蕾期低温胁迫对棉花光合特性的影响
2022-08-10钟信念梁其干李有忠赵建华马海霞李志博
钟信念,梁其干,李有忠,3,赵建华,马海霞,李志博
(1.石河子大学农学院/新疆兵团绿洲生态农业重点实验室;2.新疆农业科学院/海南三亚农作物育种试验中心,海南三亚 572014;3.新疆农垦科学院生物技术研究所/作物种质创新与基因资源利用兵团重点实验室,新疆石河子 8320032)
0 引 言
1 材料与方法
1.1 材 料
供试材料为3个新疆北疆不同年代北疆主栽陆地棉棉花品种新陆早10号(1999年)、新陆早36号(2007年)、新陆早61号(2013年),棉花种子均由新疆兵团绿洲生态农业重点实验室棉花育种室提供。
1.2 方 法
1.2.1 棉苗培养及处理
试验于2019年11月至2021年1月于新疆石河子市石河子大学绿洲生态实验室内购置的浙江宁波仪器厂人工智能气候箱中进行。选用大小相同的新发芽盒,3个品种各挑选300粒饱满健康且已经经过硫酸脱绒洗净的新陆早10号、新陆早36号、新陆早61号棉籽作为材料,用0.4%的高锰酸钾溶液对种子进行消毒,消毒时间为10~15 min,然后用蒸馏水560将表面残留的高锰酸钾溶液冲洗3~4次,冲洗后放置于经75%酒精消毒的烧杯中备用。放置于垫有干净滤纸(防止种子根部穿过纱布网眼)及纱布的发芽盒中,用56℃温水泡种20~25 min(加快萌发),每个发芽盒放40粒种子置于加有足以润湿发芽床的少量水,放入28℃培养箱培养,观察种子发芽长势,当芽长1 cm时取出种入花盆。
采用室内人工气候箱盆栽法培养棉花幼苗,选取320x250 mm规格的花盆进行播种育苗。培养土采用花卉市场采购的花土与蛭石以4∶1的比例进行混配后作为基土栽培种植。每盆保苗4株,株距10 cm,各温度每个品种设置4盆作为重复(3盆处理、1盆对照)。出苗后每盆保留4棵长势一致的棉苗继续生长,每3 d以适宜的水肥浇灌于托盘从底部渗透进基土以防止出现土壤板结等问题及便于植株吸收,同时适宜的蹲苗防止出现弱苗、高脚苗、根扎不深等栽培问题。实时观察及时进行虫害管理。图1

图1 实验培养箱
前期培养光照强度设置为50 000xl、昼夜温度为28/20℃、光周期16 h、湿度为60%;待实验材料长至初始现蕾期时,将长势一致的实验材料放入人工智能气候箱内低温处理培养。 试验条件湿度均设置为60%,低温条件分别设置为①光照强度20 000 lx(光照强度根据低温强度设置设置相应的弱光进行调节)昼夜温度20℃/15℃(白天20℃晚上15℃)光周期16 h、②光照强度15 000 lx昼夜温度15℃/10℃光周期16 h、③光照强度10 000 lx昼夜温度10℃/5℃光周期16 h三个低温梯度处理。
对照组设置光照强度为50 000 lx、昼夜温度28℃/20℃光周期16 h进行处理,于实验材料现蕾期开始给予低温胁迫至初花期,期间每处理5d后选取棉花倒4功能叶进行光合及叶绿素荧光参数的测定。去掉一个最高和最低测定值,取其余的平均值作为相应的测定值。
1.2.2 指标测量
使用美国LI-COR 公司便携式光合仪 LI-6400XT测定植物的净光合速率(Pn)、气孔导度(Gs)、胞间 CO2浓度(Ci)和蒸腾速率(Tr),测定光强为 1 000 μmol/(m2·s),CO2浓度为400 μmol/mol,叶室温度为 28℃,流速为 500 μmol/s。选取蕾期棉花倒四片功能叶,并做标记,测定时避开叶脉。之后每次测定使用相同叶片,每枚叶片测定2次,每个处理6 株重复。
Nguyen(2012)发现当首席执行官和董事属于同一社会网络时,CEO不太可能因为业绩不佳而被解雇,董事会成员和CEO之间密切的社会关系会影响董事会职能的发挥;Chahine et al.(2014)发现只有在控制董事会成员与高管的社交联系后,董事会独立性才能发挥效应。那么关于社会关系的度量又该如何展开呢?国外大部分学者的研究集中在工作经历、毕业院校、老乡、专业方面 (Kramarzy et al.2007、 Hwang et al.2009、Liu 2010、Nguyen 2011、Hoitash 2011、Fracassi et al.2012等)。
1.3 数据处理
使用SPSS 26.0软件处理本试验数据统计分析,P<0.05表示差异显著,应用Excel 2019、origin2021等画图软件综合分析作图。
2 结果与分析
2.1 蕾期低温胁迫对净光合速率(Pn)的影响
研究表明,随着处理温度的降低,不同品种的净光合速率均呈显著下降趋势,且温度越低则降幅越大;随低温胁迫天数的增加,不同品种的净光合速率显著下降,胁迫时间越长Pn值越小。T1(20℃/15℃)处理下,各品种相对于对照,Pn缓慢下降。T2(15℃/10℃)低温下Pn显著低于对照和T1(20℃/15℃),T2(15℃/10℃)处理 10 d的Pn大幅度下降,极显著低于处理0 d、5 d;T3(10℃/5℃)的Pn显著低于T1(20℃/15℃)、T2(15℃/10℃)且随胁迫时间的延长Pn值越来越小。
当低温胁迫温度高于20℃/15℃时新陆早61号的净光合速率(Pn)下降的幅度较新陆早36号、新陆早10号趋势小,T1(20℃/15℃)处理下,同一时间3个蕾期棉花品种叶片的Pn值由大到小依次为新陆早61号>新陆早36号>新陆早10号>。当低温胁迫温度低于15℃/10℃时新陆早36号相对于新陆早10号、新陆早61号下降的幅度较小。T2(15℃/10℃)、T3(10℃/5℃)处理下,同一时间3个蕾期棉花品种叶片的Pn值由大到小依次为新陆早36号>新陆早61号>新陆早10号>,中度、重度低温胁迫下新陆早36号相对其它2个品种更耐低温。 同一个低温下,随着低温胁迫时间的延长,净光合速率(Pn)下降加剧;且同一个处理时间段内(15 d),低温胁迫强度越大,净光合速率(Pn)受到的影响越大。图2
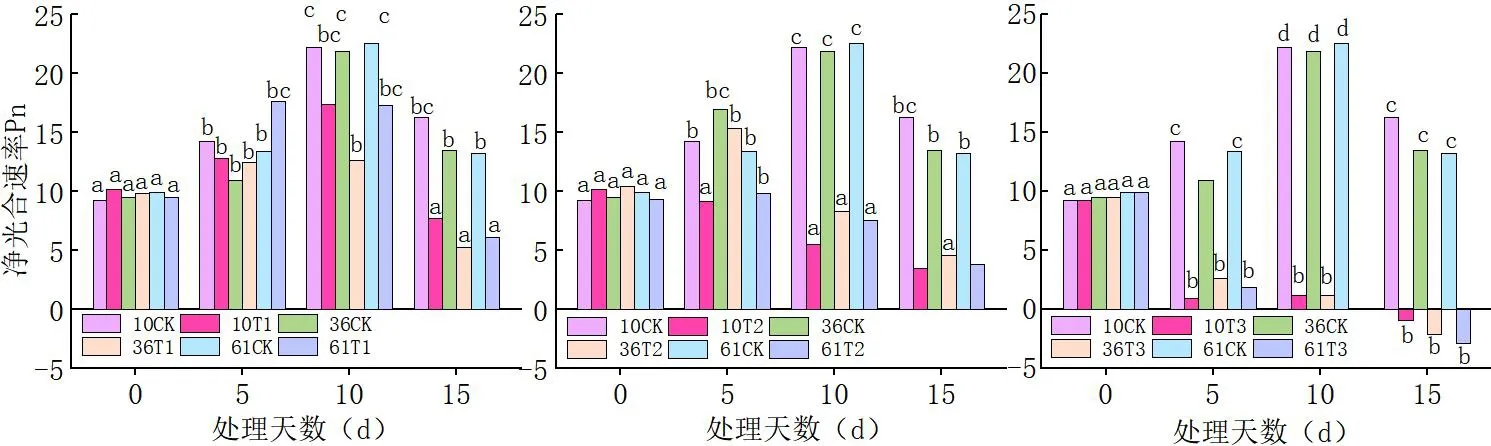
图2 低温胁迫下棉花净光合速率(Pn)变化
2.2 蕾期低温胁迫对气孔导度(Gs)的影响
研究表明,T1(20℃/15℃)与T2(15℃/10℃)、T3(10℃/5℃)存在显著差异,总体上温度下降气孔导度值变小;同一温度下不同时间各处理也差异明显,T1、T2处理下Gs值先升后降。T3处理下Gs值直线下降。胁迫温度从 20℃/15℃降至10℃/5℃时,蕾期棉花参试品种的Gs含量均大幅度下降,低于 15℃/10℃的低温处理叶片的Gs含量虽呈下降趋势,但降幅较小,10℃/5℃处理10 d 植株死亡。胁迫 15d 各处理的Gs含量均下降到最低水平,不同天数间差异显著。从单变量全因子分析的结果来看,温度、时间对蕾期棉花叶片气孔导度的变化均有影响,而且温度与处理天数有交互作用,P<0.001。温度和时间产生的低温危害效应与Pn相似。图3
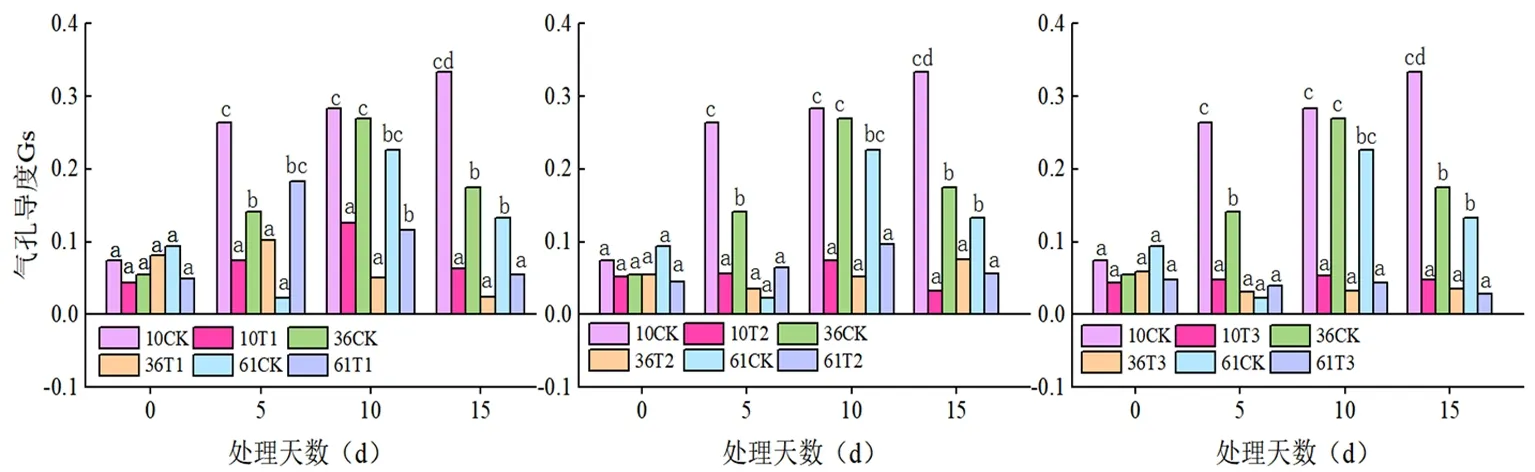
图3 低温胁迫下气孔导度(Gs)变化
2.3 蕾期低温胁迫对细胞间CO2浓度(Ci)影响
研究表明,不同低温胁迫初期(5 d)各品种的Ci值差异不显著,胁迫后期(15 d)各品种Ci值差异较为显著。T1胁迫10 d内各品种蕾期棉花的Ci差异不明显,T3胁迫下同一处理温度不同胁迫天数之间差异较明显。T2、T3胁迫下各品种的Ci与CK组的差异均随处理持续时间的延长而加大,T1(20℃/15℃)处理下Ci值呈升-降-升的变化趋势,T2(15℃/10℃)处理下Ci呈先升后降的变化趋势,T3(10℃/5℃)处理下Ci的变化趋势呈急剧上升,总体上各低温胁迫组的Ci值均大于对照。T3(10℃/5℃)胁迫 15 d 时达最大值。新陆早61号的Ci值上升幅度最小,新陆早61号蕾期的抗冷性较好。图4
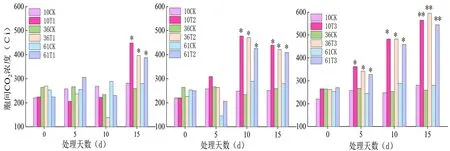
图4 低温胁迫下细胞间CO2浓度(Ci)变化
2.4 蕾期低温胁迫对蒸腾速率(Tr)的影响
研究表明,各低温处理组的Tr均呈降低趋势,各处理间差异显著。同一低温条件下,随低温处理时间的延长,3个蕾期棉花的Tr均显著下降。各处理中,新陆早61号的Tr值随处理时间延长降幅显著低于其余3个品种,而新陆早36号的Tr值下降速度最快;T2(15℃/10℃)低温下相对新陆早36号的Tr值最低,新陆早61号最高;T3(10℃/5℃)低温下相对其它种质新陆早10号的Tr值最低,新陆早36号最高。在T2(15℃/10℃)低温胁迫10 d 时,新陆早36号Tr值出现一个跃升,显著高于其他2个品种。T1、T2低温胁迫下新陆早61号的抗寒性较强,T3低温胁迫下新陆早36号抗寒性较强,新陆早10号抗寒性最弱。Tr于T3(10℃/5℃)低温胁迫10 d 和15 d 下均表现为缓慢下降。时间和温度均对蕾期棉花的冷害起较重要的效应作用,本研究中蕾期棉花的Tr值极显著(P<0.01)受“温度×时间”互作的影响,且时间和温度对不同蕾期棉花品种的危害效应同于上述Pn和Gs。图5

图5 低温胁迫下蒸腾速率(Tr)变化
2.5 蕾期低温胁迫对水分利用效率(WUE)影响
研究表明,水分利用效率WUE是植物生产中单位水分的能量转化率指标。T1低温胁迫处理时,3个品种的蕾期棉花叶片水分利用效率增大, T2处理时先升高后下降、T3处理时降低。低温胁迫强度大于T2及低温胁迫时间超过10 d时蕾期棉花可造成较大的影响。图6

图6 低温胁迫下水分利用效率(WUE)变化
T1处理的10 d、 T2处理的5 d、T3处理的10 d、15 d均与CK之间差异显著。T1、T2处理5 d时,新陆早61号、新陆早36号发生了较大的跃升,推测是新陆早61号、新陆早36号可通过提升水分利用效率(WUE)来增强植株的抗冷性,推测T1、T2处理5 d内为蕾期棉花植株可调节恢复的范围。T3处理下的蕾期棉花的水分利用效率(WUE)表现为急剧降低,当T3胁迫15 d时为负值,表明T3的低温胁迫处理对蕾期棉花可造成较大的影响,且胁迫超过10 d可造成植株死亡。当T1、T2处理持续0~5 d时,WUE不断增加,即叶片WUE在T1、T2处理持续5 d时达到最大,之后开始迅速下降。T1、T2处理持续5 d是蕾期棉花受低温胁迫的极限, T3胁迫强度或 T1、T2胁迫强度持续5 d以上,都会使叶片无法通过调节气孔的开张和蒸腾速率缓解低温对植株造成的伤害。图6
研究表明蕾期棉花在轻度(T1-20℃/15℃)低温胁迫下水分利用率(WUE)呈上升趋势的现象一致,表明蕾期棉花可在轻度(T1-20℃/15℃)低温胁迫下存活。中度(T2-15℃/10℃)下,处理前5 d水分利用率(WUE)升高,而处理持续时间超过5 d后水分利用率(WUE)下降,中度(T2-15℃/10℃)低温胁迫持续5 d是两蕾期棉花的生存阈值。
2.6 蕾期低温胁迫对气孔限制值(Ls)的影响
研究表明,T1处理时,处理5d时的气孔限制值Ls下降比较急剧,低温胁迫初期会迫使蕾期棉花叶片对光能的利用效率急剧下降。T1处理过程中表现为降升降的趋势。T1和T3处理Ls峰值出现在第10 d,T1处理5 d新陆早10号、新陆早36号、新陆早61号气孔限制值分别为0.205、0.068、0.099;T2处理5 d新陆早10号、新陆早36号、新陆早61号气孔限制值分别为0.270、0.570、0.236; T3处理5 d新陆早10号、新陆早36号、新陆早61号气孔限制值分别为0.256、0.063、0.228,各处理间差异不显著(P>0.05)。图7
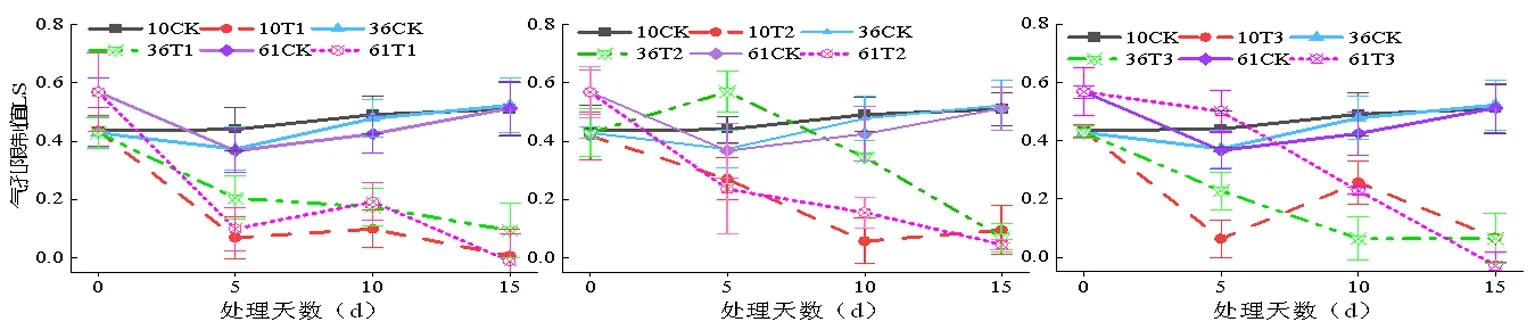
图7 低温胁迫下水分利用效率(WUE)变化
T1和T2处理下新陆早36号的气孔限制值Ls受影响最小,T3处理则新陆早61号的气孔限制值Ls表现为受影响最小。而T1、T2和T3处理的新陆早10号的气孔限制值Ls都表现为受的影响最大。新陆早10号品种的Ls平均值在T1、T2和T3胁迫下分别比CK组下降了53.41% 、38.64%和41.82%。蕾期棉花在遭受低温胁迫时会表现出Ls大幅度降低。同一低温胁迫强度处理下,随着低温处理时间的延长,蕾期棉花气孔限制值Ls越小,与CK处理组的差异越来越大。低温胁迫时非气孔限制是导致蕾期棉花光合速率降低的主要因素[20]。
3 讨 论
Pn是光合系统功能的直接体现,大多数的植物在遭遇低温胁迫时净光合速率Pn都表现出明显下降[21]。不仅环境因子和气孔导度会导致Pn下降,非气孔限制因素也是可能降低Pn的因素。植物的光合与蒸腾受气孔的制约,气孔对植物与大气进行水汽和 CO2交换起到很重要的作用[23]。
气孔限制因素引起的Pn下降表现为Gs、Tr下降,Ci也相应下降,非气孔限制因素引起的Pn下降表现为Gs、Tr下降的同时Ci升高[24-25];这与研究中气孔导度Gs、净光合效率Pn下降时Ci升高的结果一致,表明本研究中限制蕾期棉花Pn的主要原因是非气孔因素,与郭菊兰等[26]表明非气孔因子限制是迫使秋茄幼苗净光合速率Pn下降的主要因素。蕾期棉花相同胁迫时间下,T1、T2的温度处理Pn下降主要受气孔限制因素引起光合作用受限制所致,而T3以下低温处理Pn下降很大程度来源于非气孔限制因素。
低温胁迫下,净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)均逐步下降,低温胁迫越重,光合参数下降幅度越大;重度低温已严重损害叶片光合机构,极大抑制了光合作用[22]。与研究中蕾期棉花在低温胁迫后, 光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)均比对照明显下降。且下降趋势随胁迫程度加大而增大,表现为T3>T2>T1的结果相一致。研究发现蕾期棉花在中度低温胁迫(T2-15℃/10℃)下处理超过5 d及重度(T3-10℃/5℃)低温胁迫下水分利用率(WUE)呈下降趋势。与郑芊彤等[18]对三七进行低温处理7 d后发现重度处理的WUE先上升后下降,轻度处理的WUE随持续时间的延长而上升的结果一致。水分利用率(WUE)的变化趋势不仅取决于胁迫的温度,还与胁迫持续时长有着密切的关联。
试验中蕾期棉花随低温胁迫时间的延长,Ls呈下降趋势,低温环境使得蕾期棉花叶片对光能的利用效率下降,与许耀照等[16]表明T2(15℃/8℃)与T1(19℃/12℃)低温处理下气孔限制值的降低程度达到显著差异的研究结果一致。
4 结 论
4.1可根据蕾期棉花的气孔导度(Gs)、蒸腾速率(Tr)、细胞间二氧化碳浓度(Ci)、水分利用率(WUE)来判断其受到的低温胁迫程度。轻度胁迫下蕾期棉花光合速率降低是由气孔限制因素导致;中度胁迫下蕾期棉花光合速率降低的主要原因先为气孔因素后转为非气孔因素;重度胁迫下非气孔因素是限制蕾期棉花光合作用的主要因素。
4.2低温胁迫对蕾期叶片的光合作用有抑制的作用。轻度胁迫时新陆早61号的耐低温性较好,中度、重度胁迫时新陆早36号相比于新陆早61号和新陆早10号耐低温性更好。胁迫温度越低,对蕾期棉花生长的抑制作用越明显,且相同低温度值处理下,低温处理的时间持续越长,对蕾期棉花生长的抑制作用越明显。
