我国夏季不同类型植被BVOCs 排放观测与模拟研究
2022-06-22王楚迪节龙飞李苗苗王添文李玲玉
王楚迪,节龙飞,李苗苗,王添文,高 阔,李玲玉
青岛大学环境科学与工程学院,山东 青岛 266071
近年来,我国部分地区的臭氧(O3)浓度呈逐年上升趋势,O3污染日益突出[1-2]. 挥发性有机物(VOCs)是近地面O3生成的关键前体物[3-4],VOCs 和氮氧化物(NOx)在紫外光作用下可发生光化学反应产生O3.因此,准确描述VOCs 排放特征是实现O3污染精准防控的基础和关键. VOCs 的来源分为人为源和天然源,其中全球天然源VOCs (BVOCs)排放量占VOCs排放总量的90%以上[5]. BVOCs 主要来自植被排放,且不同类型植被的排放水平和组成不同[6-7]. BVOCs包括异戊二烯、单萜烯、倍半萜烯和其他VOCs,BVOCs 所含的不饱和键具有高反应活性,极易在大气中发生光氧化反应[8-9]. 因此,研究不同类型植被BVOCs 排放特征对明确大气复合污染的来源和生成机制,以及研究精准有效的大气复合污染防治措施具有重要科学意义.
为研究我国本土不同类型植被BVOCs 排放特征,学者开展了排放观测和模拟研究. 自20 世纪90年代,我国相继开展了本土植物BVOCs 排放的测量工作,研究初期多采用静态封闭式采样法确定了我国不同植物排放的主要BVOCs 物种和排放规律[10-14].但是,静态封闭式采样法由于封闭系统无气体循环,可在短时间内导致封闭室内温度升高、CO2浓度降低、H2O 含量增加,造成植物异常排放,观测结果误差较高,难以准确描述不同植物类型的BVOCs 排放差异[15]. 目前,已有学者应用动态封闭式采样法对云南省[16-17]、内蒙古自治区[18]、浙江省[19]、陕西省[20]、天津市[21-22]、辽宁省[23]、北京市[24]部分树种的排放开展了定性和定量观测,积累了较为准确的BVOCs 排放速率,并研究了各植物的排放规律. 动态封闭式采样法因设有恒定流量的零空气循环系统,封闭室内的环境更接近自然状态,观测结果具有更高的准确性[15,22]. 但目前我国不同类型植物BVOCs 排放观测结果误差较大,且多集中于异戊二烯及部分主要单萜烯化合物;另外,部分研究仅开展定性测定,但单萜烯化合物种类多且排放占比较高,因此应开展更多的定量观测研究. 从20 世纪90 年代开始,学者基于排放模拟研究我国BVOCs 排放特征,采用不同的排放算法和模式,建立了全国和区域BVOCs 排放清单[25-30],研究了我国不同类型植被BVOCs 排放特征. 然而,由于清单中使用的植被和气象数据分辨率较低,排放估算结果具有较高的不确定性[31],难以准确探究我国本土不同植被类型的排放差异[25,32-33].
该研究选取不同类型的植物,应用动态封闭式采样法和热脱附-气相色谱质谱联用(TD-GC-MS)技术对各植物BVOCs 排放进行现场观测和定量分析,获得各植物单萜烯排放速率及排放组成;基于该研究和文献建立我国本土化的排放因子数据库;应用MEGAN 2.1 排放模式和中尺度气象模式WRF 构建2018 年夏季(6—8 月)我国高时空分辨率的BVOCs排放清单,研究我国6 种植被类型(阔叶树、针叶树、针叶阔叶混交林、灌木、草地、农作物)的BVOCs 排放特征及差异,以期为研究精准有效的大气复合污染防治措施提供基础数据.
1 材料与方法
1.1 BVOCs 排放观测
1.1.1 样品的采集和分析
分别选取阔叶树、针叶树、灌木、草地、农作物5 种植被类型的植物进行BVOCs 排放的野外和室内盆栽观测,阔叶树种包括广玉兰,针叶树种包括油松、雪松、马尾松、侧柏,农作物物种为小麦,草地植物为紫马唐,灌木为大叶黄杨. BVOCs 排放样品的采集时间为2020 年和2021 年的夏季(6—8 月),野外观测选择晴朗无风天气进行,观测点位于青岛大学校园内. 采用动态封闭式采样系统(见图1)进行BVOCs的采集,用聚四氟乙烯采样袋(110、18 L)或有机玻璃采样罩(73 L,草地植物观测)将长势较好的树枝或植株封闭,以恒定流速向采样袋内持续通入经活性炭吸附VOCs 后的空气,同时采样袋内的气体以相同流速流出,封闭室体积为110、73、18 L 时对应的零空气流速分别为20、20、2.5 L/min. 经过1~2 h 的平衡,使袋内空气混合均匀,利用Tenax 吸附管(Markes,英国)采集采样袋内的气体,采样流速为200 mL/min,采样时间为30 min,采样体积为6 L. 每种植物选择3个植株,每个植株采集3~4 个平行样品和1 个空白样品. 样品放置于恒温(4 ℃)冰箱密封保存,并在1 周内完成定量分析. 在平衡和采样过程中,测量封闭室外光合有效辐射、封闭室内温度,野外观测的温度为21~35 ℃,光合有效辐射为600~1 400 μmol/(m2·s);实验室盆栽观测的温度为25~30 ℃,光合有效辐射为900~1 000 μmol/(m2·s). 采样结束后,测定封闭室内植株叶片的干质量. 试验使用仪器设备包括空气泵(Rocker 430 型,台湾洛科仪器股份有限公司,中国)、空气采样泵(GilAir Plus STP 型,Sensidyne,美国)、质量流量计(CS200A 型,北京七星华创电子股份有限公司)、数据采集器(CR300 型,Campbell,美国)、温度传感器(109 型,Campbell,美国)和光量子传感器(LI-190R 型,LI-COR,美国).

图 1 动态封闭式采样系统示意Fig.1 Schematic of the dynamic enclosure system
样品经热脱附-气相色谱-质谱联用法(TD-GC-MS)进行14 种单萜烯组分的定量测定. 全自动热脱附装置(ATD II-26 型,Acrichi 公司,中国)的工作条件:系统压力为80 kPa;进样阀温度为150 ℃;热解吸温度为290 ℃;冷阱温度为—30 ℃. GC-MS(7890A-5975C型,Agilent,美国)的工作条件:载气流速为1.2 mL/min,分流比为1∶5;程序起始温度为35 ℃,保持2 min,然后以10 ℃/min 升至260 ℃,再升至280 ℃保持2 min;离子源为EI 源,离子能70 eV,温度为200 ℃. 色谱柱型号为Agilent DB-5 (30 m×0.25 mm ID×0.25 μm). 应用响应因子法和外标法(最小二乘法)对各单萜烯组分进行定量,标准样品体积分别为2、5、10、20、50、70 mL (标准样品体积分数为1×10—6),响应因子的相对标准偏差<20%,标准曲线R>0.99. 当采样体积为6 L 时,方法检出限为0.03~0.97 μg/m3,测量精密度为4%~15%.
1.1.2 排放速率计算
排放速率的计算公式[22]:

式中:ER 为排放速率,μg/(g·h);F为采样袋气体的流出流速,L/min,与零空气流入采样袋的流速相同;C为单萜烯质量浓度,μg/m3;m为叶片干质量,g.
利用Guenther 等[34-35]的标准化算法,将实际条件下的单萜烯排放速率转换为标准条件〔温度为30 ℃,光量子通量密度为1 000 μmol/(m2·s)〕下的排放速率.该研究中排放速率均为标准排放速率,标准化后的误差为25%[34].
1.2 BVOCs 排放清单的建立
1.2.1 MEGAN 排放模式
利用MEGAN 2.1 排放模式估算了2018 年夏季我国6 种不同类型植被的BVOCs 排放量. MEGAN作为一种先进的全球和区域尺度的BVOCs 排放估算模式[31],已经被广泛应用于全球[36-37]和区域[37-40]BVOCs 排放量的估算. BVOCs 的排放量计算公式[35]:

式中:E为BVOCs 排放量,mg/(m2·h);ε为标准条件下的冠层尺度排放因子,mg/(m2·h);γ为环境校正因子,由于标准排放因子用以描述在标准状态下的BVOCs 排放水平,不能反映真实环境中的排放,因此通过γ对其进行校正,包括冠层环境内温度和辐射、叶龄、土壤湿度、CO2环境浓度等对冠层排放的影响,该研究中未考虑土壤湿度和CO2的影响;ρ为逸散系数,描述冠层内BVOCs 由于生物、化学和物理过程导致的生成和损耗,该研究中ρ取值为1[41-42].
1.2.2 气象数据
应用中尺度气象模式WRF 模拟MEGAN 2.1 所需气象参数,包括2 m 温度、太阳短波辐射、风速、相对湿度、降雨、气压等,时间分辨率为1 h,水平分辨率为36 km×36 km. 模拟区域覆盖整个中国,中心经纬度为34.53°N、108.92°E,包括128×188 个网格.气象初始场和侧边界场为NCEP 提供的全球1°×1°分辨率的再分析数据—NCEP/DOE 再分析Ⅱ(NCEP/DOE-R2,http://rda.ucar.edu/datasets/ds091.0).
1.2.3 植被分布
利用《中华人民共和国植被图(1∶1 000 000)》[43]获得各植被类型/树种的空间分布. 该植被图是目前我国最全面、详细的植被图集,包含阔叶林、针叶林优势树种/植被类型、栽培植被、高山植被、草原、草丛、草甸、荒漠、沼泽、灌丛等植被型组. 结合中国森林资源清查结果、中国统计年鉴、中国天然草地类型系统,对《中华人民共和国植被图(1∶1 000 000)》中的植被重新进行了分类,共得到82 种植被物种/类型的分布,包括10 种阔叶优势树种、18 种针叶优势树种、6 种森林类型、13 种主要农作物、7 个灌丛亚类、28个草地亚类. 以往研究中植被分类较粗,多分为4~12 类,MEGAN 2.1 模型中植被类型默认有15 种.
1.2.4 排放因子
基于叶片排放速率和叶生物量,利用全冠层环境模型外推计算得到各植被的标准冠层排放因子[35]. 该研究中使用的排放速率和叶生物量为笔者所在课题组已有研究结果[37,40]. 利用统计学方法,构建了我国BVOCs 排放速率数据库;基于我国各省份植被蓄积量和产量的调查统计数据(不包括港澳台地区数据),利用生物量分配模型,估算82 种植被的叶生物量. 其中,各省份各优势树种各龄级的蓄积量来源于《第九次全国森林资源清查统计(2014—2018 年)》,各省份主要农作物的经济产量来源于《中国统计年鉴》,各省份各草地亚类的产草量来源于《中国草地资源数据》.
2 结果与讨论
2.1 不同植物单萜烯排放特征
2.1.1 排放速率
8 种植物单萜烯标准排放速率如表1 所示,其中,马尾松排放速率最高,大叶黄杨排放速率最低. 根据Guenther 等[44-45]的排放速率划分标准,将单萜烯排放速率为>3.0、>0.6~3.0、≤0.6 μg/(g·h)的植物划分为高、中、低排放强度植物. 广玉兰、马尾松和雪松属于单萜烯高排放强度植物,油松属于中排放强度植物,侧柏、小麦、紫马唐和大叶黄杨属于低排放强度植物. 从植物类型来看,针叶树单萜烯排放强度最高,其次是阔叶树,草地和农作物排放强度较低,灌木排放强度最低.

表 1 8 种植物单萜烯标准排放速率Table 1 Standard emission rates of monoterpenes from 8 plants μg/(g·h)
2.1.2 排放组成
所有植物排放的单萜烯中,均检出月桂烯、α-松油烯、柠檬烯、顺式-β-罗勒烯、γ-松油烯、异松油烯,其余化合物在个别植物的排放中被检出. 油松和雪松不排放3-蒈烯,侧柏和小麦不排放三环烯、莰烯和反式-β-罗勒烯,紫马唐不排放α-蒎烯、β-蒎烯和水芹烯,大叶黄杨不排放三环烯和香桧烯. 广玉兰和马尾松排放的单萜烯化合物种类(14 种)最多,侧柏、小麦、紫马唐排放的单萜烯化合物种类(11 种)最少,油松、雪松排放的单萜烯化合物种类均为13 种,大叶黄杨排放的单萜烯化合物种类有12 种.
各植物单萜烯化合物排放速率占比如图2 所示.由图2 可见:广玉兰以排放α-蒎烯、柠檬烯、顺式-β-罗勒烯为主,分别占总单萜烯排放速率的30.35%、21.45%、18.73%;马尾松以排放异松油烯和γ-松油烯为主,分别占总单萜烯排放速率的31.14%和24.13%;油松以排放顺式-β-罗勒烯为主,占总单萜烯排放速率的36.45%,其次是柠檬烯,占比为28.08%;雪松以排放柠檬烯和α-蒎烯为主,分别占其总单萜烯排放速率的51.60%和17.91%;侧柏以排放香桧烯为主,占比为28.44%;小麦以排放异松油烯为主,占比为39.93%;紫马唐以排放反式-β-罗勒烯、异松油烯为主,分别占总单萜烯排放速率的28.50%、27.26%;大叶黄杨以排放柠檬烯、异松油烯为主,分别占总单萜烯排放速率的27.08%、18.75%.

图 2 各植物单萜烯化合物排放速率占比Fig.2 Contribution of monoterpene emission rates by compounds from each plant
2.1.3 不确定性分析
与已有研究结果相比(见表2),笔者研究中马尾松和雪松的单萜烯排放速率高于其他研究结果[13,20,22],但侧柏、油松和广玉兰的单萜烯排放速率较其他研究结果[13,20,46-47]低. 差异的原因可能在于采样方法的不同,此外不同温度和辐射条件下的排放速率标准化后将产生不同的误差. 从排放组成来看,笔者研究中油松主要排放的化合物为柠檬烯、顺式-β-罗勒烯、β-蒎烯,与吕迪[20]的研究结果较为一致. 吕迪[20]研究表明,油松、雪松、侧柏排放速率最高的化合物均为α-蒎烯,而笔者研究中分别为顺式-β-罗勒烯、柠檬烯和香桧烯. 从排放强度来看,笔者研究与其他研究结果均表明,雪松和油松分别为高排放强度和中排放强度植物;笔者研究中侧柏为低排放强度植物,而其他研究中其为中排放强度植物.
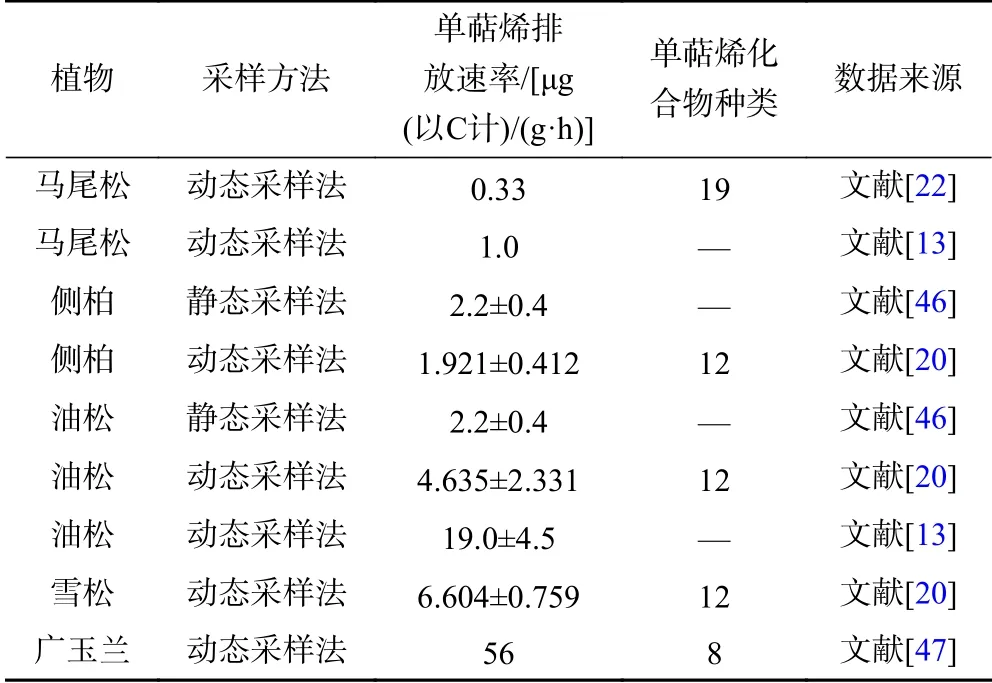
表 2 该研究与其他本土植被BVOCs 观测研究结果对比Table 2 Comparison with BVOCs observations of other native vegetation
BVOCs 排放受生物环境和非生物环境的影响,其中环境因素包括温度、辐射、相对湿度、大气CO2和O3浓度、土壤水分、土壤氮含量以及土壤微生物等[47-49]. 因此,由于生长环境与气候条件不同,在不同地区,同一植物的BVOCs 排放机制和特征可能存在较大差异. 该研究选择北方城市青岛市开展了初步的观测研究,为研究我国不同植被类型的排放特征与差异提供了初步的基础数据积累. 但难以准确评估我国其他地区不同植被类型的排放差异,为全面了解我国不同地区不同植被类型的排放特征,未来应在不同地区(如我国南部、西部等典型地区)开展更多植物的观测研究. 此外,该研究重点关注不同植被类型的排放规律和差异,因此对每种植被类型选取了较为常见的森林或城市绿化植物开展单萜烯排放测量. 我国不同植被类型所包含的植物较多,该研究针对7 种植物开展的测量研究难以全面反映所有植物的排放特征.即便如此,该研究可为了解我国不同类型植物的排放差异提供初步的数据支撑.
2.2 我国夏季BVOCs 排放特征
2.2.1 BVOCs 排放水平及组成
2018 年我国夏季BVOCs 总排放量为33.6×1012g,异戊二烯、单萜烯、倍半萜烯和其他VOCs 占比分别为64.13%、9.63%、2.11%和24.13%. 由图3 可见:该研究将MEGAN 2.1 计算输出的37 种单萜烯分为8 类,分别为月桂烯、桧烯、柠檬烯、3-蒈烯、罗勒烯、β-蒎烯、α-蒎烯和其他单萜烯;将30 种倍半萜烯分为3 类,分别为金合欢烯、石竹烯和其他倍半萜烯.单萜烯排放量较高的化合物为α-蒎烯和β-蒎烯,分别占总单萜烯排放量的27.84%和15.85%;倍半萜烯排放量较高的化合物为金合欢烯和石竹烯为主,分别占倍半萜烯排放量的36.66%和24.30%.

图 3 我国夏季单萜烯和倍半萜烯的排放组成Fig.3 Emission composition of monoterpenes and sesquiterpenes in summer in China
各植被类型的BVOCs 排放量及其组成如表3 所示. 由表3 可见:BVOCs 排放量最多的是阔叶树,虽然阔叶树覆盖面积只占植被覆盖总面积的7.9%,但由于阔叶树种具有较高的异戊二烯排放速率,其对总BVOCs 排放量的贡献率最高,为56.01%;灌木次之,其对总BVOCs 排放量的贡献率为14.16%,灌木BVOCs 排放速率虽较低,但其叶生物量密度相对较高且覆盖面积占比(23.7%)较大;针叶树、农作物和草地的排放贡献率相当,分别占排放总量的7.9%、9.91%和10.31%,但三者覆盖面积差异较大,说明三者BVOCs 排放能力、叶生物量密度不同;针叶阔叶混交林的排放能力较强,但其面积只有2.13×104km2,因此对总BVOCs 排放量的贡献率仅为1.71%. 不同种类的BVOCs 来源构成有差异,异戊二烯主要来自阔叶树排放,占异戊二烯总排放量的80.71%,主要是由于阔叶树具有较高的异戊二烯排放速率,灌木的贡献率为13.76%,其余植被类型的贡献率小于5.53%;单萜烯主要来自针叶树(35.16%),草地次之(22.84%),灌木和阔叶树的排放贡献率相当,均为14%左右,农作物为10.12%;倍半萜烯和其他VOCs 的来源构成相近,草地和农作物贡献率较高,针叶树和灌木次之.不同植被类型的排放组成也不同,总的来说,异戊二烯是阔叶树、针叶阔叶混交林、灌木的高排放物种,其他VOCs 是针叶树、农作物、草地的高排放物种,此外单萜烯也是针叶树的高排放物种.

表 3 我国夏季6 种植被类型的BVOCs 排放量Table 3 BVOCs emissions of six vegetation types in summer in China 103 t
2.2.2 BVOCs 空间分布
BVOCs 排放具有明显的地域差异. BVOCs 的高排放地区分布在东北、华中、华东以及广西壮族自治区和云南省交界、云南省西部等地区,每网格排放量大于9 t. 由于异戊二烯对总BVOCs 排放贡献最高,BVOCs 排放总量的空间分布与森林(尤其是阔叶树)分布一致. 西部地区BVOCs 排放较低,尤其是西藏自治区和青海省,一方面由于该地区植被稀疏,另一方面该地区全年温度较低,而温度是影响植被排放的重要环境因素[42].
不同植被类型BVOCs 排放差异显著,其空间分布不仅与植被覆盖和叶生物量的分布有关,还与具有不同排放强度的植物组成有关,同时也与各地区气象条件有关. 总体来说,由于南方的温度和辐射全年较高,各植被类型的BVOCs 排放通量高于北方. 阔叶树BVOCs 排放主要集中在东北、华东和华中地区,如吉林省、黑龙江省、江西省、湖南省,这些省份每网格的排放通量大于5 t;其中,东北地区阔叶树蓄积量较大,并分布大量栎类、杨树及其他阔叶混交林等具有高异戊二烯排放强度的树种;华东地区阔叶树种组成较复杂,包含各种落叶、硬叶阔叶林以及阔叶混交林,部分地区种植了大量的竹子;其余大部分地区每网格排放量均小于100 g. 针叶树的高BVOCs 排放分布区域与阔叶树有所差异,主要分布在华东、华中、华南和西南地区,包括四川省、湖南省、云南省、广西壮族自治区、福建省、江西省等地区,这些地区每网格排放量大于2 t,该区域针叶树分布较密集,蓄积量较高,且分布的针叶树多具有高BVOCs 排放速率和叶生物量密度,如马尾松、云杉、铁杉. 针叶阔叶混交林排放的BVOCs 主要集中在东北地区. 农作物的BVOCs 排放与其种植密度和不同种类农作物的排放强度差异有关,集中在华北偏南、华东和华中地区,包括河南省、安徽省、湖北省、河北省、山东省等省份,这些省份农作物的年产量较高;此外,海南省每网格排放量大于500 g,该省种植了大量的橡胶,而橡胶是农作物中BVOCs 排放强度最高的,同时我国南部橡胶的种植面积也较大;河南省、山东省、黑龙江省、河北省的农作物产量较高,但叶生物量密度比浙江省、福建省等省份低,导致BVOCs 排放量较低. 草地的BVOCs 排放主要分布在华北、西北和西南地区,其中内蒙古自治区的草原分布最为广泛,排放量最高;南方部分地区分布有大面积的热性草丛,而热性草丛的BVOCs 排放能力较强. 灌木的BVOCs 排放则主要集中在华中、华南和西南地区,包括浙江省、湖北省、重庆市、贵州省、云南省西南部以及海南省,北方地区排放较低.
总体来说,阔叶树BVOCs 排放最高,灌木、针叶树次之,草地和农作物最低;青藏高原地区的植被分布稀疏,所以各类型植被排放的BVOCs 水平均较低.不同地区各植被类型的排放贡献也不同,吉林省、辽宁省、河南省、山东省、陕西省、黑龙江省等省份70%以上的BVOCs 来自阔叶树排放,上海市、天津市的BVOCs 排放主要来自于农作物,青海省、宁夏回族自治区、内蒙古自治区、西藏自治区等地区的草地贡献最高,重庆市和贵州省以灌木排放的BVOCs为主. 针叶树排放的BVOCs 中贡献最高的地区为四川省和西藏自治区.
2.2.3 不确定性分析
已有研究[25,32-33]中采用不同植被、气象数据、排放算法估算了我国BVOCs 的排放量. 将各排放清单中夏季(6—8 月)排放结果与笔者研究结果进行对比(见表4)发现,各研究结果存在较大差异,其研究结果均低于笔者研究. 导致差异的原因主要是清单使用的排放速率、叶生物量、气象参数、植被分布、排放算法等不同.

表 4 该研究与其他研究中我国BVOCs 排放清单对比Table 4 Comparison with other estimates of BVOCs emissions in China 1012 g
排放速率的选取影响排放清单的不确定性. 高超等[33]中使用的排放速率为MEGAN 3.1 模型的默认取值,缺少对我国本土植物的测量结果;闫雁等[25]采用传统的“排放强度分档取值法”确定排放速率,该方法中排放强度的分类、排放区间的确定以及特征值的选取理论依据均不明确,给排放速率的取值带来较大的不确定性. 笔者研究基于统计学方法确定了更详细的植被排放强度分类,并据此确定各植被的排放速率,降低了排放速率的不确定性. 因排放速率的确定方法和来源不同,故各研究排放速率的取值不同,如笔者研究中栎树、杨树、竹的异戊二烯排放速率均为70 μg(以C 计)/(g·h),而闫雁等[25]中取值分别为60、60 和0.5 μg(以C 计)/(g·h).
叶生物量的选取影响排放清单的不确定性. 闫雁等[25]采用平均叶生物量密度法,同一植被类型选取一个平均叶生物量密度值,未体现叶生物量的地区和时间差异. 高超等[33]选用MEGAN 3.1 模型默认的叶生物量密度. 笔者研究基于全国各省份植被蓄积量和产量估算叶生物量,体现了叶生物量地区和树龄的差异,但仍存在一定的不确定性,其主要来源于统计数据和分配模型的不确定性[50]. 中国森林和草地资源统计的平均误差分别低于5%和10%[51]. 就全国而言,由生物量分配模型估算得到的森林叶生物量的误差低于3%,但在区域尺度上误差可能更高.
气象参数影响排放清单的不确定性. 闫雁等[25]利用全国有限观测站点的实测数据插值得到气象分布,给估算带来较大的误差,且数据密度、数据分布和空间异质性均会影响插值的精度. 高超等[33]和笔者研究应用中尺度气象模式WRF 模拟高时空分辨率的气象分布,提高了排放清单的准确性.
植被分布影响排放清单的不确定性. Tie 等[32]采用USGS 中的1 km×1 km 土地覆盖数据确定20 种植被类型. 闫雁等[25]利用《中华人民共和国植被图(1∶1 000 000)》提供的植被分布,但年份较久远,且仅按照植被亚类将植被分为九大类,未研究细分树种的排放,各类植被的覆盖面积与笔者研究不同,其中草地覆盖面积比笔者研究结果少约150×104km2.
排放算法影响排放清单的不确定性. 闫雁等[25,32]采用的排放算法未反映冠层内环境的影响. 高超等[33]和笔者研究应用MEGAN 排放模式估算BVOCs排放量,考虑了冠层内温度和辐射等环境因素的分布对排放量的影响,排放清单的准确性更高. 但笔者研究中忽略了土壤水分和CO2等环境因素对排放的影响,这将带来一定的误差.
3 结论
a)广玉兰、马尾松、油松、雪松、侧柏、小麦、紫马唐和大叶黄杨的单萜烯排放速率分别为16.169、40.475、1.218、13.995、0.109、0.278、0.565 和0.048 μg/(g·h). 针叶树的单萜烯排放速率最高,其次是阔叶树,草地和农作物排放速率较低,灌木排放速率最低.
b)广玉兰排放的单萜烯以α-蒎烯为主,油松以排放顺式-β-罗勒烯为主,雪松和大叶黄杨以排放柠檬烯为主,侧柏以排放香桧烯为主,马尾松、小麦和紫马唐排放的单萜烯以异松油烯为主. 针叶树以排放异油松烯和柠檬烯为主,阔叶树以排放α-蒎烯和柠檬烯为主,农作物和灌木以排放异松油烯和柠檬烯为主,草地以排放异松油烯和反式-β-罗勒烯为主.
c) 2018 年我国夏季BVOCs 总排放量为33.6×1012g,其中异戊二烯贡献率最高,占总排放量的64.13%,其他VOCs 贡献率次之,占24.13%,单萜烯和倍半萜烯分别占9.63%和2.11%. 单萜烯中排放贡献率最高的是α-蒎烯,倍半萜烯中排放贡献率最高的是金合欢烯.
d)阔叶树、针叶树、针叶阔叶混交林、农作物、草地、灌木分别排放18.8、2.7、0.6、3.3、3.5 和4.8 ×1012g BVOCs. 异戊二烯是阔叶树、针叶阔叶混交林、灌木的高排放物种,其他VOCs 是针叶树、农作物、草地的高排放物种,此外单萜烯也是针叶树的高排放物种.
e)我国BVOCs 的高排放地区分布在东北、华中、华东以及广西壮族自治区和云南省交界地区、云南省西部等地区,BVOCs 排放总量的空间分布与森林分布一致. 不同类型植被的BVOCs 排放差异较显著,与各地区植被覆盖及其植物分布、气象条件的差异有关.
