斑翅果蝇YolkProtein1基因原核表达及抗体制备
2022-04-27祝晴王瑞娟覃冬云陈浩代晓彦刘艳周紫章翟一凡
祝晴 王瑞娟 覃冬云 陈浩 代晓彦 刘艳 周紫章 翟一凡



摘要:斑翅果蝇(Drosophilasuzukii)是一种农业害虫,在世界范围内分布广泛,可对多种水果产生不可逆损害,造成巨大经济损失。本试验对斑翅果蝇YolkProtein1(Ds-YP1)基因进行原核表达,并对Ds-YP1蛋白的功能结构域和信号肽进行预测,克隆多肽并构建pET-32a-c(+)-Ds-YP1原核表达载体,IPTG诱导表达并免疫小鼠制备抗体;用成功制备的抗体进行斑翅果蝇蛋白WesternBlot验证。结果表明构建的pET-32a-c(+)-Ds-YP1表达载体在大肠杆菌中表达了His-Ds-YP1融合蛋白,通过免疫小鼠成功制备并纯化了Ds-YP1多克隆抗体。制备的Ds-YP1多克隆抗体不但能识别His-Ds-YP1融合蛋白,而且能识别斑翅果蝇Ds-YP1蛋白,具有较好的特异性。本研究成功制备了斑翅果蝇Ds-YP1的多克隆抗体,对进一步研究YolkProtein蛋白作用机制具有重要意义。
关键词:斑翅果蝇;卵黄蛋白;原核表达;抗体
斑翅果蝇(Drosophilasuzukii)是一种入侵性、破坏性作物害虫,最早在日本发现[1],雄性在每个翅膀上均有一个黑点[2,3]。斑翅果蝇不像其他大多数果蝇物种只攻击腐烂的水果[4,5],其更喜欢未受损的成熟水果[4,5];利用锯齿状产卵器可刺穿果实(如樱桃和一些浆果)相对坚硬的表皮并在其中产卵,在2008年首次侵入美国时就对其造成了约50亿美元的损失[6-8]。
卵黄蛋白(yolkprotein)是一种对生殖极为重要的糖脂蛋白,是所有卵生脊椎动物和无脊椎动物卵黄的主要成分[9,10]。其前体在雌性脂肪体内合成,释放到循环系统中,并通过受体介导的内吞作用运输到生长中的卵母细胞中,或由卵巢滤泡细胞直接合成作为营养源被利用;经20-羟基蜕皮酮调控后也可在少数雄性体内合成,但量比雌虫低数千倍[11,12]。大多数果蝇体内存在至少两种YP基因,有些果蝇例如黑腹果蝇(D.melanogaster)存在3种[13]。在表达方面YP1表达含量往往更高,其次是YP2,其在卵巢与脂肪体中的表达也并不一致[14]。
有关果蝇的生殖滞育已有报道,但关于斑翅果蝇的滞育却鲜有人研究[15]。本实验室前期已对其滞育温度、光周期与滞育率进行研究,在此基础上继续对其滞育机制进行探究[16,17]。本研究通过对斑翅果蝇的YolkProtein1(Ds-YP1)基因进行分析,构建pET-32a-c(+)-Ds-YP1原核表达载体,表达并制备蛋白特异性抗体,以期为卵黄蛋白的作用机制研究提供支持。
1材料与方法
1.1供试昆虫
斑翅果蝇采自山东省泰安市樱桃园,由本实验室人工饲养。饲养条件:温度(25±1)℃,湿度(50±5)%,光周期16L∶8D。成、幼虫皆用人工饲料饲喂,成虫期补给20%糖水。人工饲料配方:麦麸39.1g、蔗糖20.8g、啤酒酵母5.3g、山梨酸0.17g、尼泊金乙酯0.1g、对羟基苯甲酸甲酯0.07g、抗坏血酸0.09g、维生素溶液1mL、水100mL
1.2序列分析
斑翅果蝇YP1基因序列下载自NCBI,序列AccessionNO.为XM_017081211.2。分别用InterProScan5(http://www.ebi.ac.uk/Tools/pfa/iprscan5/)和signalP4.1Server(http://www.cbs.dtu.dk/services/SignalP/)对Ds-YP1的功能结构域和信号肽进行预测。用DNAMAN(version6)进行序列翻译及展示。
1.3引物设计
用VectorNTI软件与NCBI的Primer-BLAST进行引物设计(表1)。由上海生工生物工程技术服务有限公司合成。
1.4RNA提取和cDNA合成
取10只斑翅果蝇置于1.5mL离心管中,放于液氮速冻研磨,按R6934-01TotalRNAKitⅡ(Omega)试剂盒使用说明提取RNA,最后加入20μLNuclease-FreeWater洗脱RNA,-80℃保存。
按TQ2501-02M-MLVFirstStrandcDNASynthesisKit(Omega)试剂盒说明书合成cDNA,合成产物保存于-20℃冰箱备用。
1.5PCR扩增
以斑翅果蝇cDNA为模板进行PCR扩增,扩增体系(50μL):2×phantaMaxBuffer25μL,dNTPMix(10mmol/Leach)1μL,上下游引物Ds-YP1-1F/R或Ds-YP1-2F/R各2μL,cDNA1μL,phantaMaxSuper-FidelityDNAPolymerase1μL,Nuclease-FreeWater補足至50μL。反应程序:94℃预变性5min;94℃变性30s,62℃退火30s,72℃延伸1min,共35个循环;72℃延伸5min。PCR结束后用1%琼脂糖凝胶电泳检测(120V、25min)。
1.6重组表达载体构建
扩增产物经1%琼脂糖凝胶电泳检测后,切取凝胶中的目标条带,用胶回收试剂盒(北京聚合美生物科技有限公司)进行回收。将胶回收产物与pET-32a-c(+)(Novagen)一起用限制性内切酶EcoRⅠ(Thermo)、NotⅠ(Thermo)进行双酶切。将酶切后DNA产物与载体产物胶回收后,用T4连接酶(Takara)进行连接,组成pET-32a-c(+)-Ds-YP1重组表达载体。
将酶连产物转入EscherichiacoliDH5α(天根生化科技有限公司),在含有氨苄青霉素抗性的LB固体培养基上涂板,用无菌玻璃涂布器将菌液轻轻涂布均匀,封口膜封口后,倒置于37℃恒温培养箱培养12~16h。挑取单菌落至LB液体培养基中,37℃过夜培养,进行阳性克隆鉴定,体系为10μL:2×TaqMasterMix5μL,T70.4μL,T7Terminator0.4μL,菌液1μL,ddH2O补足至10μL,反应程序同1.5。
将鉴定正确的菌液送上海生工生物工程技术服务有限公司进行测序,测序结果用VectorNTI软件进行比对。选择测序结果正确的克隆提取质粒(Vazyme,FastPurePlasmidMiniKit),-20℃保存。
1.7蛋白诱导表达
将重组质粒转入E.coliBL21(天根生化科技有限公司)中,在含有氨苄青霉素抗性的LB固体培养基上%板,待长出菌落后挑单菌落于LB液体培养基中,37℃、220r/min培养,菌液OD600值达到0.6左右时加入IPTG诱导剂至终浓度0.1mmol/L,37℃、150r/min培养6h,之后4℃、8000r/min离心10min收集菌体。
在未诱导与诱导6h样品菌体中加1×PBS(0.2g/mL),振荡悬浮后用超声波细胞粉碎机(宁波新芝生物科技有限公司)进行破碎:功率20%,超声每次2s,间隔3s,破碎至菌体均匀清亮。
离心收集上清和沉淀,上清作为蛋白上清样品;沉淀经含8mol/L脲的1×PBS溶解后再離心得到上清液作为包涵体;超声破碎后未经离心的样品作为混合物样品。
将蛋白上清、包涵体和混合物样品在蛋白电泳仪(Bio-Rad)上进行SDS-PAGE电泳,恒压120V电泳至溴酚蓝条带跑出胶板下边缘关闭电泳仪。将蛋白胶于考马斯亮蓝R250染色液中染色30min,过夜脱色后观察蛋白表达情况。
1.8WesternBl
在SDS-PAGE电泳结束后,用湿转电泳仪(Bio-Rad)在冰浴条件下进行转膜,300mA恒流转膜120min。结束后将0.45μmPVDF膜(MilliPore)浸入封闭液中,摇床常温孵育1h,用TBST漂洗10min;加入用BSA稀释10000倍的MouseantiHis-TagmAb(ABclonal)4℃过夜孵育;回收一抗后再用TBST漂洗3遍,每遍10min;加入用BSA稀释10000倍的辣根酶标记山羊抗小鼠IgG(北京中杉金桥生物技术有限公司),常温摇床孵育2h;TBST漂洗4遍,每遍10min;最后使用极超敏化学发光试剂盒(山东思科捷生物技术有限公司)显色,于ChemiScope系列荧光及化学发光成像系统(上海勤翔科学仪器有限公司)中显影。
1.9抗体制备
目的蛋白条带切胶-20℃保存。将蛋白胶条放入干净研钵中,研磨至黏稠状,加0.9%生理盐水稀释备用。SPF级KM小鼠(济南朋悦实验动物繁育有限公司)7只,每只20g左右,喂养3天后用1mL针管45度角注射腹部位,每次每只小鼠注射300~500μL,注意排尽针管空气。每周免疫一次,连续4周,第5周摘眼球取血。
血液放于1.5mL离心管中,立即放入37℃水浴锅1h,然后4℃放置6h,之后4℃、12000r/min离心15min,取上清,液氮速冻后放于-80℃冻存。
1.10果蝇总蛋白提取
提取蛋白全程均在冰上操作,研磨棒需预冷。取15只斑翅果蝇放入液氮速冻,加200μL研磨液进行冰浴研磨,然后冰上静置30min,每隔几分钟混匀一次;离心机预冷至4℃,12000r/min离心5min,取上清;加入蛋白上样缓冲液100℃金属浴5min;-20℃保存备用。
2结果与分析
2.1序列分析及克隆
Ds-YP1序列开放阅读框为1320bp,编码蛋白共439个氨基酸。Ds-YP1蛋白具有信号肽(1~19aa)和完整的vitellgenin功能结构域(129~410aa,图1),去除信号肽后经软件预测分析,分子量为46.96kD。对完整的开放阅读框进行PCR克隆及测序,结果表明,序列长度与数据库序列一致,且无突变碱基(图2)。
2.2重组表达载体的构建
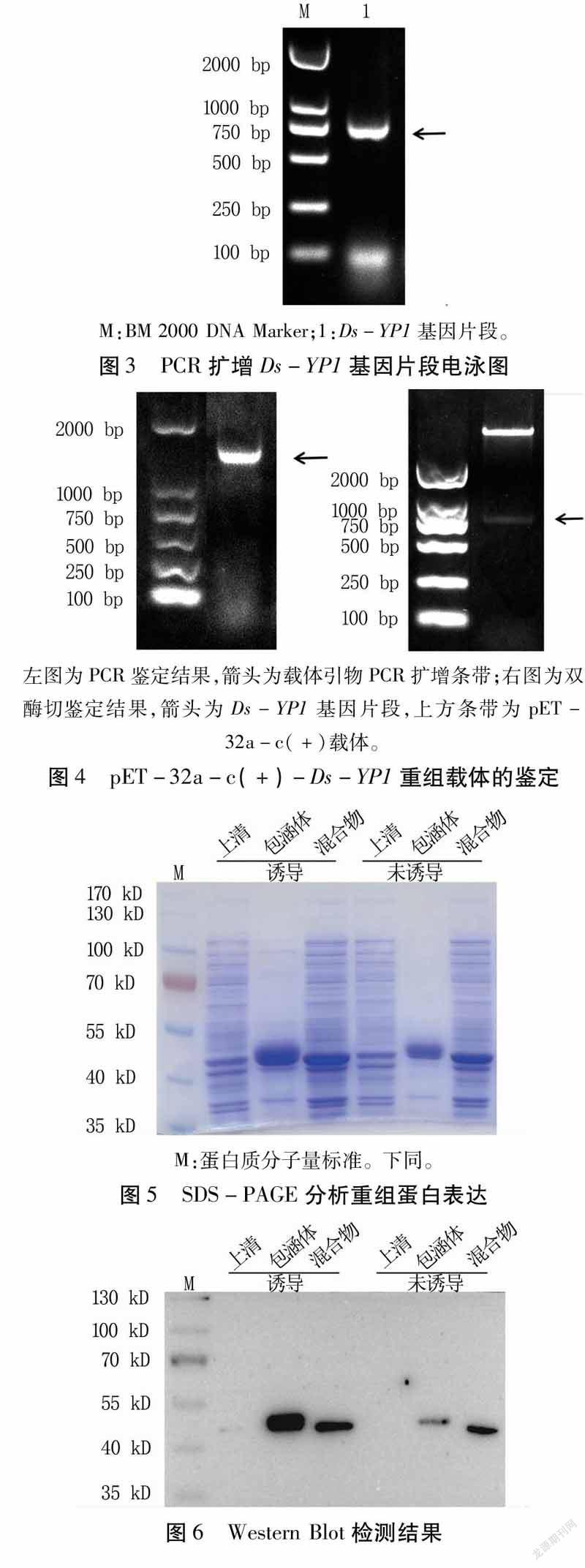
首先选取不含信号肽的部分序列进行扩增(图3),将扩增目的片段与pET-32a-c(+)载体经EcoRⅠ、NotⅠ酶切,经连接转化后构成重组表达载体pET-32a-c(+)-Ds-YP1。构建的载体用通用引物经PCR和双酶切鉴定,得到的片段大小与预期一致(图4)。测序结果发现重组表达载体中的插入片段与Ds-YP1基因的序列完全一致,且无移码现象。以上结果说明,pET-32a-c(+)-Ds-YP1重组表达载体构建成功,可以用来诱导表达重组蛋白。
2.3重组蛋白在大肠杆菌中诱导表达
将未诱导与诱导样品的蛋白上清、包涵体和混合物样品分别进行SDS-PAGE电泳,考马斯亮蓝R250染色后,发现目的蛋白在蛋白上清、包涵体及混合物中都有表达,且包涵体中表达量最高(图5)。从WesternBlot检测结果也可看出,目的蛋白与His抗体发生特异结合且与预期大小一致,证明重组蛋白正确表达(图6)。
2.4抗体制备与检测
将包涵体中的重组蛋白经SDS-PAGE电泳后切胶回收免疫KM小鼠,4周后收集血清,检测血清中的抗体Anti-Ds-YP1。将血清稀释5000倍后,检测未诱导与诱导样品,结果显示,条带单一且效价高(图7)。随后用制备的Anti-Ds-YP1抗体WesternBlot检测斑翅果蝇蛋白样品,结果显示,在抗体稀释18000倍时,条带清晰,信号强(图8)。
以上结果表明制备的Anti-Ds-YP1特异性强且效价高,结果与预期相符,可用于后续试验。
3讨论与结论
卵黄蛋白是卵黄发生时沉积在卵内供胚胎发育营养所需的卵内贮藏蛋白。昆虫的卵黄蛋白主要分为卵黄蛋白原(vitellogenin)、卵黄多肽(yolkpolypeptides)和小卵黄蛋白(minorYP)三种[18]。其中卵黄蛋白原是一类大分子量的糖脂复合蛋白,与非昆虫的卵黄蛋白原进化上同源,可能起源于共同的祖先[19]。卵黄多肽存在于高等双翅目昆虫中,功能等同于其它昆虫卵黄蛋白原,但是分子量较小,约为45kD[20]。小卵黄蛋白是在鳞翅目昆虫卵内分布的一类特异性蛋白[21]。本研究中,斑翅果蝇的卵黄蛋白大小为46.96kD,属于卵黄多肽。
虽然卵黄多肽功能上与卵黄蛋白原相似,但其进化上却完全不同,卵黄蛋白原与脊椎动物的血清载脂蛋白B(apolipoproteinB)具有较高的同源性,卵黄多肽与肝酯酶(hepaticlipases)和胰酯酶(pancreaticlipases)进化上同源。在黑腹果蝇中有3种YP,主要在脂肪体和卵巢泡细胞中合成,但在不同组织中的分布和表达不同[22]。果蝇(Drosophilagrimshawi)中也有3种YP,在脂肪体中YP1合成最多,而YP2和YP3较少;在卵巢内,YP2合成最多,YP1较少,而YP3几乎不存在[23]。本试验用抗体检测斑翅果蝇YP(图8),发现在目的条带上方存在条带,这可能是斑翅果蝇的其它YP,其不同YP的含量不同可能与果蝇的发育时期和组织分布有关。
本试验通过构建重组质粒,转化大肠杆菌后进行原核表达,并用小鼠进行抗原免疫,成功获得了高质量抗体。后续研究将利用该抗体对卵黄蛋白在斑翅果蝇中的功能进行分析。本研究结果将有助于对斑翅果蝇生殖发育等机制开展更深入的研究,为果蝇防治工作提供新的途径。
