔节期、灌浆期差异达到显著水平,Gs在3个生育时期差异均达到显著水平,Tr在灌浆期差异达到显著水平,Ci在拔节期、抽穗期差异达显著水平;M3水分条件下,腐植酸处理Pn、Gs和Tr平均较CK分别增加了19.15%,20.75%和19.69%,而Ci平均较CK降低了10.89%,其中Pn、Gs和Ci在3个生育时期差异均达显著水平,Tr在灌浆期差异达显著水平。以上结果表明,干旱胁迫给燕麦叶片光合作用带来了不利的影响,喷施腐植酸可有效缓解干旱胁迫对燕麦光合和蒸腾作用的抑制效应,尤其在M3水分条件下效果更明显。

表1 干旱胁迫下腐植酸对燕麦叶片光合参数的影响
2.3 腐植酸对干旱胁迫下燕麦叶片内源激素含量的影响
由表2可见,从拔节期到灌浆期,燕麦旗叶IAA、ZR、GA含量均先升高后降低,而ABA含量则呈上升趋势。干旱胁迫导致燕麦旗叶IAA、ZR、GA含量显著降低,ABA含量显著升高。M1水分条件下,喷施腐植酸后燕麦叶片中仅ZR含量在拔节期较CK显著降低,其他激素含量无显著变化。M2水分条件下,喷施腐植酸后3个生育时期燕麦旗叶ABA含量平均较CK降低了6.59%,而GA、ZR和IAA含量平均较CK分别增加了7.27%,11.68%和10.03%,GA含量在不同生育时期达到显著水平,ABA含量在抽穗期达到显著水平,IAA含量在拔节、灌浆期达到显著水平,ZR含量在抽穗、灌浆期达到显著水平。M3水分条件下,腐植酸处理ABA含量平均较CK显著降低了10.72%,而GA、ZR和IAA含量分别显著增加了11.10%,17.26%和14.99%。结果表明,喷施腐植酸可改变干旱胁迫对燕麦叶片中内源激素含量的影响,有利于燕麦植株维持较高的代谢活力,尤其在M3水分条件下效果更佳。

表2 干旱胁迫下腐植酸对燕麦叶片内源激素含量的影响

表2(续) Continued table 2 ng/g
2.4 内源激素与光合作用及生长指标的相关性
由表3可以看出,Pn、Tr与ZR、IAA和GA含量均极显著正相关,与ABA含量极显著负相关。Ci与IAA含量极显著负相关。Gs与ZR和GA含量极显著正相关,与IAA含量显著正相关,与ABA含量极显著负相关。株高与Pn和IAA含量极显著正相关,与ABA含量、Ci极显著负相关。单株叶面积与Pn、Tr、Gs和IAA含量极显著正相关,与ABA含量、Ci极显著负相关。

表3 燕麦叶片内源激素含量与光合特性及生长指标的相关性
3 讨 论
本研究结果表明,干旱胁迫下燕麦叶片的Pn、Gs、Tr降低,而Ci升高。这可能是由于在干旱条件下,燕麦叶片通过部分或全部关闭气孔降低蒸腾速率,减少水分散失,导致净光合速率下降[16]。此外,Pn的降低伴随着Gs显著降低和Ci增加,表明干旱处理诱导Pn下降的主要原因是干旱条件下燕麦叶片叶绿体结构被破坏,导致叶片光合器官受损,光合活性降低[17]。王琦等[18]研究表明,在低灌水量下喷施腐植酸能显著提高燕麦的Pn、Gs、Tr。本研究中,喷施腐植酸使干旱胁迫下燕麦叶片的Pn、Gs、Tr显著增加,说明腐植酸可能对光合系统有保护作用。周海涛等[19]研究发现,干旱条件下黄腐酸能提高皮燕麦Ci,而本研究结果表明干旱胁迫下喷施腐植酸使燕麦叶片Ci显著降低,这可能是试验所用燕麦品种及光合参数测定时间与方法不同引起的。
本研究中,干旱胁迫下燕麦叶片的IAA、GA和ZR含量均显著降低,而ABA含量显著上升,这表明干旱胁迫会打破燕麦叶片内源激素平衡,降低细胞代谢强度,减少叶片含水量,促进脱落与休眠,从而抑制植物生长[20-21]。张志芬等[13]研究表明,干旱胁迫下喷施腐植酸处理燕麦叶片ABA含量较CK显著降低,但在正常供水条件下差异不显著,本研究结果与此相似,说明腐植酸可通过降低ABA含量来抵御干旱胁迫,延缓植株衰老。Yang等[14]研究表明,在干旱条件下施用黄腐酸能增加叶片ABA含量,提高玉米植株抗逆性,这与本研究结果不同,其原因可能是2种作物受到干旱胁迫时ABA应答基因的调控因子和信号传递途径存在差异,也可能是不同种类作物ABA蛋白在植物细胞的失水耐受过程中所起的作用不同[22]。本试验结果显示,干旱胁迫下喷施腐植酸可以显著提高燕麦叶片IAA、GA和ZR含量,这可能是由于腐植酸中含有吲哚乙酸类小分子生物活性激素分子,这些物质能促进内源IAA、GA和ZR的合成,导致其含量增加[23-24]。
本研究结果表明,干旱胁迫下喷施腐植酸能够显著提高燕麦株高和叶面积,这与申洁[25]在谷子上的研究结果相似。杨建昌等[26]研究认为,Gs与叶片中ABA含量呈极显著负相关,与本研究结果一致,说明在干旱条件下ABA调节了气孔开闭状态。本研究发现,燕麦叶片ABA含量,与Pn、Gs和Tr极显著负相关,这可能是ABA增加导致气孔关闭,Gs和Tr降低,最终使Pn下降。喷施腐植酸减轻了燕麦叶片的损害程度,降低了ABA含量,调控了气孔开闭状态,使蒸腾速率增加,叶片净光合速率提高[27]。前人研究表明,细胞分裂素能促进Rubisco小亚基编码基因表达[28]。本研究结果显示,干旱胁迫下喷施腐植酸能增加ZR含量,且ZR含量与Pn显著正相关,推测干旱胁迫下腐植酸增加了ZR含量,进而提高了Rubisco酶活性,促进了Pn。Cornic等[29]研究认为,IAA可减弱ABA引起的气孔关闭,增加气孔开度,其含量下降有利于气孔关闭。此外,Wang等[30]研究发现,叶片气孔的关闭和蒸腾的减弱是ABA含量增加和ZR含量下降共同作用的结果。本研究中,ZR、IAA、GA含量与Gs存在正相关关系,ABA含量与Gs极显著负相关,高浓度的ZR和IAA可以维持气孔的开张,GA能够促进作物胚轴表皮细胞分裂,促进气孔的形成,调节气孔密度[20,31]。据此,干旱条件下喷施腐植酸对燕麦叶片Gs的影响应该是4种激素共同作用的结果。本研究发现,IAA含量与Pn、Gs、Tr显著正相关,与Ci显著负相关,推测这可能与腐植酸中的吲哚乙酸类小分子生物活性激素分子促进IAA的产生,调控气孔开闭有关[32]。干旱胁迫下GA含量与IAA含量呈显著正相关的关系[6],因此可认为IAA和GA有关,与光合途径相关,但关于GA和光合途径之间的关系还有待进一步研究。
4 结 论
干旱胁迫通过改变叶片的光合作用与内源激素含量影响作物的生长发育。叶面喷施腐植酸可调节植物内源激素的代谢水平,调控燕麦叶片气孔开闭状态,并影响其他光合生理过程,维持燕麦正常的生理代谢活动,促进其生长发育,增强其抗旱能力,当土壤水分为田间持水量的30%时喷施腐植酸的效果最优。
