细胞外基质修饰的电纺纤维对施万细胞及神经生长的影响
2022-06-09管延军许文静李超超孟繁琪杨博尧张铁元任致奇刘修志仪鹏朝
管延军,许文静,张 健,李超超,孟繁琪,3,杨博尧,张铁元,任致奇,4,刘修志,仪鹏朝,王 玉,彭 江
1 解放军医学院,北京 100853;2 解放军总医院第四医学中心 骨科医学部研究所,北京 100853;3 北京大学人民医院 脊柱外科,北京 100044;4 解放军总医院第一医学中心 神经外科,北京 100853;5 66168 部队医院 门诊部,北京 101301
细胞外基质(extracellular matrix,ECM)为其周围定居的细胞提供机械支持和表型调控的生物化学信号[1]。在组织工程与再生医学发展过程中,再生微环境一直是研究人员关注的焦点,生物支架材料的设计趋向于再现天然基质特征,力求还原组织器官的最初(损伤前)微环境,即目标相关的机械线索以及生物引导信号,以此达到再生的目的[2]。周围神经损伤后的核心再生环节包括施万细胞的迁移以及轴突生长引导[3-4]。因此神经再生材料的设计思路就针对于模拟再生微环境。施万细胞(Schwann cells,SCs)是周围神经组织中的胶质细胞,参与有髓神经纤维的髓鞘包裹以及神经纤维的营养支持;在损伤的神经组织中,施万细胞是最早一批响应轴突损伤的细胞。在损伤发生后,施万细胞重编程并形成邦格氏带,伴随其分泌的可溶性蛋白(促神经生长因子)以及不可溶性蛋白(细胞外基质)作为引导轴突生长的桥梁[5]。本研究组装了一种改性静电纺丝材料,即在传统的取向聚己内酯(polycaprolactone,PCL)-丝素蛋白(silk fibroin,SF)纳米纤维上沉积大鼠SCs 的ECM。取向纳米纤维在结构上再现神经的基底膜结构,而SCs-ECM 则提供轴突生长的信息结合位点。通过体外大鼠背根神经节(dorsal root ganglion,DRG)培养评估这种策略对神经轴突生长的影响。
材料与方法
1 实验动物 大鼠原代施万细胞、背根神经节组织分别取自72 时龄、12 时龄的SD 乳鼠(雌雄不限),各5 只,均购自北京科宇动物养殖中心。本实验经解放军总医院实验动物伦理委员会审批通过(批准号:2016-x9-07)。
2 主要实验设备及材料 体视显微镜(Nikon DSRi2);静电纺丝设备(北京永康乐业 ET-2531);真空冻干机(河南兄弟仪器 LGJ-18);荧光显微镜(Nikon Ni-U);相差显微镜(Olympus 1X53);激光共聚焦显微镜(Zeiss LSM780);核酸提取分析仪(QIAGEN QIAcube HT);荧光分光光度仪(infinite 200Pro);核酸定量检测试剂盒(invitrogen P7589);DMEM/F-12(Gibco 11965084);磷酸盐缓冲液(CORNING 21-031-CV);胎牛血清 (Gibco 10099-141);EGF-D(R&D 396-HB);毛喉素(Sigma F6886);B-27 添加因子(gibco 17504-044);青-链霉素双抗(Gibco 15070063);Ⅰ型胶原酶(Invitrogen 17018-029);胰酶(Gibco 15050065);阿糖胞苷(Sigma C6645);Glutamax(Gibco 35050061);L-抗坏血酸(Wako 014-04801);聚-D-赖氨酸(sigma P6407);明胶(Sigma G7041);聚己内酯(Sigma 440744);蚕丝蛋白(美伦生物 MB4792);六氟异丙醇(阿拉丁 H1-7501);Triton-X100(Sigma T8787);DNA 酶Ⅰ(Sigma D5025);氨水(北京化工厂);封闭用山羊血清(索莱宝 SL038);兔来源Fibronectin 一抗(Abcam ab2413);兔来源 S100β 一 抗 (Abcam ab52642);小鼠来源NF-200 一抗(Sigma N0142);山羊抗兔-Alexa Fluor ® 594(Abcam ab150080);山羊抗小鼠-Alexa Fluor ®488(Abcam ab150117);DAPI 染色液(四正柏生物 FXP 139-100)。
3 大鼠原代施万细胞的提取、纯化与增殖培养施万细胞基础培养基:DMEM/F-12、10%胎牛血清、2 mm/L GlutaMAX-I、1×双抗。施万细胞纯化培养基:施万细胞基础培养基添加10 µm/L 阿糖胞苷。施万细胞完全培养基:施万细胞基础培养基添加2 µm Forskolin、10 ng/mL EGF-D。在无菌环境中,体视显微镜下获取5 只乳鼠的全部坐骨神经(1 cm),以DF-12 冲洗血迹并离心,加入10 mL胰酶/Ⅰ型胶原酶/DF-12 混合液后,在37℃下消化30 min;离心,弃上清,加入4 mL 施万细胞基础培养基,枪头吹打10~20 次,至无明显组织块,转移至 35 mm2培养瓶中,于37℃,5% CO2环境下培养。24 h 后更换施万细胞纯化培养基以抑制并杀死成纤维细胞,继续培养72 h。之后更换施万细胞完全培养基以增殖施万细胞。细胞每融合至90%左右进行传代,P2~P3 代的施万细胞用于细胞外基质的获取。
4 静电纺丝材料制备 配制聚己内酯-丝素蛋白(PCL-SF)纺丝溶液:将0.8 g 聚己内酯、丝素蛋白分别溶解于10 mL 六氟异丙醇中,室温下搅拌过夜形成均一稳定的溶液。将制得的8%聚已内酯/丝素蛋白纺丝溶液1∶1 混匀,吸入注射器装入电纺丝供给装置,通过流量泵调控电纺丝的流量供给速度为0.166 mm/min。通过高压发生器给电纺丝过程提供电压,施加的正电压为15.00 kV,负电压为-5.00 kV。将转速为2 800 r/min 的滚筒与接地端相连,覆盖锡箔纸,作为纺丝收集基板。
5 施万细胞ECM 修饰的电纺丝材料制备 将电纺膜裁剪并固定于2.2 cm×2.2cm 玻片,辐照灭菌后置于六孔板,以0.2%明胶包被4 h,PBS 冲洗3 次;胰酶消化增殖的SCs,以8 000/cm2的密度种植于电纺材料表面。待细胞达到100%融合后,更换添加50 µm/L L-抗坏血酸完全培养基以刺激细胞外基质分泌,刺激时间为12 d,每3 d 更换1 次培养基。为了获得无细胞材料,将培养基弃去并用PBS 冲洗,在脱细胞缓冲液中(0.5% Triton X-100 和20 mm/L 氨水溶解在PBS 中),37℃下孵育5 min,然后用100 U/mL DNA 酶Ⅰ在37℃下处理2 h。去细胞材料用PBS 洗3 次以去除残留的DNA 酶,在4℃下保存。
6 去细胞材料DNA 残留检测 为检测脱细胞效率,脱细胞和未经脱细处理的复合材料经真空冻干机冻干24 h 后,称重并经过核酸提取分析仪程序提取总DNA,按照核酸定量检测试剂盒操作说明进行定量,于荧光分光光度仪以激发光波长480 nm、发射波长520 nm 读取荧光强度数值;绘制标准曲线并计算DNA 含量。该实验重复3 次。
7 复合材料的扫描电子显微镜检测 为验证此策略是否成功将SCs-ECM 与PCL-SF 纳米纤维复合,通过扫描电镜直接观察复合情况。脱细胞处理后的样本经4%戊二醛溶液固定后,冲洗并与无ECM 复合的电纺膜一同干燥、喷金、上机观察拍照。
8 复合材料的免疫荧光染色 SCs-ECM分泌后,经脱细胞处理后的与未处理的复合材料经4%多聚甲醛溶液固定10 minPBS 冲洗,之后以封闭用山羊血清工作液孵育30min,兔来源Fibronectin 一抗4℃孵育过夜,第2 天以PBS 冲洗3 次后,取山羊抗兔-Alexa Fluor ®594 二抗避光孵育2 h,冲洗3 次,水性封片剂封片,至荧光显微镜观察。
9 大鼠DRG 的种植 脱细胞处理后的复合材料与单纯PCL-SF 电纺材料在种植前使用多聚赖氨酸工作液37℃下包被3 h,PBS 冲洗3 次备用。取生后12 h 乳鼠1 只脱颈处死,浸泡于75%乙醇中消毒15 min。将乳鼠俯卧位固定于蜡台,体视显微镜下剪开背部正中皮肤、皮下及肌层,暴露脊柱下颈段至腰段,完整剥离并取出脊柱,置于DMEM/F-12 中洗去血迹。用显微镊从正中将脊柱剪开,去除脊髓,暴露两侧椎间孔,于两侧椎间孔处取出各节段DRG,剥除DRG 外膜,接种于复合电纺材料和单纯电纺材料表面上,加2.5 mL DRG 培养液(DRG 培养液配方:DMEM/F-12、2 mm GlutalMAX-I、1×双抗、1×B-27 因子),置于37℃、5% CO2环境下培养7 d。
10 DRG 免疫荧光染色及轴突长度测量 去除复合电纺材料和单纯电纺材料表面培养的DRG 培养液,PBS 清洗后以4%多聚甲醛固定15 min。再以PBS 清洗,山羊血清封闭30 min,取兔来源S100β、小鼠来源NF-200 一抗稀释,4℃孵育过夜。第2 天以PBS 冲洗3 次后,取山羊抗兔-Alexa Fluor®594、山羊抗小鼠-Alexa Fluor®488 二抗避光孵育2 h,冲洗3 次,水性封片剂封片,至共聚焦显微镜观察拍摄全景图片。并用Image Pro Plus 软件分析计算各组DRG 轴突长度,取每个DRG 最长的轴突计算平均长度,每组随机选取6 个DRG 进行分析。
11 统计学方法 采用GraphPad Prism 7.0 进行统计学分析以及绘图,计量资料以±s表示,两两比较采用Student'st检验,P<0.05 为差异有统计学意义。
结果
1 大鼠施万细胞纯化培养的情况 经阿糖胞苷纯化后的施万细胞经传代后镜下观察呈细长纺锤形且细胞核折光性强,而少见宽大的核无折光性成纤维细胞(图1A),针对SCs 表面特异性标记蛋白S100β 的免疫荧光染色(图1B),可见融合的SCs之间未见成纤维细胞掺杂,提取的细胞可用于后续实验。
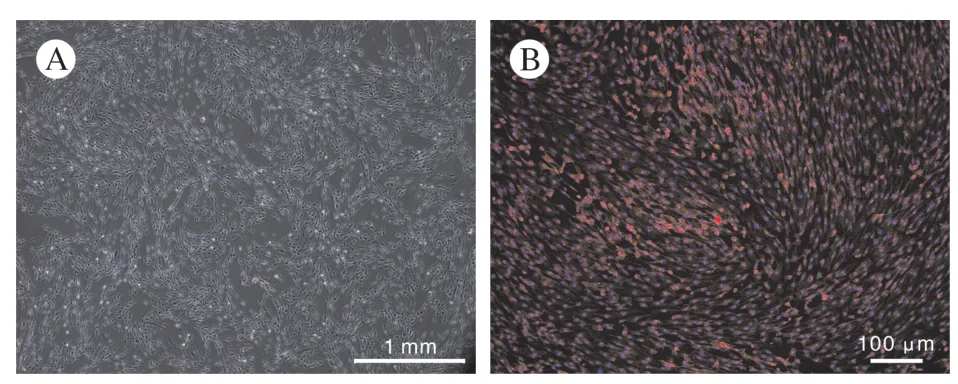
图1 培养成功的施万细胞(A)相差显微镜(40 ×)和S100β 免疫荧光染色(B)Fig.1 Phase contrast micrograph of purified P2 Schwann cells (A)and immunofluorescence staining of S100β of Schwann cells (B)
2 ECM 复合材料的表征 经ECM 修饰后的电纺材料在扫描电镜下可见原始的纳米纤维周围包绕ECM 蛋白,这与未经修饰的纺丝材料初始形貌有很大差异(图2)。经免疫荧光染色验证了这些基质中Fibronectin 的存在:电纺材料在种植SCs 并完成刺激程序后,可见一致取向性排列,并且被ECM 包绕(图3CD)。对复合材料进行去细胞处理后,细胞内容被脱去,且未见完整深染的核结构,但保留一定的细胞外基质成分(图3AB)。在脱细胞处理的效率方面,与未做去细胞处理[对照组=(3 022±51.13) ng/mg]相比,去细胞处理后复合材料的DNA 残余含量[(2.398±0.232 9) ng/mg]显著降低(图3E)。
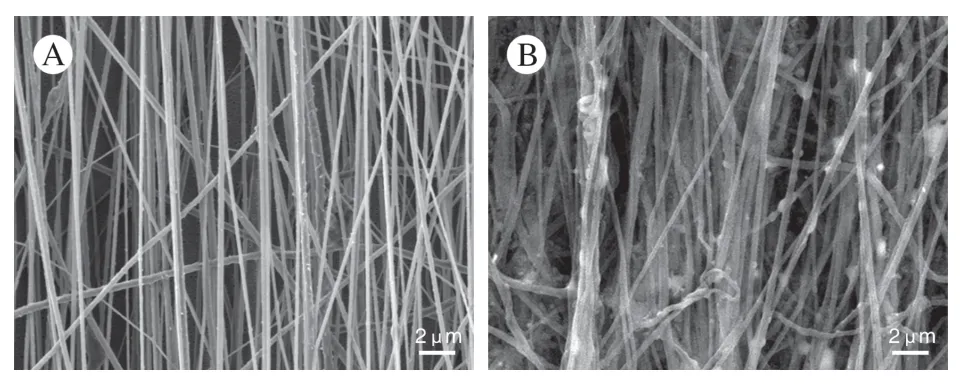
图2 电纺材料扫描电子显微镜照片 A:原始聚己内酯-丝素蛋白取向电纺纤维;B:施万细胞细胞外基质修饰的纳米纤维Fig.2 Scanning electron micrographs A:Initial polycaprolactonesilk protein aligned electrospun fibers;B:Extracellular matrixmodified nanofibers of Schwann cells

图3 去细胞前(A、B)、后(C、D)组复合电纺纤维免疫荧光染色及DNA 含量测定(E)(标尺=100 µm)Fig.3 Immunofluorescence staining of composite electrospun fibers in decellularized group (A,B) and before decellularization(control) group (C,D),and determination of DNA content (E)
3 复合材料对背根神经节轴突延伸以及施万细胞迁移的影响 与单纯的PCL-SF 电纺材料相比,SCs-ECM 修饰的PCL-SF 材料可显著支持轴突的取向性再生和快速延伸(图5B);SCs-ECM 修饰的PCL-SF 材料上DRG 组织中施万细胞的迁移距离显著大于轴突的生长距离[(2 073±112.9) µmvs(1 503±147.4) µm,P=0.005 6](图4B)。

图4 施万细胞外基质修饰的电纺纤维上生长的背根神经节免疫荧光染色(A)和复合材料上施万细胞迁移距离与轴突生长长度比较(B)[施万细胞:S100β(红色);轴突:NF200(绿色);细胞核:DAPI(蓝色)]Fig.4 Immunofluorescence staining of dorsal root ganglia grown on electrospun fibers modified by the extracellular matrix of Schwann cells (A) and comparison of Schwann cells migration distances and axon growth lengths on composites (B)(Schwann cells:S100β [red];axon:NF200 [green];nucleus:DAPI [blue])
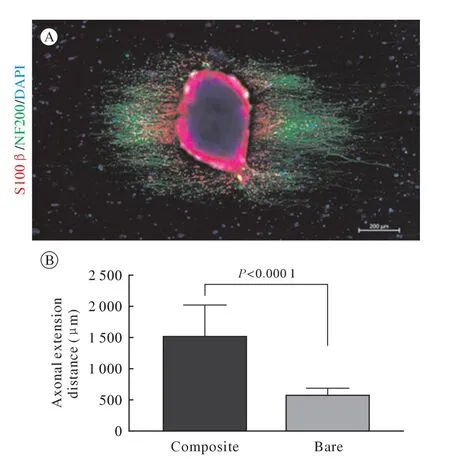
图5 单纯电纺纤维上生长的背根神经节免疫荧光染色(A)和两种材料上轴突纤维生长长度比较(B)[施万细胞:S100β(红色);轴突:NF200(绿色);细胞核:DAPI(蓝色)]Fig.5 Immunofluorescence staining of dorsal root ganglia grown on simple electrospun fibers (A) and comparison of axonal fiber growth lengths on the two materials (B)(Schwann cells:S100β[red];axon:NF200 [green];nucleus:DAPI [blue])
讨论
再生支架材料包括天然聚合物,如壳聚糖、丝素蛋白、海藻酸盐等;人工聚合物如硅胶、聚乙烯醇、聚己内酯等。这些聚合物已经被应用于3D 打印、静电纺丝、凝胶等工艺以模拟神经组织的天然3D 场景[2]。本研究所选用的经典的聚己内酯具有良好的亲和性和可定制性,目前在药物递送和组织工程等应用中进行了试验[6]。然而,在实践中发现PCL 电纺材料没有很强的亲水性能,不利于细胞的黏附和增殖,我们因此引入丝素蛋白与聚己内酯共纺,丝素蛋白提取自蚕丝,因其生物相容性、生物降解性而成为生物医学应用的热门材料[7]。我们通过两者的结合进行静电纺丝制造,聚己内酯为电纺材料提供物理强度支撑和取向性微纳结构,以模拟天然神经基质结构;而丝素蛋白则能增强材料的亲水性能。然而这些产物在置入后仍有可能会因异物反应导致低水平的宿主整合,且它们缺少原生ECM 所具备的生化信息[8]。
与合成材料相比,天然的组织器官或体外培养细胞沉积的ECM 分子显示出增强的宿主反应和整合。然而,在大多数情况下,它们的快速重吸收阻止它们提供长期的结构支撑[9]。为了寻求优化,研究人员将单一的ECM 蛋白[10-12]、短肽段[13-14]、天然ECM(组织器官/细胞来源)[15-16]与聚合材料以物理或化学手段复合,可使两者取长补短,既能维持ECM 的分子耐稳定性,又能赋予材料以可附着位点和再生相关的基质效应。除此之外,异种/同种异体去细胞神经基质能够保留细胞活动的原始位点[17-19]。然而,脱细胞进程机械、化学处理十分容易对复杂基质结构产生破坏。另外,异体组织ECM 来源有限,而且具有生物不相容性和潜在的病原体传播等难题[20]。
不同于组织来源细胞外基质,体外培养的细胞沉积的ECM 具备灵活的组织工程设计方案[21]。通过表面覆盖或混合的方法与器官脱细胞基质、金属、高分子生物材料等组合能得到具有生物活性的再生支架,修饰的支架材料被赋予了多种优秀的生物学属性:细胞外基质分子为再生提供生化环境以及特殊的生物行为启动位点等。已有报道称细胞来源的细胞外基质蛋白修饰的3D 打印支架、静电纺丝等材料可更好地辅助体内骨、肝、心肌等的再生[15,22-23]。在周围神经再生的领域内,Wang 等[24]利用骨髓间充质干细胞分泌的细胞外基质修饰壳聚糖导管,修饰大鼠坐骨神经缺损效果与自体神经移植相似;借鉴于此,我们选择细胞来源细胞外基质修饰传统的静电纺丝材料,探究其支持轴突再生的可行性及优势。
施万细胞是周围神经的胶质细胞,其在生理以及损伤修复状态下均发挥重要作用,其细胞外基质蛋白分子也随不同场景开放不同的功能位点[25]。因此我们选择施万细胞作为细胞外基质的来源。首先,高纯度施万细胞的获取十分重要,在其原代培养过程中,很容易发生成纤维细胞的污染,并且随着细胞的增殖以及传代,施万细胞易发生表性改变以及衰老。通过在培养基中添加抗有丝分裂药物-阿糖胞苷,能够成功抑制成纤维细胞的有丝分裂而不影响施万细胞的活性,完全培养基中的EGF-D 因子可支持施万细胞的快速增殖。在细胞传代后可获得高纯度的施万细胞。扫描电镜和Fibronctin 蛋白的免疫荧光染色证明我们成功将细胞外基质蛋白与取向性的电纺支架结合,而且经典的脱细胞方法未对ECM 蛋白造成很大的破坏。这样,我们成功地在机械和生物效能两个维度模拟神经再生的微环境,力求为神经再生提供最佳辅助。
在体外背根神经节培养模型中,复合材料能够更好地支持轴突的取向性再生和快速延伸,这与未经ECM 修饰的电纺材料形成鲜明对比(图4A、图5A)。轴突的取向再生能够提高再生效率,这在周围神经损伤的恢复中意义重大。同时,我们还发现在复合材料组中,施万细胞的迁移速度大于轴突生长。这是因为施万细胞自身的基质蛋白为自身提供活动位点。施万细胞迁移速度的提高也有利于神经再生进程[26]。
综上所述,本研究成功构建了以施万细胞细胞外基质修饰的聚己内酯-丝素静电纺丝材料,并发现该材料可以显著提高轴突生长速度和施万细胞迁移速度。总之,本研究提供了一种针对经典再生材料的仿生化修饰策略,为周围神经再生材料的设计提供了新见解。
致谢:感谢解放军总医院骨科研究所所有工作人员的技术指导和帮助。
利益冲突声明:作者声明本文无任何利益冲突。
