非编码RNA 调控口腔鳞状细胞癌葡萄糖代谢重编程的研究进展
2022-04-19杨一言高继萍续国强王晓堂宋国华
杨一言高继萍续国强王晓堂宋国华
(山西医科大学实验动物中心,实验动物与人类疾病动物模型山西省重点实验室,太原 030001)
口腔鳞状细胞癌(oral squamous cell carcinoma,OSCC)是口腔癌中最常见的实体肿瘤,占所有病例的95%以上[1-2],结合肿瘤的临床T 分期、临床N 分期、淋巴结外扩展等多个因素,患者术后复发率高,且5 年生存率无明显提升[3]。 此外,接受治疗后仍会面临肿瘤复发,预后差等问题,严重影响了患者的生活状态与心理健康。 因此,为进一步完善临床治疗方案,我们需要阐明OSCC 的分子调控机制。
葡萄糖代谢重编程是一种广泛存在肿瘤细胞中的生物学现象,即有氧条件下肿瘤细胞通过有氧糖酵解优先利用葡萄糖这一独特代谢表型。 越来越多的证据表明,ncRNA 可以与癌基因或抑癌基因相互作用,调节肿瘤细胞的能量代谢[4-7]。 因此,本文将对部分ncRNA 影响调控OSCC 有氧糖酵解相关的机制进行综述,发掘葡萄糖代谢在临床治疗中的潜在价值。
1 肿瘤葡萄糖代谢重编程
代谢重编程是肿瘤的10 大特征之一,其中最经典、研究最多的是有氧糖酵解(aerobic glycolysis),其特点为即使在氧气足够支持OXPHOS 的情况下,癌细胞仍倾向于将葡萄糖“发酵”成乳酸,这一现象被称为Warburg 效应,我们广泛认为这是人类癌症的中心标志,同时也是促进肿瘤细胞增殖转移的重要过程。 这种代谢转换通常是原癌基因(如Myc)、转录因子(如缺氧诱导因子-1,HIF-1)、信号通路(如PI3K)的激活,以及肿瘤抑制因子(如p53)失活所致。 此外,信号通路、癌基因和抑癌基因等能够直接控制增殖细胞中的代谢途径。 经过上述几种途径作用,激活或过表达葡萄糖转运蛋白(GLUT)和糖酵解通路关键酶,增强肿瘤有氧糖酵解[4]。
肿瘤发生发展过程中,增殖细胞不仅需要大量ATP,还允许糖酵解中间产物(“碳源”)穿梭到几个生物合成途径产生核苷酸、NADPH、脂质和非必需氨基酸,为合成大分子物质提供所需底物或中间体。 与此同时糖酵解还能够生成有助于细胞增殖的小分子前体或中间体,比如乙酰辅酶A、非必需氨基酸的中间体、核糖等物质,以此满足DNA 快速复制的需要[5-6]。 乳酸是糖酵解的产物,当乳酸堆积在细胞外基质并降低其酸碱度时,酸性的环境将促进肿瘤侵袭和转移[7]。 综上所述,Warburg 效应是肿瘤细胞茁壮生长的最佳方式,是癌细胞的一种基本变化。 糖酵解过程需要GLUT、己糖激酶2(HK2)、磷酸果糖激酶2(PFK2)、乳酸脱氢酶A(LDHA)和丙酮酸脱氢酶激酶1(PDK1)等多种酶,将葡萄糖转化为乳酸,与此同时也伴随着PI3K/Akt/mTOR、Wnt/Snail 等信号通路的改变,这些酶、
信号通路的激活或失活会影响肿瘤的葡萄糖代谢。
2 OSCC 中葡萄糖代谢重编程调控作用机制
2.1 葡萄糖转运
GLUT 家族有14 个成员,GLUT1 是GLUT 家族中第一个被鉴定的,其研究最为广泛[8]。 其功能为将葡萄糖从毛细血管转运到细胞中。 正常生理情况下,GLUT 能够快速转运葡萄糖,而恶性肿瘤细胞常通过上调细胞膜GLUT 表达增加葡萄糖摄取量,满足细胞增殖所需。
研究发现,OSCC 组织中GLUT1 的表达显著增加[9],提示GLUT1 可作为OSCC 患者预后的标志。Eckert 等[10]通过对OSCC 样本进行免疫组织化学染色并与临床病理资料比较,结果证明GLUT1 的表达水平能够为肿瘤侵袭和预后提供信息,同时也是筛选OSCC 高危人群的标志物。 对患有OSCC 并发生淋巴结转移的受试者进行免疫组化检测,发现GLUT1 表达显著升高。 同时一项针对舌鳞状细胞癌的研究表明GLUT1 与淋巴结转移相关,说明可以将GLUT1 作为淋巴结转移的指标[11-12]。 非典型GLUT 表达不一定意味着细胞内葡萄糖摄取增加,也可能是受到癌细胞中转录水平的组织特异性基因表达的调控。 最近研究表明,von Hippel-Lindau(VHL)肿瘤抑制基因会下调编码血管内皮生长因子(VEGF)和GLUT1 的mRNA 表达水平。 在检测的OSCC 细胞株中VHL 与GLUT1 的mRNA 表达呈负相关,VHL 的降低可能会增强GLUT1 表达导致葡萄糖摄取增加,在OSCC 的发病机制中起重要作用[13]。
2.2 葡萄糖代谢相关酶
调控糖酵解的主要3 种限速酶,分别是己糖激酶、磷酸果糖激酶、丙酮酸激酶。 3 种酶介导不同过程,在糖代谢中发挥着重要的作用。 除此之外,乳酸脱氢酶通过电子受体NAD 的再生在有氧糖酵解中发挥关键作用。
己糖激酶(HK)是调控糖酵解步骤的第一个限速酶,催化葡萄糖磷酸化。 在头颈部肿瘤中,HK2的表达显著升高[14]。 在缺氧条件下会引起HK2 上调进而增强糖酵解,研究发现低氧环境中HIF-1α 表达显著升高,肿瘤微环境内部处于缺氧状态,HIF-1α 促进HK2 过表达,从而促进舌鳞状细胞癌细胞糖酵解[15]。 Qu 等[16]发现在OSCC 中,去泛素酶USP13 对GLUT1 和HK2 具有调控作用,从而影响糖酵解。
磷酸果糖激酶(PFK)是糖酵解过程中第二个限速酶,有PFK1 和PFK2 两个亚型。 PFK1 能够催化6-磷酸果糖(F-6-P)转化为果糖-1,6-二磷酸(F-1,6-BP)。 果糖-2,6-二磷酸(F-2,6-BP)能够通过与PFK1相互作用,拮抗三磷酸腺苷(ATP)的抑制作用并增加葡萄糖摄取。 果糖-2,6-二磷酸酶(PFKFB3)作为PFK1 的变构活化剂,是糖酵解的速率调控酶,能够维持高糖酵解率并在多种人类肿瘤中高度表达。 已有研究表明,PFKFB3 可以作为阻断糖酵解的靶点,阻止肿瘤细胞的迁移和侵袭[17]。 恶性肿瘤中,PFK2 通过活化Ras、c-Src 等原癌基因提升其表达,并且诱导丙酮酸激酶(PK)转化为糖酵解酶复合物,促进糖酵解[9]。
丙酮酸激酶(PK)是糖酵解通路中另一个限速酶,能够催化磷酸烯醇式丙酮酸(PEP)去磷酸化为丙酮酸并产生ATP。 PKM1 和PKM2 是其中两种异构体,癌变组织中PKM2 表达升高并在能量代谢中居于主导地位。 有研究表明,PKM2 能够通过增加其他蛋白的磷酸化、稳定性和表达来促进肿瘤的侵袭和迁移[18]。 OSCC 中PKM2 通过ETS1 基因依赖方式诱导癌细胞中基质金属蛋白酶9 的表达,从而增加细胞的侵袭能力[19]。 此外PKM2 能够通过诱导上皮细胞向间充质细胞转化(epithelial to mesenchymal transition,EMT)的发生促进肿瘤的发生发展。 例如OSCC 中细胞质中的PKM2 会被转移到细胞核中,与TGF-β 诱导同源因子2(TGF-βinduced factor homeobox 2,TGIF2)结合,使其泛素化降解,诱导EMT 发生,促进OSCC 的发生发展[20]。另外,有些生物合成途径则是通过降低PK 活性发生,研究发现酪氨酸激酶受体刺激和信号转导以及HIF-1α 的诱导会诱导丙酮酸激酶二聚体M2 同种型(PKM2)表达,由此限制PEP 到丙酮酸的转化,使得上游糖酵解中间体(“葡萄糖碳”)积累,进入其他生物合成途径[4]。
LDHA 是乳酸脱氢酶(LDH)的亚型之一,糖酵解过程中,催化丙酮酸转化为乳酸并产生NAD+。乳酸作为一种重要的致癌物质,能够降低肿瘤微环境酸碱度,进而导致肿瘤生长、免疫逃逸等情况发生。 有研究表明,OSCC 中LDHA mRNA 的表达水平显著上调,沉默LDHA 后细胞的葡萄糖消耗和乳酸产量降低,抑制肿瘤细胞的生长迁移能力[21]。LDH5 是催化效率最高的同工酶,有利于恶性肿瘤发生、迁移和侵袭,研究发现其在癌细胞中与GLUT1、HIF-1α 具有相关性。 在有氧糖酵解过程中,LHD5 还能够提高乳酸产量,通过抑制脯氨酸羟化酶进而维持HIF-1α 激活。 将LDH5 作为治疗靶点,有助于修改癌症治疗中修改HIF-1α 调控的基因,达到下调葡萄糖摄取抑制肿瘤增殖的效果[22]。因此LDH 对肿瘤细胞有氧糖酵解的影响能够为OSCC 临床治疗提供新方向。
2.3 肿瘤免疫微环境
肿瘤微环境(tumor microenvironment,TME)主要由免疫细胞、成纤维细胞、血管和淋巴管网络等组 成。 肿 瘤 免 疫 微 环 境 ( tumor immune microenvironment,TIME)是免疫系统与肿瘤的交锋点,在维持免疫抑制环境中代谢改变起重要作用。研究发现在OSCC 中肿瘤相关巨噬细胞(TAMs)浸润可能与免疫逃逸有关,CD68+和CD163+TAMs 浸润与肿瘤中程序性死亡配体1(PD-L1)高表达显著相关,PD-L1 抑制抗肿瘤免疫,导致T 细胞凋亡[23]。此外GLUT1 缺乏会损害效应T 细胞的增殖和功能,削弱CD8+T 细胞的抗肿瘤效应[24]。 研究发现OSCC 细胞侵袭前高CD8+T 细胞浸润的患者,复发所致死亡可能性较小,因此可以将侵袭前CD8+T 细胞浸润作为预后的生物标志物[25]。 Warburg 效应产生的乳酸堆积会降低TME 的pH,分解细胞基质导致肿瘤细胞迁移。 乳酸作为糖酵解的终产物,在TME 中的积累能够阻断T 细胞内乳酸转运从而破坏T 细胞代谢,抑制T 细胞的增殖和活化[26]。 此外细胞内pH 降低还会诱导NK 细胞凋亡,在结直肠癌肝转移中导致自然杀伤细胞(natural killer cell,NK)耗竭[27]。 目前有关OSCC 中TME 与葡萄糖代谢的研究鲜有报道。
2.4 葡萄糖代谢信号通路
葡萄糖代谢受到复杂信号网络通路的调控。其中PI3K/Akt/mTOR 信号通路在癌症的发生发展中起重要的调控作用。 磷酸肌醇3-磷酸(PI3K)可调节下游效应因子Akt 和哺乳动物雷帕霉素靶标(mTOR),Akt 是驱动肿瘤糖酵解表型的重要因子,一方面可调节GLUT1 的表达和膜转位进而激活糖酵解,另一方面还能调节HK2、PKM2 等糖酵解酶的表达和活性影响糖酵解。 mTOR 能够诱导许多有氧糖酵解和肿瘤生长相关基因转录,如HIF-1α、核转录因子-κB(NF-κB)和c-Myc 等,通过这种方式调节糖酵解。 因此,PI3K/Akt 能够调控有氧糖酵解这一特性使其成为临床治疗的新靶点。
Wnt/Snail 信号通路是另一条调节通路,Lee等[28]研究表明,Wnt/Snail 信号通路能够通过诱导丙酮酸羧化酶促进糖酵解,这一过程依赖于β-连环蛋白/T 细胞因子4/Snail 信号通路。 p53 基因则能够调节糖酵解和OXPHOS 之间的转换影响Warburg效应,一项研究表明p53 在口腔癌中低表达[29],细胞色素c 氧化酶2(SCO2)是p53 的转录靶标,同时也是COX 复合物的关键调节因子[30],在氧化应激情况下经SCO2 介导细胞会在p53 影响下完成从糖酵解到OXPHOS 这一转变,而缺乏功能性p53 的细胞则继续进行糖酵解。
3 非编码RNA 调控OSCC 葡萄糖代谢重编程
非编码RNA 是不能编码蛋白质的RNA 分子,包括小分子RNA(microRNA,miRNA)、长链非编码RNA(long non-coding RAN,lncRNA)以及环状RNA(circular RAN,circRNA)等。 在不同类型的肿瘤细胞中,他们能够通过不同的机制调节肿瘤的葡萄糖代谢。例如乳腺癌细胞中,miR-155 通过miR-155-sOCS1-sTAT2-HK2 和miR-155-C/EBPβ-miR-143-HK2两条途径级联促进癌细胞葡萄糖代谢[31]。 lncRNA XIST通过发挥分子海绵作用吸附miR-126,激活IRSI/PI3K/Akt 通路并以此增强葡萄糖代谢能力,最终促进胶质母细胞瘤侵袭、转移,提高肿瘤抗凋亡能力[32]。 OSCC 中,针对ncRNA 调控葡萄糖代谢重编程进行了研究并获得一些成果(图1)。
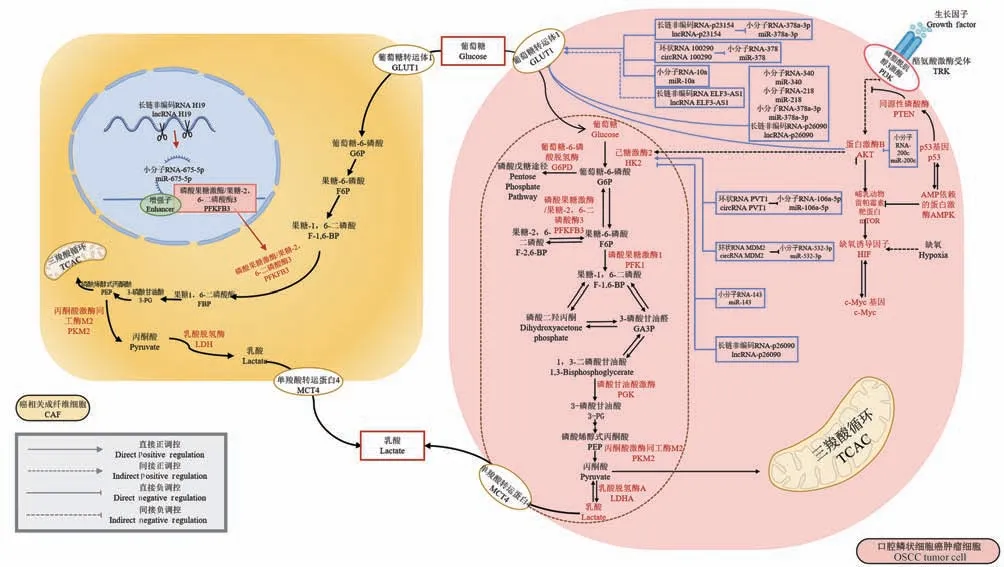
图1 非编码RNA 调控OSCC 肿瘤细胞、成纤维细胞糖酵解示意图Figure 1 Schematic diagram of non coding RNA regulating glycolysis of OSCC tumor cells and fibroblasts
3.1 miRNA 在OSCC 葡萄糖代谢中的调控作用
miRNA 是长度约为22 个核苷酸的非编码单链RNA,在调控基因表达中起关键作用。 近年来,越来越多的miRNAs 被认为是OSCC 细胞代谢的关键内源性调节因子。
例如,miR-340 能够直接与GLUT1 的3’UTR 区域结合,下调GLUT1 表达减弱OSCC 细胞的有氧糖酵解。 体外功能研究显示,miR-340 能够显著降低葡萄糖的摄取率,提示miR-340 所诱导的糖酵解改变为OSCC 细胞的生长提供了必须的能量条件[33]。Xu 等[34]证明miR-218 过表达能够显著抑制OSCC细胞系中GLUT1 的表达,同时也降低了细胞系的葡萄糖摄取,表明miR-218 通过靶向抑制GLUT1 表达,从而削弱糖酵解以抑制OSCC 细胞生长。miRNA 不仅可以抑制糖酵解,也能够促进糖酵解。Chen 等[35]通过qRT-PCR 检测发现,肿瘤组织中miR-10a 和GLUT1 表达水平显著增加,体外细胞实验表明,miR-10a 能够同时促进口腔癌细胞的葡萄糖摄取和增殖。 认为miR-10a 通过作为GLUT1 的上游激活剂,增加GLUT1 的表达,促进OSCC 细胞的葡萄糖代谢和癌细胞增殖,在OSCC 中发挥致癌作用。 HK2 作为糖酵解的第一个限速酶,诸多研究证明其能够调节肿瘤的发生和迁移。 Sun 等[36]研究发现,miR-143 可以直接靶向HK2,抑制其表达,从而抑制细胞葡萄糖代谢,阻止OSCC 癌细胞的生长和侵袭。 miR-532-3p 可靶向HK2,由于竞争性作用,通过circRNA MDM2/miR-532-3p/HK2 轴增加HK2 的表达,促进有氧糖酵解[37]。 除了直接靶向GLUT1、HK2,miRNA 还可以通过信号通路调节下游GLUT1 的表达,影响OSCC 细胞的增殖。 miR-200c能够抑制Akt 通路使下游GLUT1 失活,从而抑制OSCC 细胞对葡萄糖的摄取,减少能量供应抑制增殖[38]。
目前,有关miRNA 调控肿瘤细胞糖酵解的研究越来越多,很多成果也证实了miRNA 对肿瘤细胞Warburg 效应的调控机制。 研究miRNA 在葡萄糖代谢中的调控作用,开发新的治疗靶点将作为miRNA 在OSCC 发生、发展、诊断及治疗的进一步研究思路和方向。
3.2 lncRNA 在OSCC 葡萄糖代谢中的调控作用
lncRNA,是一类长度超过200 个核苷酸的转录本,缺乏编码蛋白质的潜力,是参与转录和转录后调控的支架、阻滞剂、激活剂或海绵。 近期研究发现,OSCC 中存在大量差异表达的lncRNA,这些lncRNA 可以调控多个参与葡萄糖代谢的下游靶点,如GLUT1 和GLUT4、酶(丙酮酸羧化酶、G6P)、癌基因(c-Myc)等,此外也可能与一个或多个信号通路相互作用,参与OSCC 的发生和发展[39-41]。 Wang等[42]发现了lncRNA-p23154-miR-378a-3p/GLUT1轴可调控糖酵解,miR-378a-3P 靶向结合GLUT1 的3’ 非编码区,抑制OSCC 细胞中GLUT1 表达,lncRNA-p23154 与miR-378a-3p 呈负相关,lncRNAp23154 通过与miR-378a-3p 的启动子区域相互作用抑制其转录,使GLUT1 表达增高,改变葡萄糖代谢。lncRNA-p26090 在OSCC 细胞系及癌组织中的表达水平显著上调,同时细胞中糖酵解相关基因GLUT1、HK2、PKM2 等表达显著降低,生成的乳酸减少,提示lncRNA-p26090 可能通过调节相关基因的表达从而调控OSCC 细胞糖酵解过程[43]。 Chu等[44]研究发现在OSCC 细胞中lncRNA ELF3-AS1通过间接调控GLUT1 表达来影响OSCC 细胞对葡萄糖的摄取。 除肿瘤细胞外,lncRNA 也可以调控癌相关成纤维细胞(CAFs)的糖酵解,影响肿瘤进展。CAFs 作为肿瘤微环境的重要组成部分,能够分泌能量代谢产物,为肿瘤进展提供能量。 Yang 等[45]发现lncRNA H19 通过 lncRNA H19/miR-675-5p/PFKFB3 轴促进口腔CAFs 中的葡萄糖代谢,进而促进细胞增殖和迁移。
然而对于lncRNA 在OSCC 糖酵解中的具体作用机制,目前研究相对较少。 因此,需要进一步深入研究lncRNA 调节葡萄糖代谢的途径和机制,推进对癌症代谢复杂调控网络的理解,并为临床诊断和治疗提供更好的理论依据。
3.3 circRNA 在OSCC 葡萄糖代谢中的调控作用
circRNA 呈封闭环状环结构,且高度稳定。 常作为竞争性内源RNA(ceRNA),通过内源性竞争作用“海绵吸附”miRNA,调控下游靶基因的表达水平。
circRNA 可以靶向miRNA 介导Warburg 效应进而调节肿瘤进展。 例如circRNA100290 在OSCC 中充当ceRNA,抑制miR-378 表达,从而促进GLUT1表达,调节OSCC 细胞糖酵解[46]。 此外,circRNA 也可以介导HK2 表达水平调节OSCC 的糖酵解。 Zhu等[47]发现miR-106a-5p 能够抑制HK2 的表达来阻碍OSCC 进程中的糖酵解,circRNA-PVT1 经海绵吸附作用直接吸附miR-106a-5p,进而增加HK2 的表达,促进OSCC 细胞的糖酵解而发挥致癌作用。
有文献表明OSCC 患者唾液中hsa-circ-0001874与has-circ-0001971 的含量明显增高,此类在患者唾液样本中显著增高的circRNA 可以作为诊断OSCC的有力证据[48]。 由于circRNA 的表达具有阶段特异性和稳定性,因此可以作为诊断靶点对患者进行快速无创的早期筛查、诊断。
4 展望
综上所述,有氧糖酵解表型是癌细胞代谢重编程的重要组成部分,一系列错综复杂的分子机制使得肿瘤细胞能够不受控制、持续生长并进行侵袭和转移。 OSCC 发病率日渐增高的当下,现有成果表明ncRNA 可能通过影响葡萄糖代谢的部分环节调控OSCC 发展方向。 目前一个亟待解决的问题是仍需大量研究明确ncRNA 在OSCC 细胞葡萄糖代谢中复杂的调控网络及各级调控机制。 我们需要再进一步探寻更加全面的分子作用网络的基础上,结合其作用位置找寻新的治疗靶点,为临床OSCC 诊断和治疗提供更多研究思路。
