厚朴酚对断奶仔猪生长性能、肝脏抗氧化功能及脂代谢的影响
2022-06-10屈圣富梅华迪李贞明李袁飞马现永高凤仙
屈圣富 田 琦 梅华迪 李贞明 李袁飞 马现永,3 高凤仙 余 苗,3*
(1.广东省农业科学院动物科学研究所,农业部华南动物营养与饲料重点实验室,畜禽育种国家重点实验室,广东省畜禽育种与营养研究重点实验室,广东省畜禽肉品质量安全控制与评定工程技术研究中心,广州510640;2.湖南农业大学动物科学技术学院,长沙410128;3.岭南现代农业科学与技术广东省实验室茂名分中心,茂名525000)
随着生猪养殖业的迅速发展,为降低仔猪生产的经济成本和减少母猪向仔猪垂直传染疾病的几率,在生产中常采用仔猪早期断奶技术。然而,由于仔猪消化和免疫功能发育不成熟,早期断奶极易引起仔猪出现氧化应激和脂代谢功能紊乱,进而引发诸多疾病[1-3],最终阻碍仔猪的生长发育。肝脏是脂代谢的中枢器官,同时也是应对氧化应激的靶向和效应器官,可直接影响宿主动物生长潜能的发挥。当肝脏出现氧化应激时,肝脏中会出现过量的自由基,破坏肝细胞的生物膜以及功能,造成肝脏器官的损伤,影响肝脏脂代谢功能的稳定性,继而诱导肝脏出现脂代谢功能紊乱[4-5]。此外,脂代谢功能紊乱亦会导致脂质过氧化,产生大量自由基,破坏肝细胞的生物膜以及功能,造成肝细胞的损伤,最终导致肝脏出现氧化应激[5-6]。因此,如何通过营养手段来调节断奶仔猪肝脏氧化功能和脂代谢状况已成为我国当前生猪养猪业亟需解决的问题。
厚朴酚是从中药厚朴的茎皮中分离出的一种联苯酚类化合物,具有抗炎、抗氧化、改善脂代谢等生理功能[7-8]。Liu等[9]在酒精性肝损伤小鼠模型研究中发现,厚朴酚可显著提高肝脏超氧歧化酶(SOD)以及谷胱甘肽过氧化酶(GSH-Px)的活性,缓解氧化应激损伤并保护肝脏。在肉鸭上的研究发现,饲粮中添加200和300 mg/kg厚朴酚可提高临武鸭的生长性能,提高血清中SOD活性、总抗氧化能力(T-AOC)以及肝脏中过氧化氢酶(CAT)活性,降低血清中甘油三酯和总胆固醇含量与谷草转氨酶活性[10-11]。然而,当前厚朴酚在断奶仔猪上的应用研究相对较少,且对其生长性能、肝脏抗氧化功能和脂代谢的研究还未见系统报道。因此,本试验拟以断奶仔猪为试验对象,探究厚朴酚对断奶仔猪生长性能、肝脏抗氧化功能和脂代谢的影响,并进一步探究其影响的机制,以期为厚朴酚的饲料开发和断奶仔猪绿色生态健康养殖提供理论依据。
1 材料与方法
1.1 试验材料
试验用厚朴酚购自上海某生化科技股份有限公司,纯度≥98%。
1.2 试验设计及饲粮
试验选取体重[(7.91±0.00) kg]相近且健康的21日龄“杜×长×大”三元杂交断奶仔猪144头,随机分为3组,每组8个重复,每个重复6头猪。3组仔猪分别饲喂基础饲粮(对照组)、基础饲粮+0.02%厚朴酚(0.02%厚朴酚组)、基础饲粮+0.04%厚朴酚(0.04%厚朴酚组)。饲养试验为期42 d,其中预试期7 d,正试期35 d。试验用基础饲粮参照NRC(2012)仔猪营养需要配制,仔猪7~11 kg阶段(试验第1~19天)与11~25 kg阶段(试验第20~35天)基础饲粮组成及营养水平见表1。
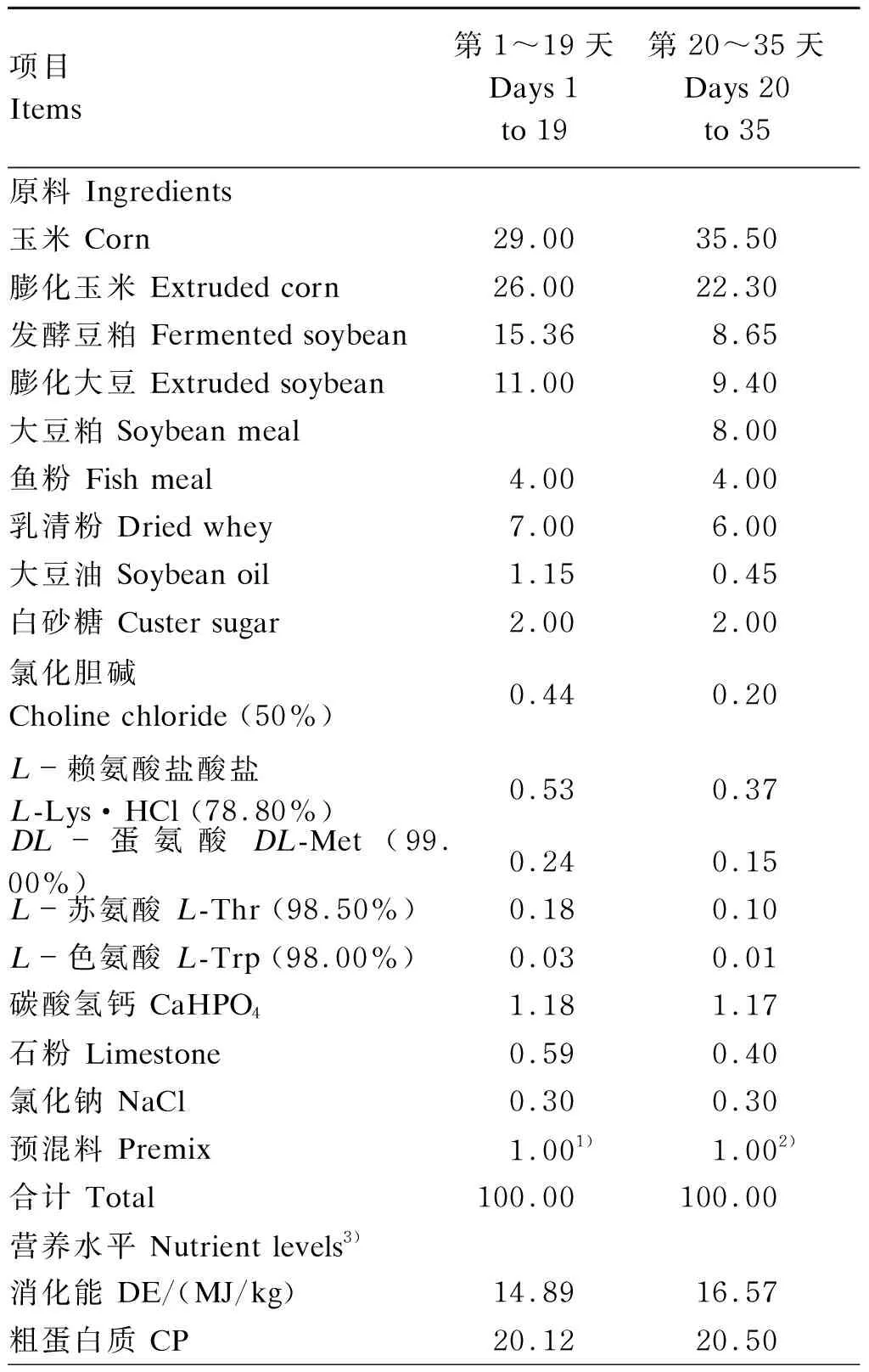
表1 不同阶段基础饲粮组成及营养水平(饲喂基础)

续表1项目 Items第1~19天Days 1 to 19第20~35天Days 20 to 35赖氨酸 Lys1.551.17蛋氨酸+半胱氨酸 Met+Cys0.900.69苏氨酸 Thr0.970.73色氨酸 Trp0.270.19
1.3 饲养管理和样品采集
饲养试验在广东省农业科学院动物科学研究所试验基地进行。试验期内,猪只分组后均饲养于同一圈舍内,并分栏饲养在1.2 m×2.1 m大小且带有塑料漏风地板的高保育床上。试验期间猪只自由饮水和采食,日常饲养管理、消毒、常规免疫按照基地动物饲养要求进行。于试验结束时,从每栏内选取1头接近平均体重的仔猪,进行麻醉后颈静脉放血致死,分离其肝脏组织样品置于冻存管中,液氮冷冻后置于-80 ℃冰箱保存备用。
1.4 测定指标及方法
1.4.1 生长性能和腹泻
正式试验期间,每天08:00开始投喂,以栏为单位,记录每日饲喂量以及每日余料量,计算平均日采食量(average daily feed intake,ADFI)。在正式试验第1、19、35天以及试验结束当天禁食12 h后对每栏的仔猪进行空腹称重,计算平均日增重(average daily gain,ADG)和料重比(feed/gain,F/G)。计算公式如下:
ADFI=每个重复正试期总采食量/
(正试期试验天数×该重复试验猪头数);
ADG=(末重-初重)/(正试期试验天数×
该重复试验猪头数);
F/G=ADFI/ADG。
每天09:00逐只观察各重复仔猪的粪便状况,同时结合精神状态、肛门有无红肿及水样粪便痕迹等,记录仔猪腹泻头数和腹泻评分,腹泻评分标准见表2。腹泻率和腹泻指数的计算公式如下:
腹泻率(%)=[每个重复腹泻仔猪头数/
(每个重复仔猪头数×天数)]×100;
腹泻指数=每个重复仔猪腹泻评分之和/
(每个重复仔猪头数×天数)。

表2 腹泻评分标准
1.4.2 肝脏抗氧化和脂代谢指标
取0.1 g肝脏组织样品和0.9 mL生理盐水,将其制成10%的匀浆液,离心,移取上清液,测定上清液中抗氧化指标(总超氧化物歧化酶、谷胱甘肽过氧化物酶、过氧化氢酶活性、丙二醛含量及总抗氧化能力)和脂代谢指标(甘油三酯、胆固醇、高密度脂蛋白、低密度脂蛋白含量)。上述指标测定所用试剂盒采购于南京建成生物工程研究所,严格按照说明书进行测定操作。
1.4.3 肝脏抗氧化和脂代谢相关基因mRNA相对表达量
采用Trizol试剂(TaKaRa,大连)提取肝脏总RNA,具体提取步骤参照Yu等[13]的方法。采用核酸蛋白检测仪(Nanodrop 2000,Thermo Fisher Scientific,美国)测定RNA的浓度和纯度,260和280 nm处吸光度(OD)比值应介于1.9~2.1。随后采用1%甲醛变性凝胶电泳对RNA完整性进行检测。选取RNA质量较高的样品,使用PrimerScriptTMRT Reagent Kit(TaKaRa,大连)逆转录试剂盒进行cDNA合成,具体操作步骤参照说明书,获得的cDNA置于-20 ℃保存备用。以cDNA为模板,分别选取7个与肝脏抗氧化功能和4个与肝脏脂代谢功能相关的基因进行荧光定量PCR,相关特异性引物分别见表2和表3。采用实时定量PCR仪(CFX96 Real-Time PCR System,Bio-RadLaboratories,美国)进行检测,PCR反应体系和反应条件均参照本课题前期已发表的相关文献[14-15]。定量结果以β-肌动蛋白(β-actin)作为内参基因,每个样品测定重复3次。采用2-△△Ct法分析相关基因的mRNA相对表达量[16]。

表3 抗氧化相关基因的引物序列

表4 脂代谢相关基因的引物序列
1.5 数据处理
试验数据首先用Excel 2016进行初步处理,再用SPSS 26.0软件中的one-way ANOVA模块进行显著性分析,差异显著者进一步采用Tukey’s法进行多重比较。试验结果采用“平均值±标准误”表示,P<0.05视为差异显著,P<0.01视为差异极显著。
2 结果与分析
2.1 厚朴酚对断奶仔猪生长性能和腹泻的影响
厚朴酚对断奶仔猪生长性能和腹泻的影响见表5。与对照组相比,饲粮添加0.04%厚朴酚显著提高断奶仔猪第1~19天和第20~35天的ADG(P<0.05),显著降低第1~19天和第20~35天的F/G(P<0.05)。0.02%厚朴酚组和0.04%厚朴酚组的腹泻率和腹泻指数均显著低于对照组(P<0.05)。饲粮添加厚朴酚有降低断奶仔猪第1~19天ADFI的趋势(P=0.062)。0.02%厚朴酚组断奶仔猪第1~19天、第20~35天的ADG以及第1~19天、第20~35天和第1~35天的F/G相比与对照组和0.04%厚朴酚组均无显著差异(P>0.05)。此外,各组间第19和35天的体重、第20~35天和第1~35天的ADFI、第1~35天的ADG均无显著差异(P>0.05)。

表5 厚朴酚对断奶仔猪生长性能的影响
2.2 厚朴酚对断奶仔猪肝脏抗氧化指标的影响
厚朴酚对断奶仔猪肝脏抗氧化指标的影响见表6。与对照组相比,饲粮添加0.02%和0.04%厚朴酚显著提高断奶仔猪肝脏中GSH-Px和T-SOD活性(P<0.05),而0.02%厚朴酚组和0.04%厚朴酚组间的上述指标差异不显著(P>0.05)。与对照组和0.02%厚朴酚组相比,饲粮添加0.04%厚朴酚显著增加了肝脏中CAT活性(P<0.05),而对照组和0.02%厚朴酚组间该指标差异不显著(P>0.05)。此外,各组间肝脏中T-AOC和MDA含量差异不显著(P>0.05)。

表6 厚朴酚对断奶仔猪肝脏抗氧化指标的影响
2.3 厚朴酚对断奶仔猪肝脏脂代谢指标的影响
由表7可知,与对照组相比,饲粮添加0.02%和0.04%厚朴酚显著降低断奶仔猪肝脏中胆固醇含量(P<0.05),饲粮添加0.04%厚朴酚显著降低肝脏中高密度脂蛋白含量(P<0.05)。0.02%厚朴酚组肝脏中高密度脂蛋白含量与对照组和0.04%厚朴酚组均差异不显著(P>0.05)。此外,不同饲粮处理对肝脏中甘油三酯和低密度脂蛋白含量无显著影响(P>0.05)。

表7 厚朴酚对断奶仔猪肝脏脂代谢指标的影响
2.4 厚朴酚对断奶仔猪肝脏抗氧化相关基因表达的影响
厚朴酚对断奶仔猪肝脏抗氧化相关基因表达的影响如图1所示。与对照组相比,饲粮添加0.04%厚朴酚显著提高肝脏中GPx1和GR的mRNA相对表达量(P<0.05),而0.02%厚朴酚组上述2个基因的mRNA相对表达量与对照组和0.04%厚朴酚组相比差异不显著(P>0.05)。与对照组和0.02%厚朴酚组相比,饲粮添加0.04%厚朴酚显著提高肝脏中Keap1的mRNA相对表达量(P<0.05)。与对照组和0.02%厚朴酚组相比,饲粮添加0.04%厚朴酚极显著提高肝脏中Nrf2、CAT和SOD1的mRNA相对表达量(P<0.01)。此外,0.04%厚朴酚组肝脏中GST的mRNA相对表达量极显著高于对照组(P<0.01),显著高于0.02%厚朴酚组(P<0.05)。
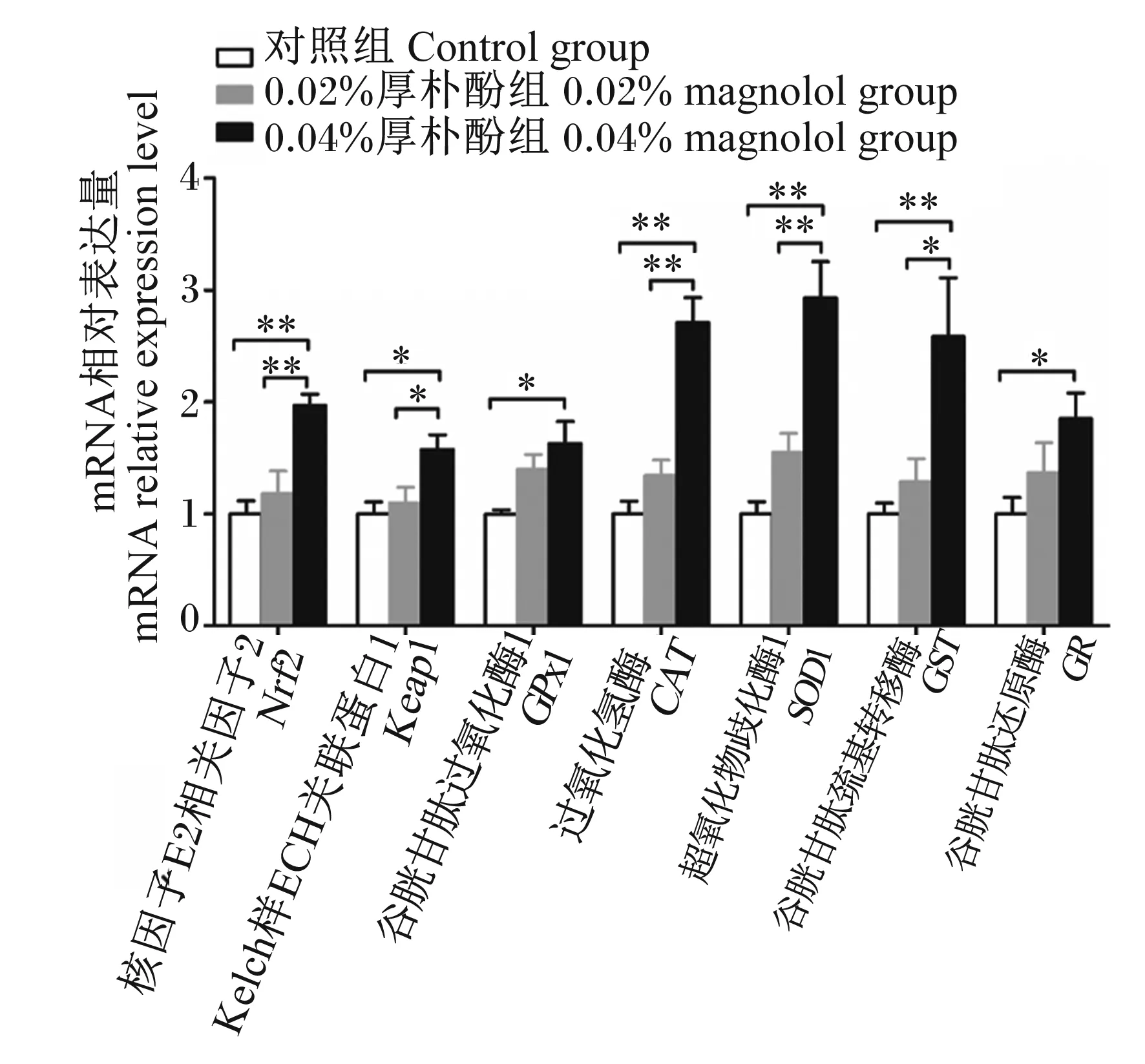
*表示差异显著(P<0.05),**表示差异极显著(P<0.01),无*表示差异不显著(P>0.05)。下图同。
2.5 厚朴酚对断奶仔猪肝脏脂代谢基因表达的影响
厚朴酚对断奶仔猪肝脏脂代谢基因表达的影响如图2所示。与对照组相比,饲粮添加0.04%厚朴酚显著降低肝脏中脂肪酸合成酶(FAS)和甘油三酯脂酶(ATGL)的mRNA相对表达量(P<0.05),而0.02%厚朴酚组上述基因的mRNA相对表达量与对照组和0.04%厚朴酚组相比差异不显著(P>0.05)。此外,不同饲粮处理对肝脏中过氧化物酶体增殖物激活受体γ(PPARγ)、胆固醇调节元件结合蛋白(SREBP)和硬脂酰辅酶A去饱和酶(SCD)的mRNA相对基因表达量无显著影响(P>0.05)。

图2 厚朴酚对断奶仔猪肝脏脂代谢相关基因表达的影响
3 讨 论
3.1 厚朴酚对断奶仔猪生长性能和腹泻的影响
诸多研究已表明厚朴酚可改善动物的生长性能。在肉鸭上的研究表明,饲粮添加200 mg/kg厚朴酚可使肉鸭的ADG提高4.24%[11];在肉鸡上的研究表明,饲粮添加200 mg/kg厚朴总酚可使肉鸡的ADG提高1.41%,料重比降低2.88%[17]。本试验也发现,饲粮添加0.04%厚朴酚可显著提高断奶第1~19天和第20~35天的ADG,显著降低第1~19天和第20~35天的F/G,这与上述研究结果相似。前人研究发现,厚朴酚可增加空肠和回肠的绒毛高度及绒隐比,提高回肠营养代谢相关基因[亚精胺-精胺乙酰转移酶(SAT1)与碱性鞘磷脂酶7(ENPP7)]和紧密连接蛋白基因[封闭蛋白-1(Claudin-1)和封闭蛋白-5(Claudin-5)]的表达,同时还可增加回肠微生物的多样性和有益微生物乳酸杆菌的丰度,从而促进肠黏膜细胞发育、维护肠黏膜结构及其屏障功能,进而维持肠道内环境稳态,最终维系宿主肠道健康[18]。在临武鸭上的研究发现,厚朴酚可提高肝脏中抗氧化酶的活性,改善回肠黏膜形态[19]。断奶仔猪各方面机能未完全发育成熟,自身抗应激能力较弱,易受到氧化应激的影响,导致机体代谢紊乱、饲料利用率低、腹泻等症状,严重影响了仔猪的生长潜力。而在本试验中,饲粮添加0.04%厚朴酚可提高断奶仔猪肝脏中抗氧化酶的活性,表明其可有效抑制氧化应激,改善仔猪的健康状态,有益于仔猪生长。因此,本试验中,厚朴酚促生长的原因一方面可能是维系了仔猪的肠道健康;另一方面可能是提高了断奶仔猪机体的抗氧化应激能力,进而促进营养物质的消化吸收,最终提高断奶仔猪的生长性能,但其具体作用机制还需进一步研究。
腹泻是阻碍仔猪快速生长与影响仔猪健康的主要原因之一。本研究发现,饲粮添加0.04%厚朴酚可降低断奶仔猪的腹泻率和腹泻指数,表明厚朴酚的添加降低了仔猪腹泻,有利于仔猪健康生长。前人研究发现,厚朴酚可通过抑制大肠杆菌、变形杆菌、金黄色葡萄球菌等细菌的增殖,调节胃肠蠕动、阻碍钙离子通道、抑制炎症因子的产生与释放,来发挥抗腹泻作用[20]。这可能是本试验中厚朴酚降低仔猪腹泻的作用途径。
3.2 厚朴酚对断奶仔猪肝脏抗氧化功能的影响
肝脏作为机体营养物质代谢和氧化还原反应的重要场所,在脂类代谢过程中可生成大量的自由基[21]。维系机体的氧化平衡状态对机体正常的生理功能是有益的,而一旦自由基未及时清除,氧化平衡被打破,过量的自由基就会改变细胞膜结构,诱导肝脏发生过氧化损伤,继而导致肝脏功能紊乱,影响机体营养物质代谢[22-23]。肝脏中自由基的清除主要依靠其自身分泌的各种抗氧化酶,如T-SOD可清除超氧阴离子自由基(O2-·),GSH-Px可将过氧化氢(H2O2)转化为水,将脂质过氧化物转化为醇[24]。肝脏抗氧化酶活性高低也是动物机体免疫水平高低的重要体现[25]。研究发现厚朴酚可以提高肉鸭肝脏中抗氧化酶活性,继而提高机体抗氧化能力[10]。本试验结果显示,饲粮添加0.04%厚朴酚显著提高断奶仔猪肝脏中GPSH-Px、T-SOD和CAT活性。这表明厚朴酚可能通过提高断奶仔猪肝脏抗氧化酶的活性来维持肝脏中的氧化平衡状态,减少氧化应激对肝脏功能的影响,提升机体的免疫力,继而使机体营养物质代谢有条不紊地进行,最终使仔猪生长性能得到改善。
Nrf2-抗氧化原件(ARE)信号通路是机体抗氧化应激的主要防御机制,Keap1是转录因子Nrf2相关的细胞质蛋白,当Keap1受到氧化应激的刺激时,会导致Nrf2的释放和核转位,随后Nrf2进入到细胞核内结合ARE序列,激活各类抗氧化酶基因的表达[26-27]。其中SOD1基因可以参与细胞内超氧化物歧化酶(SOD)的合成过程,同时,CAT基因可以参与细胞内CAT的合成过程[28]。并且,SOD1和CAT基因的mRNA相对表达量分别与SOD和CAT的活性呈正相关[29]。此外,GPx1、GR、GST基因可以调控谷胱甘肽氧化还原系统中对应的抗氧化酶活性,3种基因表达的上调可以分别提高各自对应抗氧化酶的活性[30-32]。Liu等[9]在酒精性肝损伤小鼠模型的研究中发现,厚朴酚显著提高肝脏中Nrf2信号通路的活性,进而提高抗氧化酶SOD以及GSH-Px的活性,缓解氧化应激损伤并保护肝脏。Lin等[11]在肉鸭上的研究发现,200 mg/kg厚朴酚提高了肝脏中Nrf2、CAT、SOD1、锰超氧化物歧化酶(MnSOD)、GPx1、谷胱甘肽过氧化物酶4(GPx4)、GST和血红素氧合酶-1(HO-1)的mRNA相对表达量。本试验研究发现,饲粮添加0.04%厚朴酚显著提高了断奶仔猪肝脏中Nrf2、Keap1、CAT、SOD1、GPX1、GR、GST的mRNA相对表达量。这表明厚朴酚通过激活Keap1-Nrf2-ARE信号通路,进而提高肝脏中CAT、SOD1、GPx1、GR、GST基因的相对表达量,增强肝脏中抗氧化酶的活性,最终提高断奶仔猪肝脏的抗氧化功能。
3.3 厚朴酚对断奶仔猪肝脏脂代谢的影响
肝脏是脂肪代谢的中枢器官,在脂肪的合成和分解、能量代谢、抗氧化过程中起着重要作用[33]。因此,通过研究肝脏脂代谢产物含量以及脂代谢相关基因表达的变化可了解机体脂代谢情况[34-35]。同时,肝脏的脂代谢与肝脏的抗氧化能力密切相关,脂代谢的稳定是肝脏抗氧化能力强的重要体现[34]。胆固醇是一种合成多种类固醇的脂,肝脏中胆固醇含量升高,容易导致肝脏脂肪沉积,从而加重肝脏负荷并诱导肝脏出现氧化损伤,降低肝脏的生理机能,增加脂肪肝疾病的发生几率[36],进而影响动物的生长性能。高密度脂蛋白是脂肪运输蛋白,可以运输血液中胆固醇到肝脏进行代谢,其在肝脏中的含量可以反映肝脏中胆固醇的含量[37]。本试验中,饲粮添加0.04%厚朴酚显著降低了肝脏中胆固醇和高密度脂蛋白的含量,表明厚朴酚可以减少肝脏中脂肪沉积,改善肝脏脂代谢,这有利于维护肝脏的健康,促进仔猪的生长发育。
调控脂代谢的相关基因有FAS、ATGL和SCD等。其中FAS是提高脂肪酸合成速度的酶[38]。ATGL是甘油三酯的水解酶,可以把储存的甘油三酯水解成游离的脂肪酸[39-40]。在脂肪细胞分化前中期,FAS的mRNA相对表达量明显增加,提高脂肪沉积速度,而当脂肪成熟时,ATGL的mRNA相对表达量也会明显提高,表明细胞内脂肪分解也开始活跃[41-42]。因此,脂肪合成和分解处于一个动态平衡,当脂肪积累超出负荷时,脂质分解增加,引起脂肪酸浓度升高、高甘油三脂症、高胆固醇等疾病,导致脂代谢紊乱。在减少脂肪酸合成的同时,抑制脂肪的分解更有利于维持脂代谢的平衡[41]。有研究显示,抑制FAS和ATGL的mRNA相对表达量和活性可有效抑制脂解反应,减少游离脂肪酸含量,改善脂代谢[43-44]。本试验研究发现,添加0.04%厚朴酚显著降低断奶仔猪肝脏中FAS和ATGL的mRNA相对表达量,表明厚朴酚可以降低断奶仔猪肝脏中脂肪的合成以及甘油三酯的水解,从而减少肝脏中游离脂肪酸释放,起到降脂和改善脂肪代谢的作用。
4 结 论
综上所述,饲粮添加0.04%厚朴酚可提高断奶仔猪的生长性能;可通过Nrf2信号通路增加断奶仔猪肝脏抗氧化能力,进而维持体内的氧化平衡,缓解断奶造成的氧化应激;同时可通过调控肝脏中FAS和ATGL基因的表达,促进肝脏中脂质分解,抑制脂质的生物合成,进而改善肝脏的脂代谢,维护肝脏的健康。
