青海省格尔木牦牛的母系遗传多样性及支系组成
2022-05-18白诗睿马志杰李广祯杨振菲陈生梅谢银禄郭东风
白诗睿, 马志杰*,李广祯,杨振菲,陈生梅,谢银禄,郭东风,陈 立
(1.青海大学畜牧兽医科学院,青海省高原家畜遗传资源保护与创新利用重点实验室,西宁 810016;2.青海省格尔木市畜牧兽医站,青海 格尔木 816000;3.青海省格尔木市唐古拉山镇兽医站,青海 格尔木 816099)
牦牛是青藏高原特有、古老而原始的牛种,享有“高原之舟”的美誉,其能在高寒牧区负重驰骋,是当地牧民不可或缺的生活和生产资料[1]。青海省位于青藏高原的东北部和我国的西北部,平均海拔3 000 m以上,现有牦牛500多万头,是牦牛存栏数最多的省份,被誉为“世界牦牛之都”。格尔木市位于青海省中西部的海西蒙古族藏族自治州,地处青藏高原腹地,辖区内拥有丰富的自然资源,其丰富的动植物遗传资源对三江源区生态保护和维持起着重要作用。畜牧业是格尔木市的主要产业之一,而牦牛业在该地区畜牧业发展中有着举足轻重的作用。线粒体DNA(mitochondrial DNA,mtDNA)具有结构简单、遵循母系遗传和突变率高等特点,被广泛运用于哺乳动物母系遗传多样性、群体结构及系统发育等研究[2]。近年来,基于线粒体基因组序列变异对牦牛遗传多样性的研究相对较多,且mtDNA D-loop区序列的运用最为广泛[3-4]。赖松家等[5]首次测定了我国5个牦牛品种35个个体的mtDNA D-loop区全序列,分析表明其总的单倍型多样度为0.970±0.018且有2个支系,提示这5个牦牛品种母系遗传多样性丰富且有2个母系起源。郭松长等[6]分析了我国10个家牦牛品种296个个体的mtDNA D-loop区部分序列,表明我国家牦牛具有丰富的母系遗传多样性且品种间存在显著的遗传多样性差异,其中环湖牦牛拥有最高的单倍型多样度(0.985±0.040),而巴州牦牛单倍型多样度最低(0.800±0.083);聚类分析显示,家牦牛存在2个聚类簇,推测有2个母系起源。Z. F. Wang等[7]分析了405头家牦牛和47头野牦牛的D-loop区序列,结果显示家、野牦牛总的单倍型多样度分别为0.916±0.009和0.967±0.013;系统发育分析表明牦牛存在3个支系,其中家、野牦牛共享Ⅰ和Ⅱ支系,第Ⅲ支系仅有少量野牦牛分布。钟金城等[8]测定了西藏11个牦牛群体的mtDNA D-loop区、Cytb基因和COⅢ基因的核苷酸序列,其中D-loop区计算的单倍型多样度介于0.927~1.000之间,有2个聚类簇,表明西藏牦牛具有较丰富的遗传多样性且有2个母系起源。成述儒等[9]基于D-loop序列探究了甘肃省6个牦牛群体的遗传多样性,表明甘南州的3个牦牛群体(即玛曲、碌曲和夏河)的遗传多样性较丰富,单倍型多样度分别为0.911±0.021,0.902±0.033和0.883±0.024,而天祝白牦牛群体单倍型多样度最低(0.662±0.031);系统发育分析显示6个牦牛群体有2个支系,可能具有2个母系起源。官久强等[10]分析了四川4个牦牛品种(即昌台、金川、木里和麦洼牦牛)的215条mtDNA D-loop区序列,并与西藏、青海、新疆等地的品种(群体)及野牦牛进行系统发育关系分析,表明昌台牦牛在四川4个群体中具有最高的单倍型多样度(0.883±0.020),木里牦牛具有最低的单倍型多样度(0.621±0.109);四川牦牛与其他地区牦牛品种(群体)拥有明显的2个支系。X. P. Yue等[11]对我国13个家、野牦牛品种(群体)的742条D-loop序列进行分析,表明家牦牛中大通牦牛具有最高的单倍型多样度(0.945±0.025),而环湖牦牛的单倍型多样度最低(0.785±0.048)。涂世英等[12]测定分析了15头中甸牦牛mtDNAD-loop区全序列,其平均单倍型多样度和核苷酸多样度分别为0.983和0.005,表明中甸牦牛的遗传多样性较为丰富。李瑞哲等[13]分析了34头雪多牦牛的mtDNA D-loop序列,其单倍型多样度为0.900±0.043且有2个聚类支系,表明雪多牦牛具有丰富的母系遗传多样性,推测其有2个母系起源。李广祯等[14]测定分析了34头曲麻莱牦牛(即玉树牦牛)的mtDNA D-loop区部分序列,其单倍型多样度为0.952±0.021且有2个明显的分支,提示曲麻莱牦牛具有丰富的母系遗传多样性且有2个母系起源。随后,他还对31头门源白牦牛和235头天祝白牦牛的mtDNA D-loop序列进行了比较分析[15],结果表明门源白牦牛单倍型多样度为0.862±0.033,而天祝白牦牛单倍型多样度为0.946±0.007,提示天祝白牦牛的遗传多样性水平高于门源白牦牛;系统发育分析显示56种单倍型可分为2个明显的分支(即支系Ⅰ和Ⅱ),表明白牦牛有2个母系起源。侯孟典等[16]对64头玉树牦牛和4个西藏牦牛品种(群体)D-loop区进行比较分析,表明玉树牦牛的单倍型多样度最低(0.885±0.034),而申扎牦牛的单倍型多样度最高(0.956);5个牦牛品种(群体)拥有2个母系分支。最近,X. D. Wang等[17]测定分析了青海和新疆5个牦牛品种(群体)的111条全线粒体基因组序列,表明雪多牦牛(0.992±0.015)、帕米尔牦牛(0.990±0.014)、玉树牦牛(0.963±0.033)、祁连牦牛(0.948±0.036)和环湖牦牛(0.905±0.048)的单倍型多样度均较高,具有丰富的母系遗传多样性;系统发育分析显示牦牛可聚为3个分支,而少量雪多牦牛和野牦牛个体共享第Ⅲ母系支系。与此同时,Z. J. Ma等[18]基于84条mtDNA D-loop区序列和24条全线粒体基因组序列的综合分析,系统探究了野牦牛的母系遗传多样性和支系组成,表明野牦牛具有丰富的母系遗传多样性,其中D-loop序列和全基因组序列获得的群体单倍型多样度分别为0.962±0.008和0.993±0.014;系统发育分析显示野牦牛拥有3个母系支系,提示其有3个母系起源。
综上所述,研究者已基于mtDNA序列变异对我国多个牦牛品种(群体)进行了母系遗传多样性和系统发育分析[5-18]。而在先前牦牛的父系遗传研究中,Z. J. Ma等[19]对9个青海省牦牛品种(群体)父系遗传多样性、群体结构及起源的分析表明,格尔木牦牛群体(即郭勒木德牦牛群体)具有最高的Y染色体单倍型多样度(0.706±0.038),提示其具有丰富的父系遗传多样性。然而,当前尚未见对青海省格尔木牦牛群体母系遗传方面的研究报道。鉴于此,本研究基于mtDNA D-loop区序列变异对格尔木牦牛进行母系遗传多样性、群体遗传结构和系统发育分析,以明确其母系遗传多样性水平、群体结构组成和母系遗传背景,为其遗传资源的分子评估、合理保护和开发利用奠定基础。
1 材料与方法
1.1 样品采集与基因组DNA提取
在青海省海西蒙古族藏族自治州格尔木市郭勒木德镇共采集49头牦牛的血液样品,ACD抗凝,带回实验室-20 ℃冷冻保存备用。采样前仔细询问牧户有关牦牛群的选育背景信息并查阅系谱记录,采用分户随机采样方式,确保样品间无亲缘关系且具有代表性。用血液基因组DNA提取试剂盒(北京艾德莱生物科技有限公司)提取基因组DNA,操作步骤严格按照说明书进行,所得基因组DNA -20 ℃保存备用。
1.2 引物合成、PCR扩增与测序
参照Z. J. Ma等[18]在野牦牛研究中运用的引物信息合成一对引物用以扩增格尔木牦牛的mtDNA D-loop区序列,引物由上海生工生物工程有限公司合成,其中上游引物为5’-GTAAAGAGCCTCACCAGTAT-3’,下游引物为5’-TCCTCTAGCCATTGACTAT-3’。
PCR反应体系(25 μL)为:2xPreMix 12.5 μL,超纯水9.5 μL,上、下游引物各1 μL,基因组DNA 1 μL。PCR反应程序为:95 ℃预变性5 min;95 ℃变性30 s,62 ℃退火30 s,72 ℃延伸30 s,31个循环;72 ℃延伸5 min,后4 ℃低温保存。PCR产物用1%琼脂糖凝胶电泳检测后,送纯化PCR产物到北京擎科生物科技有限公司进行测序。
1.3 数据分析
将测序结果核对后用BioEdit 7.2.5软件进行多序列比对,后用DnaSP 5.10.01软件确定格尔木牦牛群体的单倍型数目及其核苷酸序列多态位点,再用Arlequin 3.5.2.2软件计算单倍型多样度以及核苷酸多样度大小以揭示格尔木牦牛的群体遗传多样性水平。使用MEGA 5.05软件基于邻接法(Neighbor-Joining,即NJ法)构建系统发育树以揭示不同单倍型(组)间的系统发育关系,进而揭示格尔木牦牛的母系遗传背景。
2 结果与分析
2.1 格尔木牦牛mtDNA D-loop区序列变异分析
通过测定49头格尔木牦牛mtDNA D-loop区序列,得到有效分析序列片段长度为618 bp,其中T、C、A、G 4种核苷酸的含量分别为27.79%,25.15%,30.85%和16.21%,A+T含量为58.64%,G+C含量为41.36%。多序列比对分析排除1处插入(缺失)位点后,共检测到45处多态位点,约占所分析序列长度的7.28%;在45处多态位点中,共有10处单一多态位点,分别为4,173,209,244,253,282,419,420,562和575位点,占分析序列长度的1.62%,其余35处(即21,35,42,60,67,83,98,113,119,146,172,174,175,181,182,192,199,201,218,221,234,235,236,240,242,250,251,259,281,316,317,323,366,426和464位点)为简约信息位点,占分析序列长度的5.66%(图1)。研究共确定了18种单倍型,其中单倍型H4由10个个体共享,为优势单倍型。单倍型H1由9个个体共享,单倍型H2、H3、H6均由4个个体共享,H9由3个个体共享,H5、H10、H16分别由2个个体共享,其余单倍型均由1个个体拥有。

注:“.”表示与H1单倍型序列中的核苷酸相同;N表示共享同一单倍型的个体数量;Hg表示单倍型组类型。图1 格尔木牦牛mtDNA D-loop区18种单倍型核苷酸变异、频率大小及单倍型组类型
2.2 格尔木牦牛的母系遗传多样性分析
对49头格尔木牦牛mtDNA D-loop区部分序列进行多序列比对分析,计算结果表明格尔木牦牛群体的单倍型多样度为0.911±0.022,核苷酸多样度为0.015±0.008。与此同时,将格尔木牦牛与已报道的野牦牛群体及17个我国其他家牦牛品种遗传多样性指数值进行比较分析(表1),结果表明格尔木牦牛群体的遗传多样性指数值较高,表明其具有较丰富的母系遗传多样性。
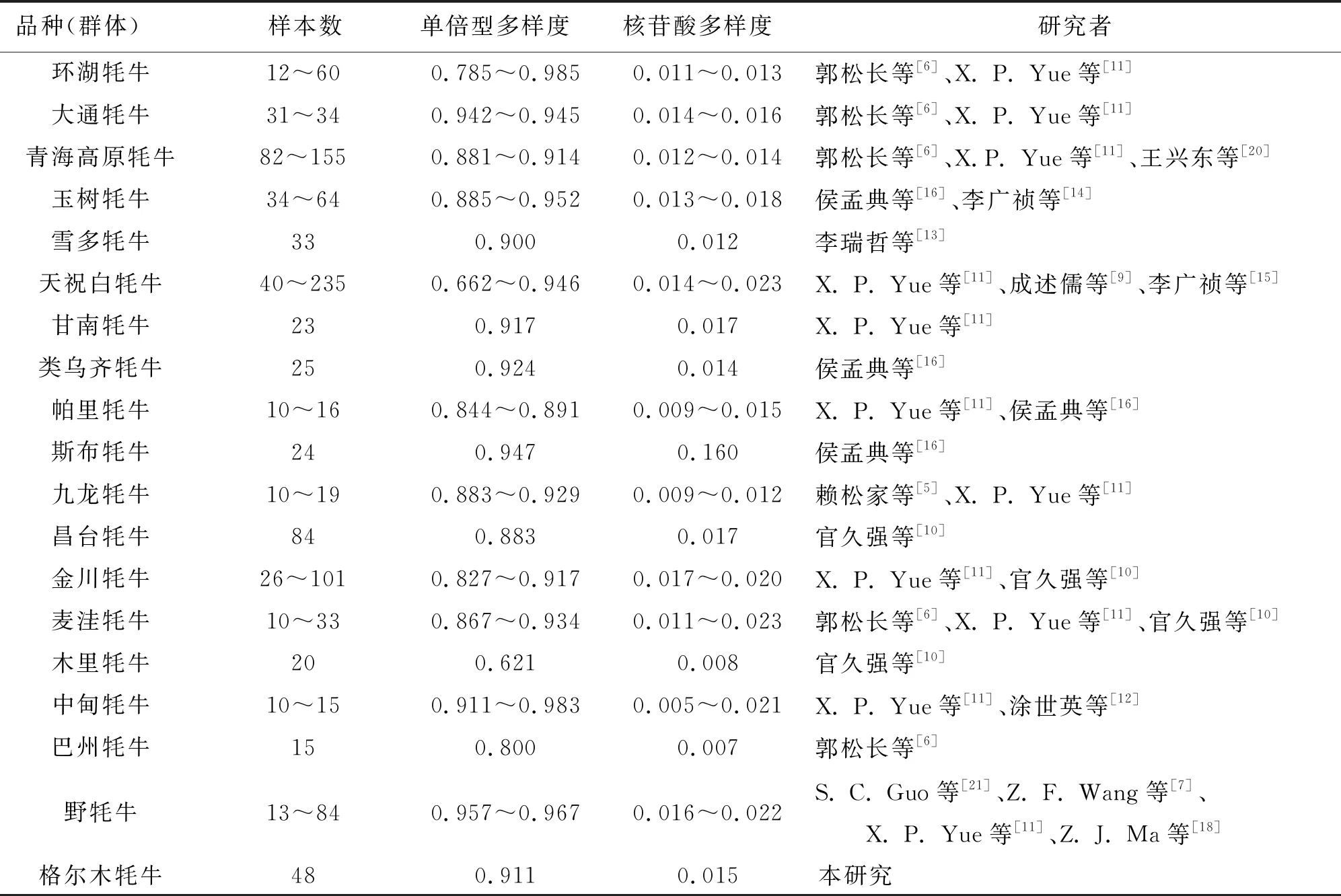
表1 格尔木牦牛与野牦牛及我国其他17个家牦牛品种母系遗传多样性指数值比较
2.3 格尔木牦牛的系统发育分析
将美洲野牛(Bisonbison)mtDNA D-loop序列(GenBank No:EU177871)作为外群,以先前研究报道中代表牦牛3个母系支系的D-loop区序列(即支系I(GenBank No.:FJ548844)、支系II(GenBank No.:FJ548843)和支系III(GenBank No.:DQ139202))作为参考序列[18],对格尔木牦牛18种单倍型序列构建系统发育树。结果显示:格尔木牦牛的18种单倍型序列明显的和3条参考序列分别聚在一起,形成3个母系分支(图2),其中支系Ⅰ由H4、H6、H9、H15、H2、H1、H11、H8、H16、H17、H18、H10和H12单倍型序列组成,占72.2%;支系Ⅱ由H5、H3、H7和H13单倍型序列组成,占22.2%;而支系Ⅲ只包括H14单倍型序列,占5.6%。从单倍型组构成来看,18种单倍型分别隶属于A、B、C、D和G 5种单倍型组,分别占50%,22%,17%,5.5%和5.5%,其中A单倍型组包括H1、H2、H4、H6、H8、H9、H11、H15和H16共9种单倍型,B单倍型组包括H10、H12、H17和H18单倍型,C单倍型组包括H3、H13和H17单倍型,而D和G单倍型组分别包括H5和H14单倍型。
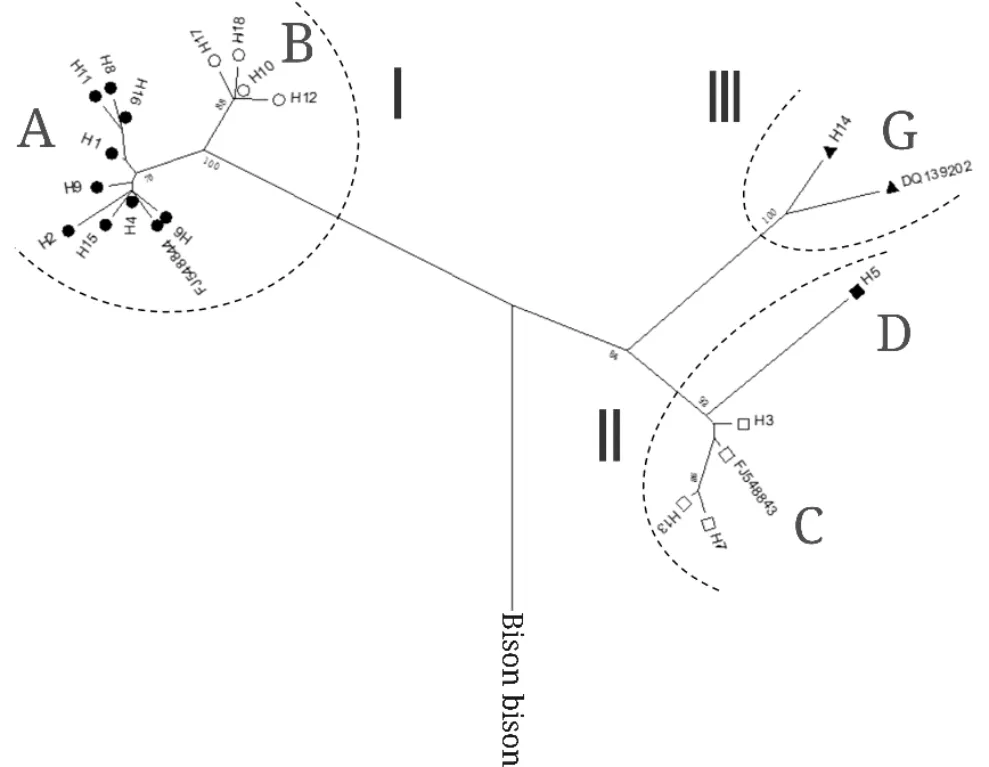
图2 格尔木牦牛母系单倍型(组)间系统发育关系及支系组成
3 讨 论
遗传多样性是生物多样性的重要组成部分,家畜的遗传多样性丰富程度与人类的生存发展休戚相关。牦牛遗传多样性的研究不仅有利于牦牛遗传资源的保护和开发利用,更有助于维持青藏高原生态系统的平衡,具有深远的经济、社会与生态意义。先前的研究表明,野牦牛及我国多数家牦牛品种都具有丰富的母系遗传多样性,其中野牦牛群体的单倍型多样度在0.957~0.967之间[7,18],而家牦牛品种的单倍型多样度在0.621~0.985之间[5-16,20]。在本研究中,格尔木牦牛群体的单倍型多样度为0.911±0.022,核苷酸多样度为0.015±0.008。与我国其他牦牛品种(群体)如野牦牛及天祝、甘南、九龙、类乌齐、帕里、斯布、昌台、金川、麦洼、木里、中甸、巴州、青海高原、环湖、大通、雪多和玉树牦牛相比[5-18,20],格尔木牦牛的单倍型多样度和核苷酸多样度值均较高,表明格尔木牦牛具有较丰富的母系遗传多样性。
牦牛的起源和系统发育研究可以明确其进化历史和遗传背景,对其遗传资源保护和开发利用具有重要作用。先前多数针对我国家、野牦牛品种(群体)的系统发育研究显示:牦牛拥有2个母系支系,推测可能有2个母系起源[5-6,8-11,13-16]。然而,近年来部分研究者如Z. F. Wang等[7]、X. D. Wang等[17]和Z. J. Ma等[18]基于线粒体基因组及其D-loop区、蛋白编码序列等对我国家、野牦牛的系统发育综合分析表明,牦牛拥有3个母系支系,其中共享第Ⅰ、Ⅱ支系的家、野牦牛个体较多,而第Ⅲ支系中仅有少量野牦牛和雪多牦牛个体的存在,推测牦牛有3个母系起源。值得注意的是,在本研究系统发育分析中,首次发现格尔木牦牛的18种单倍型聚为3个母系分支,其中分支Ⅰ、Ⅱ和Ⅲ所占比例分别为72.2%,22.2%和5.6%,表明格尔木牦牛由3个母系支系组成,可能有3个母系起源,进一步支持了牦牛由3大母系支系组成且家、野牦牛共享这3个母系支系的结论[17-18]。牦牛的3大母系分支推测大约在42万年和58万年前分化[22],这与青藏高原更新世中晚期的冰期事件发生的时间吻合,青藏高原的冰期事件促使野牦牛隔离而产生异域分化,冰期结束后重新聚集形成了高度分化的野牦牛基因库,人类在这一基因库中选取了部分个体进行了驯化。考虑到本研究在格尔木牦牛群体中首次检测到第Ⅲ母系支系个体的存在,结合X. D. Wang等[17]在雪多牦牛中也发现第Ⅲ母系支系个体的研究结果,故认为牦牛中稀少的第Ⅲ支系个体早期也应该被驯化了,只是由于个体数量较少,目前只有限的分布在雪多牦牛、格尔木牦牛这些家牦牛群体中,这在一定程度上改变了先前Z. F. Wang等[7]研究中认为第Ⅲ母系支系只由未驯化的野牦牛个体组成的观点。鉴于此,本研究建议加强格尔木牦牛群体遗传资源的保护和利用,充分发掘其遗传潜力,进而开展其保种和选育实践工作。此外,本研究中格尔木牦牛的18种单倍型分布在A、B、C、D和G 5种单倍型组中,所占比例依次为50%,22%,17%,5.5%和5.5%,说明A单倍型组是其优势单倍型组,这与野牦牛及其他家牦牛品种(群体)的研究结果[5-22]基本一致。
