异源表达棉花GhPAO3基因对拟南芥种子萌发的影响
2022-08-03成新琪庞芳芹汤欣欣胡孝明朱华国
成新琪, 庞芳芹, 汤欣欣, 乌 兰, 胡孝明, 朱华国
(黄冈师范学院生物与农业资源学院/经济林种质改良与资源综合利用湖北省重点实验室, 湖北 黄冈 438000)
多胺(Polyamines,PAs)是含有两个或两个以上氨基,并存在于生物体内具有生物活性的脂肪族含氮碱。植物体多胺主要有腐胺(Putrescine,Put)、亚精胺(Spermindine,Spd)、精胺(Spermine,Spm)和热精胺(Thermospermine,Tspm)。此外,生物体内存在着稀有多胺[1]。研究发现,多胺在植物生长和适应环境变化中扮演重要的角色[2-3],诸如细胞分裂和分化、胚胎发生、组织发生、生物以及非生物胁迫的响应等[4-8]。目前,多胺主要的合成和分解代谢途径已基本被揭示[9],与植物体多胺合成和代谢有关的部分基因已被克隆,并获得转基因阳性植株[10-11]。
多胺的降解主要是通过二胺氧化酶(DAO)和多胺氧化酶(PAO)催化来实现,二胺氧化酶氧化腐胺和尸胺,产物包括4-氨基正丁醛、氨和H2O2。多胺氧化酶有多个同源基因[10-11],其代谢途径分为两类,一类是催化Spm和Spd生成1,3-丙二胺(Dap)、H2O2和3-氨丙基-4-氨基正丁醛或4-氨基正丁醛;另一类是催化Spm生成Spd,再生成Put,同时生成3-氨基正丁醛和H2O2[4-6]。多胺氧化产生H2O2可以作为一种信号分子参与植物体内细胞的程序性死亡、细胞的防御性反应、细胞质木质素合成和生物与非生物胁迫等过程[12]。
H2O2是生物体细胞在代谢中产生的一种活性氧(ROS),正常生长情况下,植物体内的ROS清除系统能使H2O2的含量保持一定的水平,使植物体免受伤害,但在逆境胁迫条件下,活性氧的产生超出系统清除能力范围时,对植物体造成氧化损伤[13]。对H2O2深入的研究,认识到H2O2对植物生长和代谢的影响,H2O2参与了植物种子萌发和根的生长等过程,尤其是当植物受到胁迫时,H2O2的作用已成为了胁迫研究的热门课题[14]。ROS对植物生长发育也有重要作用,能对植物细胞产生一定程度的损害,但适宜浓度的ROS对激活植物应对胁迫的防卫有重要作用[15-18]。本研究以野生型(WT)和过表达GhPAO3转基因拟南芥为材料,研究不同浓度NaCl对拟南芥种子萌发的影响,揭示外源多胺对转基因拟南芥种子子叶变绿的影响,为研究多胺响应盐胁迫的生理机制提供理论基础。
1 材料与方法
1.1 实验材料处理
选取WT和3个纯合转GhPAO3基因株系(3-5、3-15和3-19)的拟南芥种子为实验材料,用15%次氯酸钠消毒20 min,不断振荡,在超净台中用无菌水冲洗3~4次,播种于1/2 MS固体培养基上,低温(4 ℃)春化2 d,转移至22 ℃人工培养箱中培养。
1.2 转基因拟南芥植株盐处理
以转基因株系3-15为实验材料,种植于0、50、100、150、200 mmol/L NaCl的1/2 MS培养基上,统计其发芽率和子叶变绿的百分率。
1.3 外源多胺处理
不同浓度(0.2、1、2 mmol/L)的Put、Spd、Spm、D-Arg 和1,8-DO处理,调查统计WT和转基因拟南芥子叶变绿的百分率。
1.4 H2O2含量测定
取WT和转基因株系相同部位叶片(0.2~1.0 g),放入含有液氮研钵中充分研磨,将粉末移至2 mL离心管中,按重量(g)∶体积(mL)=1∶9的比例往离心管中加入0.86%的生理盐水,充分混匀,4 ℃,200 r/min离心10 min,取上清液于干净离心管进行H2O2含量测定,采用南京建成科技有限公司H2O2测定试剂盒。
1.5 数据统计
发芽率(%)=(第n天发芽的种子数/所用种子总数)×100%;
子叶变绿百分率(%)=(第n天子叶变绿的数量/所用种子总数)×100%,
式中,n为种子发芽的天数,子叶变绿数量指拟南芥发芽时叶片变为绿色的数量,发芽在22 ℃恒温培养箱中进行,含水量恒定,每天记录发芽种子数和子叶变绿的数量,实验设3次重复。
2 结果与分析
2.1 NaCl处理下转GhPAO3拟南芥种子萌发情况
不同浓度NaCl条件下WT和转GhPAO3拟南
芥种子萌发的结果表明,在0(图1 A)和50 mmol/L NaCl(图1 B)处理中,WT和转基因种子发芽率无明显差异;100 mmol/L NaCl处理转基因拟南芥发芽率略低于WT(图1 C);150 mmol/L NaCl处理后推迟1 d发芽(图1 D),且转基因株系发芽率低于WT;200 mmol/L NaCl处理下则推迟至第4天发芽(图1 E),WT和转基因的发芽率都显著下降,第7天的发芽率分别为54%和49%。随着NaCl浓度的提高,拟南芥种子的萌发抑制更加明显,转基因拟南芥种子对NaCl更加敏感。
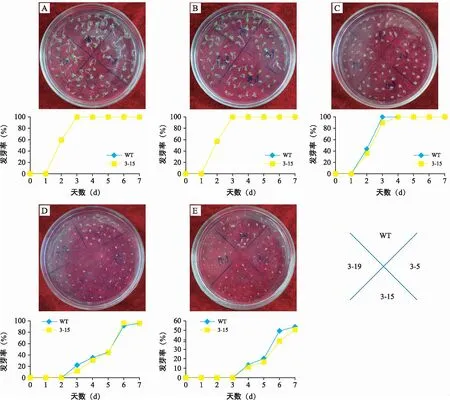
注:A~E分别表示0、50、100、150、200 mmoL NaCl处理下WT和转基因拟南芥种子发芽率;图片上方是对应处理条件下种子的发芽率。图1 NaCl胁迫下WT和GhPAO3拟南芥种子的发芽率Fig.1 Germination rate of WT and GhPAO3 seeds under NaCl stress in Arabidopsis
2.2 NaCl处理下外源多胺和多胺抑制剂对拟南芥子叶的影响
种子萌发实验发现,100 mmol/L NaCl处理下WT和转基因拟南芥种子发芽率开始出现差异,且子叶变绿情况差异明显。WT在第3天出现绿色子叶,而转基因株系在第4天出现绿色子叶,且转基因拟南芥子叶变绿的数量明显低于WT(图2 A)。为揭示转基因株系在100 mmol/L NaCl处理下出现子叶变绿差异的原因,在100 mmol/L NaCl条件下分别添加不同浓度(0.2、1、2 mmol/L)外源多胺(Put、Spd、Spm)、多胺抑制剂(D-Arg)和多胺氧化酶(PAO) 抑制剂(1,8-DO)研究对子叶变绿的影响。结果表明,Put(2 mmol/L)、Spd(2 mmol/L)、Spm(2 mmol/L)处理时,WT与转基因株系子叶变绿存在差异。加入Put时,WT和转基因株系子叶变绿数量均有所提高,且转基因株系子叶变绿提前(图2 B),而加入Spd和Spm后,WT和转基因株系子叶变绿都推迟1 d(图2 C和图2 D),加入不同多胺处理时转基因子叶变绿百分率均低于WT(图2 A-图2 D),但在第7天对照(图2 A和图3 A)和Put处理(图2 B和图3 B)子叶变绿百分率均达到100%;加入Spd和Spm,第7天仍存在差异(图2 C和图3 C;图2 D和图3 D)。而当加入D-Arg(1 mmol/L)和1,8-DO(1 mmol/L)后,第7天也未出现子叶变绿的现象(图2 E和图3 E;图2 F和图3 F)。因此,推测NaCl处理和过表达GhPAO3基因可能通过改变H2O2含量影响拟南芥子叶变绿。

注:A为100 mmol/L NaCl处理下WT和转GhPAO3拟南芥子叶变绿的百分率;B、C、D、E、F分别为100 mmol/L NaCl和多胺(Put、Spd、Spm、D-Arg和1,8-DO)处理下转GhPAO3和拟南芥子叶变绿的百分率。图2 NaCl处理和外施多胺条件下WT和GhPAO3拟南芥种子的子叶变绿的百分率Fig.2 Percentage of greenish cotyledons of WT and GhPAO3 seeds under NaCl treatment and external application of polyamines in Arabidopsis
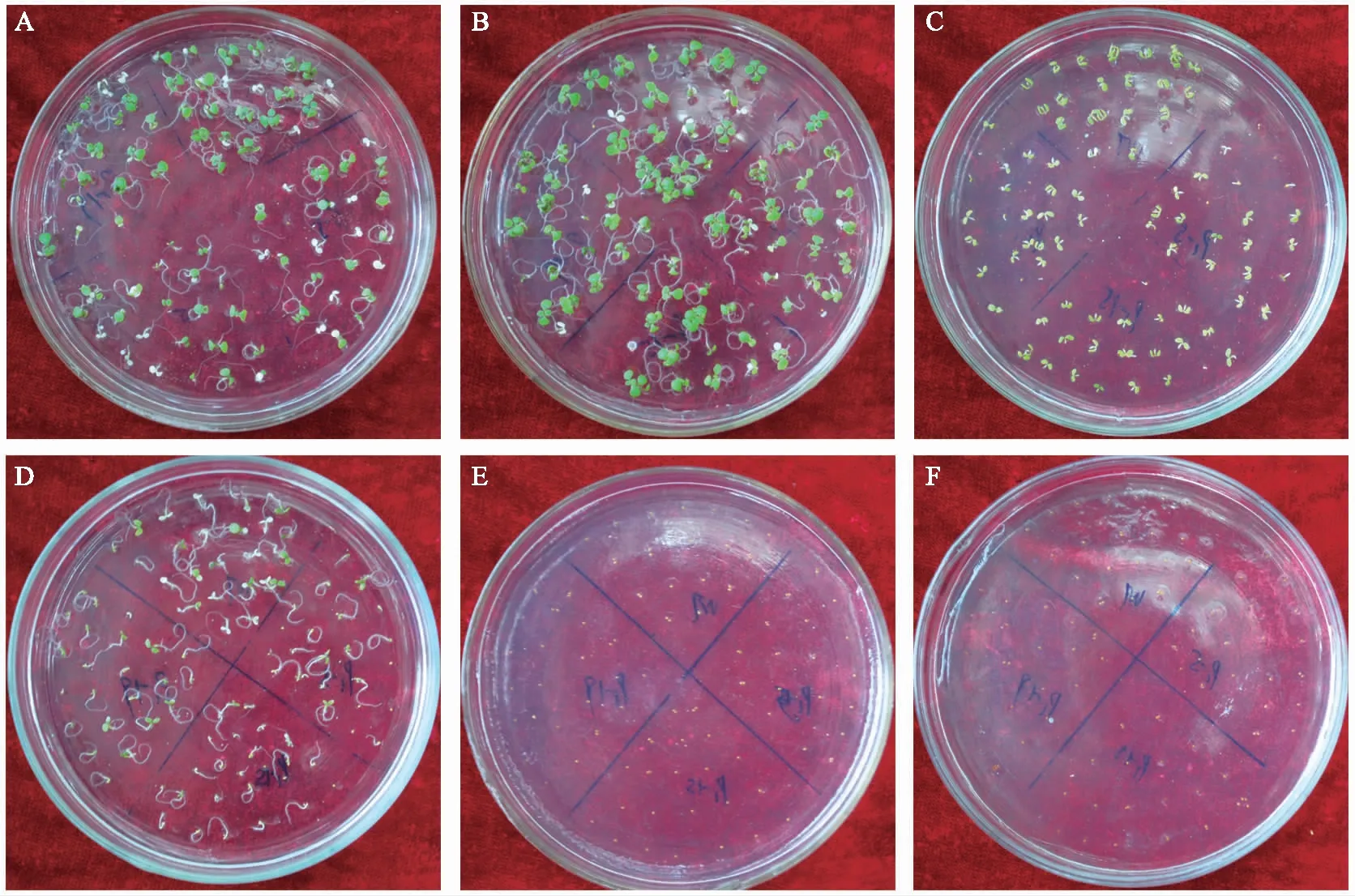
注:A为100 mmoL NaCl处理下GhPAO3和拟南芥第7天子叶变绿的差异;B、C、D、E、F分别为多胺(Put、Spd、Spm、D-Arg和1,8-DO)处理下WT和转基因GhPAO3拟南芥第7天子叶变绿的差异。图3 NaCl处理和外施多胺条件下WT和转基因GhPAO3拟南芥第7天子叶变绿的百分率 Fig.3 Percentage of cotyledons turning green on the 7th day of WT and GhPAO3 under NaCl treatment and external application of polyamines in Arabidopsis
2.3 外施多胺、D-Arg、1,8-DO对拟南芥子叶的影响
为进一步研究子叶变绿差异,在1/2 MS培养基上分别加入外源Put、Spd、Spm、D-Arg和1,8-DO统计子叶变绿情况(图4和图5)。结果显示,1/2 MS培养基上WT和转基因株系子叶变绿没有差异,第3天出现子叶变绿,第6天达到100%(图4 A和图5 A)。添加外源1,8-DO处理后WT和转基因株系的子叶变绿的百分率均有所提高,其中加入Put、Spm、1,8-DO后,子叶变绿提前出现,即第2天出现绿色子叶(图4 B、D、F);加入Spd,第3天出现绿色子叶(图4 C);而加入多胺抑制剂D-Arg时,绿色子叶推迟至第5天出现(图4 E)。此外,加入外源多胺(Put、Spd、Spm)后,WT子叶变绿的百分率高于转基因株系;而加入D-Arg后,转基因子叶变绿百分率高于WT;加入1,8-DO后,WT和转基因子叶变绿百分率基本一致。因此,加入外源多胺时,对拟南芥子叶变绿具有促进作用,但与WT相比,转基因株系的子叶变绿的百分率有所降低,可能是过表达株系产生更多的H2O2,对子叶变绿具有一定抑制作用。
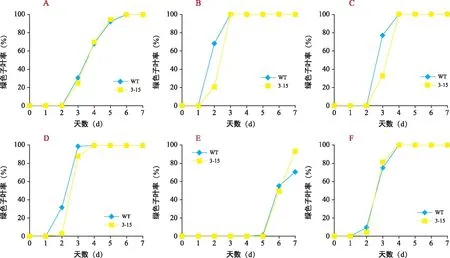
注:A为1/2 MS培养基WT和转GhPAO3拟南芥子叶变绿百分率差异;B、C、D、E、F分别加入多胺(Put、Spd、Spm、D-Arg和1,8-DO)处理下转GhPAO3和拟南芥子叶变绿百分率差异。图4 外施多胺条件下WT和转GhPAO3拟南芥种子子叶变绿百分率Fig.4 Rate of green cotyledons of WT and GhPAO3 seeds under external application of polyamines in Arabidopsis

注:A为1/2 MS培养基WT和转GhPAO3拟南芥第7天子叶变绿差异;B、C、D、E、F分别为多胺(Put、Spd、Spm、D-Arg和1,8-DO)处理下WT和转基因GhPAO3拟南芥第7天子叶变绿差异。图5 外施多胺条件下WT和转GhPAO3拟南芥第7天子叶变绿情况 Fig.5 Green cotyledon of WT and GhPAO3 transgenic on the 7th day under the condition of external application of polyamines in Arabidopsis
2.4 外施多胺和多胺抑制剂对拟南芥H2O2含量的影响
外施多胺和多胺抑制剂条件下WT和转基因株系H2O2含量分析显示,当加入Put、Spm或1,8-DO后,与WT相比,转基因株系产生H2O2的量明显升高,差异达极显著水平;加入Spd后,转基因株系产生H2O2的量升高,差异达显著水平;当加入D-Arg后,转基因株系产生H2O2的量明显降低,差异达极显著水平(图6)。
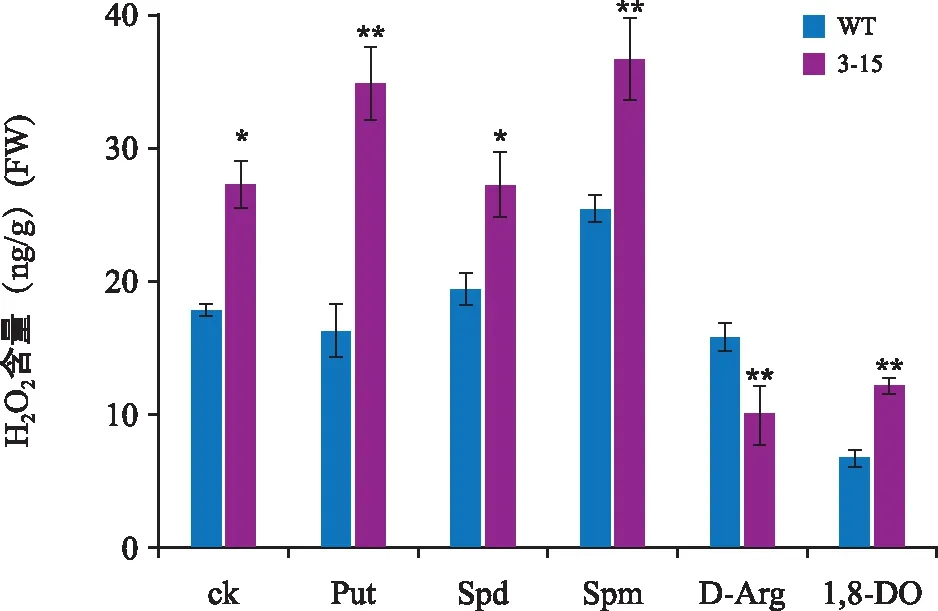
注:“*”表示差异显著(p<0.05);“**”表示差异极显著(p<0.01)。图6 不同处理条件下WT和转GhPAO3拟南芥H2O2含量Fig.6 Detection of H2O2 content in WT and GhPAO3 transgenic Arabidopsis under different treatments
3 结论与讨论
种子萌发是植物生长发育过程中的重要阶段,受温度、水分和氧气等影响,同时也受植物体内激素和多胺等的调节,研究发现,添加外源多胺可以促进莴苣种子[19]或抑制水稻种子[20]的萌发。越来越多的研究表明,多胺参与植物抗盐方面的重要作用[2],PAO催化多胺产生H2O2,参与植物的生长发育和非生物胁迫[10]。
NaCl处理(图2 B)和正常(图4 B)条件下,加入外源Put均促进子叶变绿的产生。Afzal等[21]和Verma、Mishra[22]研究发现,外源Put能促进盐处理下种子的萌发,提高种子和叶片的抗氧化能力,减少脂膜过氧化的程度[7],有利于种子萌发和植株生长发育,与本研究结果相似。同时,NaCl处理条件下,外源Spd(图2 C)和Spm(图2 D)处理下拟南芥子叶变绿的百分率降低,并且所有处理条件下,转基因株系子叶变绿的百分率低于WT,同时发现转基因株系较WT积累更多H2O2[11],推测NaCl处理和外源多胺处理下,Spd和Spm大量进入拟南芥体内,之后被 PAO分解,产生更多的H2O2对细胞膜产生伤害作用,并且过表达株系促进植物体内多胺降解产生H2O2,导致转基因株系子叶变绿百分率低于WT,本研究结果与Wang等[23]研究的结果一致。正常条件下,外源Put(图4 B)、Spd(图4 C)、Spm(图4 D)和1,8-DO(图4 F)处理拟南芥种子,其子叶变绿百分率均有所提高,但转基因株系子叶变绿百分率低于WT(图4),H2O2测定显示,转基因株系中H2O2含量高于WT(图6),与图4结果一致,是因为转基因株系过表达氧化多胺产生更多H2O2,但是没有NaCl处理条件下H2O2含量积累明显[11]。因此,与对照相比,外源多胺对拟南芥有一定的促进作用,与张凤芝等[24]研究莴苣种子萌发结果一致。
植物体H2O2的产生和清除保持着动态平衡,细胞内活性氧的含量较低时,活性氧有一定的生理功能,使植物的伤害降到最低。胁迫条件下,H2O2等活性氧产生和清除的动态平衡被打破,生长量高于清除量,使植物体内H2O2等ROS含量积累。因此,H2O2在植物体内扮演双重角色,低浓度的H2O2可以作为信号分子参与逆境胁迫,促进植物生长和提高抗逆性,而高浓度的H2O2对细胞膜的破坏和氧化胁迫的加剧,甚至加速植物细胞程序性死亡的进程[2,10]。研究发现,H2O2等ROS的含量过量会导致细胞内的生物大分子氧化损害,高含量的H2O2使蛋白分子内或分子间的二硫键产生交联,使含有巯基的酶活性降低甚至失活,同时,H2O2还可以激活蛋白水解酶等的活性,加快植物蛋白的降解,最终使植物体内的生理反应受到抑制[25]。因此, NaCl处理过表达GhPAO3拟南芥时,拟南芥体内通过NaCl胁迫和PAO氧化多胺同时产生H2O2,产生的H2O2含量超过植物体承受的阈值,对植物产生危害,从而表现对NaCl胁迫更敏感。关于拟南芥过表达GhPAO3在NaCl胁迫产生H2O2的阈值是后期研究工作的重点。
