土壤干旱胁迫对巨龙竹实生苗光合特性和抗氧化酶活性的影响
2022-05-07赵秀婷朱书红刘蔚漪辉朝茂
赵秀婷 朱书红 刘蔚漪 辉朝茂
(西南林业大学林学院 竹藤科学研究院/丛生竹工程技术研究中心 昆明650224)
随着全球气候变暖,干旱的频率、程度和范围也越来越大,水分是植物生长发育的重要条件。土壤水分缺乏或者空气中相对湿度较低会引起植物一系列生长发育和生理生化的变化,从而使植物适应干旱环境[1]。植物在干旱胁迫下关闭气孔,导致光合色素合成降低,影响光合电子传递过程,使光合酶活性降低等,进而导致植物光合速率降低[2-4]。王秀文[5]对芦竹(Arundo donax) 的抗旱性研究发现,芦竹能通对调节自身的抗氧化系统来适应干旱环境。研究植物光合和抗氧化酶活性对干旱胁迫的响应,明确植物的抗旱能力,对植物培育、引种具有重要意义。
云南省近年降水逐渐减少,且降水分布不均,这都对植物能否适应逆境生长提出挑战,而不同的竹种光合作用不同,抗旱能力也不同。孔波等[6]研究认为,不同竹种其抗旱能力不同。吴志庄等[7]研究发现,不同竹种的光合固碳能力不同,丛生竹要优于混生竹和散生竹。刘思奇[8]对美丽箬竹(Indocalamus decorus) 的光合特性和抗氧化系统研究发现,在干旱胁迫下美丽箬竹可以通过调节自身对逆境的适应,从而具有一定的抗旱能力。巨龙竹(Dendrocalamus sinicus) 属禾本科竹亚科,热性巨型大型丛生竹,是世界上最大的竹种,分布在北回归线以南地区[9]。巨龙竹以本身特大型为特色,堪称“竹中极品、世界之最”。以巨龙竹整竹用于居民竹楼、郊野别墅、旅游度假区等特色建筑已引起国内外商家关注,具有巨大的市场潜力[10]。巨龙竹的研究起步较晚,扩大竹种资源首先要提高培育种苗的技术水平和苗木质量,种苗已成为制约巨龙竹发展的瓶颈,严重影响了该物种产业链的发展[11]。目前对于巨龙竹的研究主要集中在组织培养、育苗技术、林分结构、竹材结构等[12-16],而对于土壤干旱胁迫下巨龙竹实生苗光合作用和抗氧化酶活性的研究较少。本研究选取2 年生巨龙竹实生苗,采用盆栽控水试验,在土壤自然干旱过程中连续观测巨龙竹实生苗的光合作用,探讨巨龙竹实生苗叶片光合作用和抗氧化酶活性的变化过程,可为了解巨龙竹资源在应对干旱环境下生长发育状况提供技术支撑,也可为巨龙竹实生苗的科学培育、合理栽培和水分管理提供理论支持。
1 材料与方法
1.1 材料
试验材料为盆栽巨龙竹苗,挑选无病虫害且长势较好,株型尽量一致的20 盆实生苗。对照组与试验组各10 盆。
1.2 试验方法
试验时间为2021 年10 月2—20 日,试验地点为西南林业大学温室大棚。水分调节时间于每日下午17 ∶00—18 ∶00,试验组为自然干旱,对照组每日浇水让土壤水分含量始终保持在43.2%~49.3%。利用Li-6400 便携式光合作用测定系统在每日9 ∶30—11 ∶30 测定实生苗的各项光合指标。
试验组在自然干旱条件下,根据土壤含水量将干旱程度划分为轻度干旱、中度干旱和重度干旱3 个胁迫水平(表1),研究不同水分条件下巨龙竹实生苗的光合参数和抗氧化酶活性。
1.3 指标测定
光合指标:净光合速率 (Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci)。叶绿素提取用80%的丙酮溶液[17]。
抗氧化酶活性和丙二醛含量测定:将上述用于测定光合参数后的叶片液氮速冻-80 ℃保存,用于抗氧化酶活性的测定。超氧化物歧化酶(SOD) 活性采用氮蓝四唑法测定[18],抗坏血酸过氧化物酶(APX) 活性采用紫外吸收法测定,过氧化氢酶(CAT) 活性采用过氧化氢分解法测定,丙二醛 (MDA) 含量采取紫外线吸收法测定[19]。
1.4 数据处理
采用Microsoft Excel 2010 进行数据整理,利用SPSS 25.0 进行方差分析。
2 结果与分析
2.1 干旱胁迫对土壤含水量和实生苗叶绿素含量的影响
从图1 看出,自然干旱组的土壤随干旱天数增加其含水量呈现直线下降,对照组土壤含水量由于每天补充水分而在整个试验期内保持在43.2%~49.3%,为正常含水量。
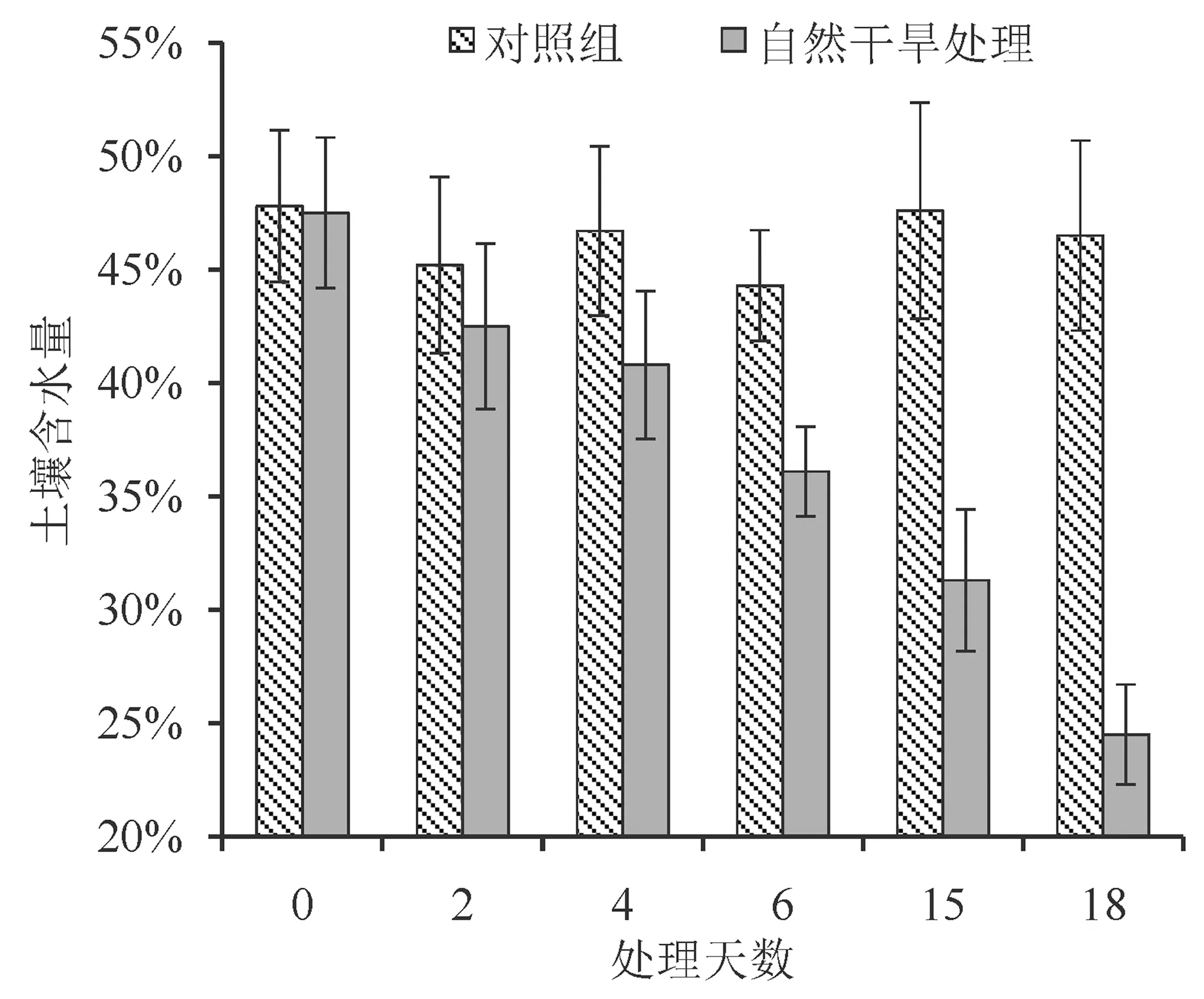
图1 不同干旱胁迫处理下土壤含水量的变化Fig.1 Changes of soil water content under different drought stress treatments
叶绿素是植物体现光合能力的主要指标之一,其浓度(SPAD 值) 与植株的生长发育状况以及叶片的光合能力有关。从图2 可看出,巨龙竹实生苗在干旱胁迫下,叶绿素浓度发生显著改变。随着土壤含水量的下降,干旱胁迫强度增加,SPAD 值呈现明显下降趋势,在轻度干旱胁迫下(0~4 d) SPAD 值下降幅度较小,在中度和重度干旱胁迫下(6~18 d) SPAD 值随着干旱胁迫强度增加而降低幅度加大,中度干旱胁迫下SPAD值降至CK 的67.35%、重度干旱胁迫下仅有CK的43.58%。
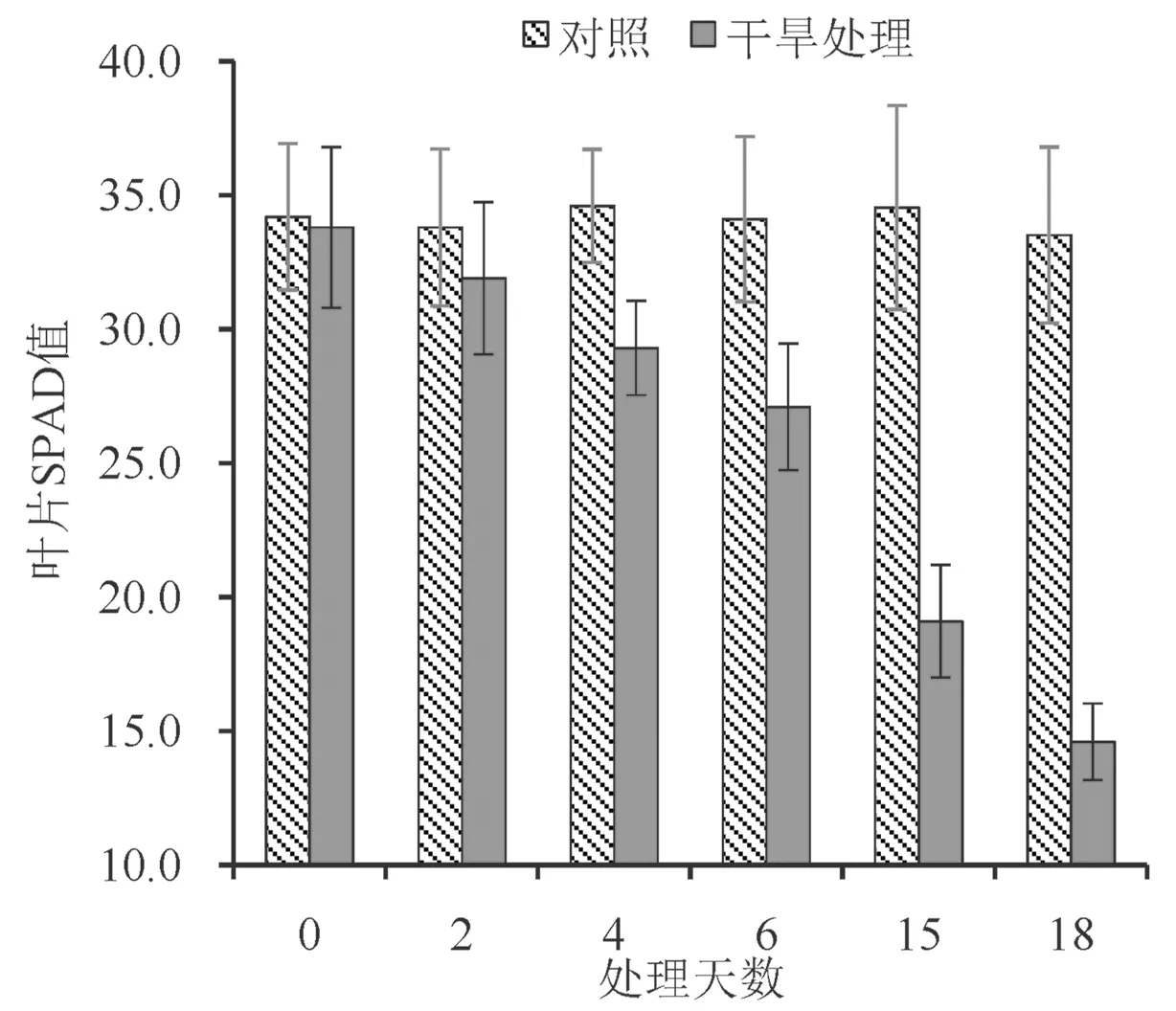
图2 不同干旱胁迫处理下叶绿素含量的变化Fig.2 Changes of chlorophyll content under different drought stress treatments
2.2 干旱胁迫对巨龙竹实生苗光合参数的影响
不同程度的土壤干旱对巨龙竹实生苗光合作用影响的分析结果见图3。巨龙竹实生苗在自然干旱胁迫处理后净光合速率、气孔导度、蒸腾速率变化趋势一致,随着胁迫程度的加重,3 个指标都有不同幅度的下降,胞间CO2浓度呈现先下降再上升的变化趋势。
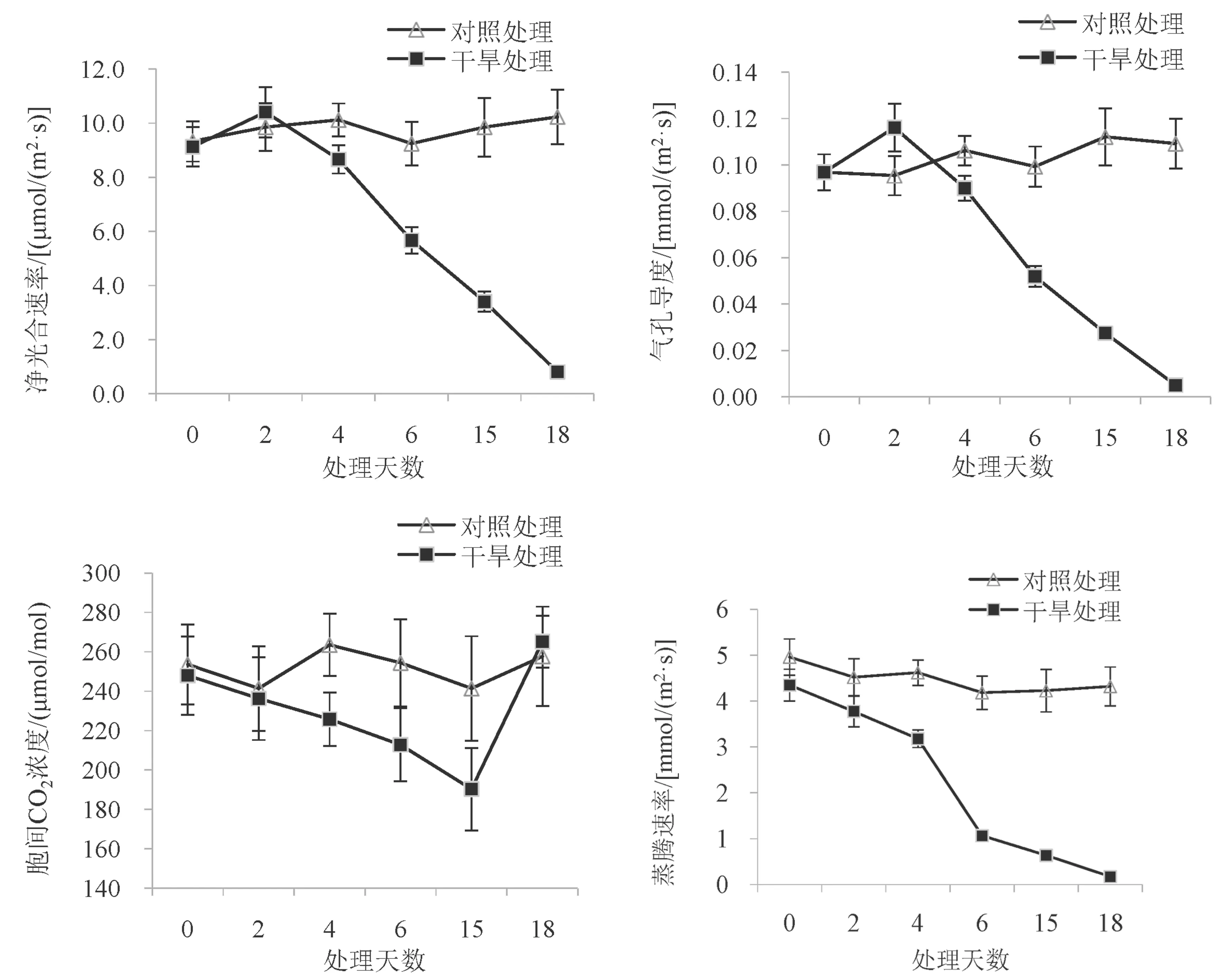
图3 不同干旱胁迫处理下巨龙竹实生苗叶片光合参数的变化Fig.3 Changes of photosynthetic parameters in D. sinicus seedling leaves under different drought stress treatments
净光合速率(Pn):在整个试验期间,自然干旱处理组净光合速率在第2 d 时略有升高,但从第2 d 以后开始下降,在第6 d 时下降趋势加大,最终在第18 d 时跌至最低为0.80 μmol/(m2·s),中度干旱下降至CK 的47.51%、重度干旱下仅为CK 的7.82%。
气孔导度(Gs):Gs变化趋势与净光合速率相同,在第2 d 由升高后开始下降,第6 d 急剧下降,最终在第18 d 时跌至最低为0.005 mmol/(m2·s),中度干旱下降至CK 的37.51%、重度干旱降至仅为CK 的4.58%。并且在重度干旱下,随胁迫程度的加大Gs下降趋势减弱,中度干旱时Gs降低了0.062 mmol/ (m2·s) 与重度干旱时降低了0.022 mmol/ (m2·s) 相比,重度干旱的下降幅度显著低于中度干旱。
胞间CO2浓度(Ci):自然干旱处理组在0~15 d期间呈持续下降,但在16~18 d 期间不断上升至265.17 μmol/mol。在中度干旱下,Ci降至CK 的81.31%。在重度干旱下,自然干旱处理组Ci开始随干旱强度的加大而增加,在第18 d 时与CK 组基本持平。
蒸腾速率(Tr):自然干旱处理组的蒸腾速率在0~18 d 期间均呈下降趋势,并在第4 d 后急剧下降,最终在第18 d 时跌至最低为0.17g/ (m2·h)。说明轻度干旱对巨龙竹实生苗的Tr影响较小,轻度干旱期间Tr降至CK 的76.03%;但在中度干旱和重度干旱胁迫下,Tr受到显著抑制,且胁迫强度越大,蒸腾速率越低,中度干旱和重度干旱胁迫时Tr分别降为对照的20.31%和3.94%。
2.3 干旱胁迫对巨龙竹实生苗抗氧化酶活性和丙二醛的影响
如表2 所示,巨龙竹实生苗处于轻度干旱时超氧化物歧化酶(SOD) 活性高于对照组,且在此胁迫下SOD 活性最高,在中度干旱胁迫下,SOD 活性下降;当干旱加剧到重度胁迫时,SOD的活性又发生显著的提高,巨龙竹实生苗再次通过提高SOD 活性来保护光合作用组织免受或减轻干旱带来的损伤。
抗坏血酸过氧化物酶(APX) 是通过参与抗坏血酸的氧化还原代谢,配合抗坏血酸清除多种活性氧自由基,植物的抗旱能力也通过APX 活性表现出来[20]。从表2 中看出,随干旱强度的加大APX 活性的变化呈上升趋势,在整个胁迫阶段APX 活性均高于对照,说明APX 是巨龙竹在发生干旱时起主要作用的酶,并且在严重干旱时达到最高值为4.32 U/g,为对照组的553%。
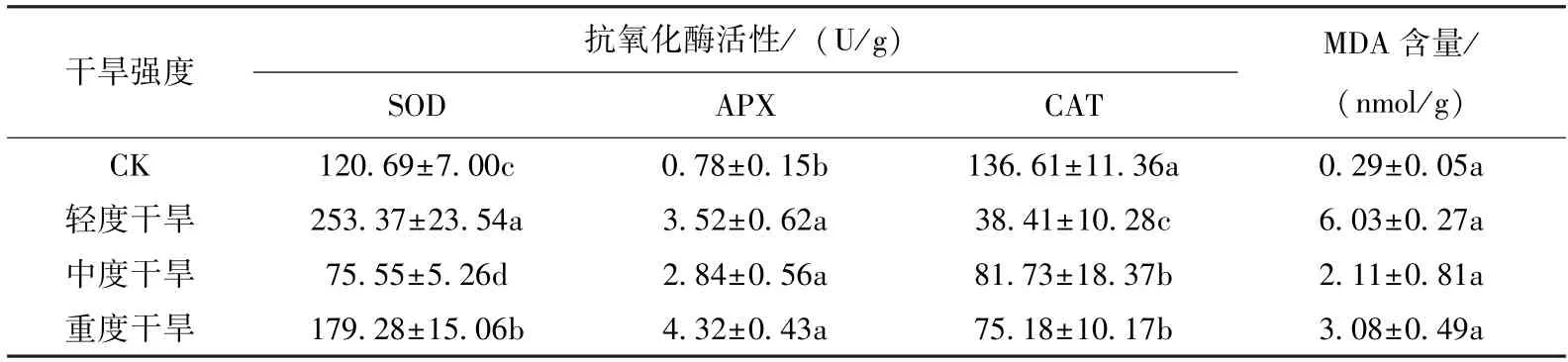
表2 不同干旱胁迫处理下巨龙竹实生苗SOD、APX、CAT 活性与MDA 含量的变化Tab.2 Changes of SOD,APX,CAT activities and MDA content of D. sinicus seedlings under different drought stress treatments
过氧化氢酶(CAT) 将SOD 歧化生成的过氧化氢转化为水和分子氧[20]。从表2 中可以看出,在干旱环境下CAT 活性均小于对照,在轻度干旱环境下达到最低值,在中度干旱时为干旱胁迫下最高值,但仅为对照的55%。
丙二醛(MDA) 是反映巨龙竹干旱能力的重要指标。当干旱发生时,巨龙竹的细胞膜受到伤害,引起膜脂过氧化的最终产物丙二醛含量变化来反映巨龙竹实生苗的抗旱能力。如表2 所示,在干旱胁迫下MDA 含量均高于对照组,且在轻度干旱时最高,随着干旱胁迫强度的加大MDA 逐渐降低,但即使在严重干旱时,MDA 的含量也仅是对照的106%。
3 讨论
3.1 土壤干旱胁迫对叶绿素的影响
植物进行光合作用的一个重要影响因素就是叶绿素含量,叶绿素的合成与分解速率是影响光合速率的主要原因,因此植物受干旱胁迫的影响可以通过叶绿素含量来进行评估[21]。有研究表明,干旱胁迫会对叶绿素的合成起抑制作用,同时加快SPAD 的分解,造成SPAD 值下降,但也有学者发现干旱胁迫会使SPAD 值上升[22-23]。本研究结果表明,在干旱胁迫下,巨龙竹实生苗叶片的SPAD 值均下降,加大干旱胁迫强度SPAD值的下降速度变快。这可能是因为干旱致使蛋白质合成受阻或叶绿体结构被破坏,造成叶绿素的合成减少,使其SPAD 值下降。
3.2 土壤干旱胁迫对光合作用的影响
光合作用是植物生命活动中的重要合成代谢过程之一,会直接影响植株的生长、发育状况,而干旱胁迫明显影响植物的光合作用[24]。大量研究表明,在干旱胁迫下,植物光合作用受到抑制,净光合速率下降,气孔导度减小以限制蒸腾速率,从而缓解干旱胁迫的影响[25-27]。本试验结果显示,在自然干旱处理后,随着干旱胁迫强度加大巨龙竹实生苗叶片的净光合速率、气孔导度、胞间CO2浓度、蒸腾速率均呈下降趋势,表明干旱胁迫下巨龙竹实生苗的部分气孔关闭,引起CO2浓度降低,从而造成光合参数降低;而在重度干旱下,由于巨龙竹自身代谢产生CO2,造成Ci的上升,也说明在轻度干旱下,巨龙竹叶片Pn的下降,Ci降低,是由气孔限制原因引起的;在中度和重度干旱胁迫下,随着胁迫强度的加大,Pn下降,Ci上升,此时引起净光合速率下降转为由非气孔因素引起。这表明随着土壤水分含量下降,干旱加剧,巨龙竹实生苗通过降低光合作用来减轻干旱带来的伤害,本研究结果与王彬等[28]的试验结果一致。
3.3 土壤干旱胁迫对抗氧化酶活性和丙二醛的影响
在干旱环境下,植物启动自身防护系统保护膜不被损伤,通过提高抗氧化酶活性来提高自身抗旱能力,SOD 起第1 道防护作用,APX 和CAT继续将SOD 歧化的过氧化氢分解为水和氧气,从而在干旱环境下起到保护作用。本研究结果显示,SOD 活性在轻度、重度干旱胁迫下高于对照,APX 活性在整个干旱阶段均高于对照,CAT 活性则均低于对照。说明SOD 和APX 是巨龙竹实生苗干旱时期起重要作用的2 种酶,并且在轻度干旱胁迫下SOD 活性的增幅要显著高于其余2 种,说明SOD 在逆境中的反应更为迅速。丙二醛在干旱胁迫下的含量均高于对照组,表明干旱引起巨龙竹抗氧化系统活性下降,活性氧的产生超出了细胞的清除能力,导致活性氧大量积累,活性氧引起光合色素严重降解和膜脂过氧化,破坏了光合机构膜系统,该结果与裴斌等[29]的研究结果一致。
4 结论
巨龙竹实生苗在轻度干旱环境中表现出良好的适应性,但在中度和重度干旱环境下其适应性较差。引起光合速率下降的原因也由在轻度干旱时的气孔限制原因转为在中度和重度干旱时的非气孔限制原因,并且在逆境中起主要作用的抗氧化酶为SOD 和APX,SOD 在逆境中反应更为迅速。本研究结果可对巨龙竹苗的科学种植和水分管理提供理论依据,也可为竹类植物对干旱环境下的适应性研究提供技术支持。

