蔗糖对拟南芥根系偏斜角度的影响
2022-03-19贾容容杨亚婷姬新宇张净然田喜元
贾容容,杨亚婷,姬新宇,张净然,田喜元
(河北农业大学,河北保定 071000)
根系是植物生长发育必不可缺的器官,不仅起到固定、支持植物的作用,还能从土壤中吸收植物生长发育必需的水分和养分。植物在生长发育过程中,根系能根据外界环境的变化不断调整其生长方向以保证其对水分和养分的最佳吸收状态[1-2]。研究表明,拟南芥在琼脂培养基上生长时,不同品种拟南芥根系的偏斜程度不同,Columbia 生长有轻微的扭曲,而Landsberg和Wassilewskija有较大的偏斜[3]。此外,琼脂的浓度也会影响拟南芥根系的倾斜幅度[4]。可见拟南芥根系的倾斜受品种、年龄以及外界环境影响。
前期研究发现,琼脂培养基上垂直生长的拟南芥根系有严重偏离重力方向的倾向,且野生型的倾斜总是向左倾斜的,研究表明,这种生长模式是根与培养基表面的机械接触和重力共同的作用结果[1,5-6]。其中,糖类是植物体内重要的碳源和能源,它不仅参与能量代谢,还作为信号分子调控植物的生长发育和生理过程[2]。糖作为植物生长中重要的营养物质,是否会对拟南芥的根系偏斜生长造成影响;根的偏斜程度是否受糖浓度的影响尚不清楚。
另外,外源糖也可以参与对植物中氮同化酶以及离体叶片中硝酸还原酶的表达[7]。而NRT1.1是植物根系吸收硝酸盐后第一个发挥作用的蛋白,除了参与植物硝酸盐转运过程以外,还对植物吸收重金属产生影响。Wang Wei的研究表明,AtNRT1.1可以通过磷酸化调控其硝酸盐转运活性来响应环境中硝酸盐的波动。这一关键发现表明,AtNRT1.1实际上是硝酸盐传感的转环[8]。那么基于外源糖与植物硝酸盐转运之间的关系,在拟南芥中,氮的转运和吸收在外源蔗糖供应下拟南芥的根系偏斜生长中是否存在一定的作用?基于此,本研究以野生型拟南芥Col-0及NRT1.1突变体chl1-5和nrt1.1-1为材料,通过在琼脂培养基中添加不同浓度的蔗糖,研究了蔗糖对拟南芥根系偏斜程度的影响,并对拟南芥中NRT1.1基因在根系偏斜程度中的作用进行了初步的探索。
1 材料与方法
1.1 试验材料
供试植物:拟南芥野生型Col-0和拟南芥NRT1.1突变体nrt1.1-1、chl1-5。
供试营养液:改良后MS营养液(具体成分见表1,pH=5.8)。
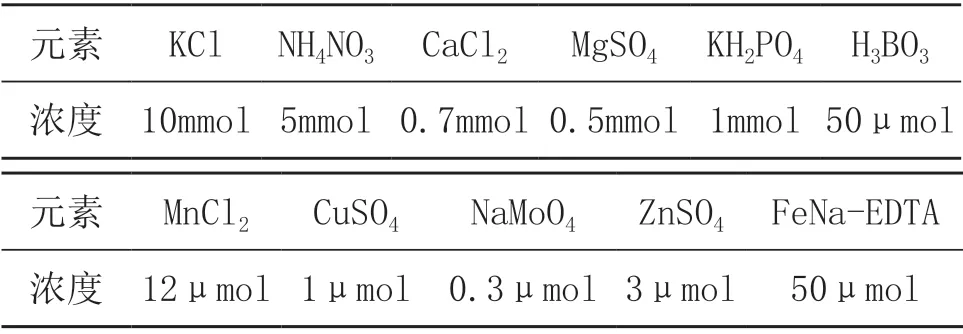
表1 改良后MS营养液成分
1.2 植物培养
培养基配制:培养基使用改良后MS配制,向配制好的改良后MS营养液中添加1%琼脂和1%蔗糖,配制完成后灭菌,再将培养基晾至50℃左右,在超净台中向方形培养皿(10cm×10cm)倒入50ml培养基,在超净台中晾至培养基凝固(约25min)。
种子灭菌:将种子放入1.5ml离心管中,加入70% 酒精,浸泡1min后加入20 %次氯酸钠+0.5%吐温20浸泡10min,使用灭菌水清洗5-6次后,待用。
植物培养:使用枪头将灭菌后种子点至琼脂培养基上,20-25粒/排,3-4排/板。直至种子点完,使用封口膜将培养皿边缘封住。将播种好种子的培养皿放入4℃冰箱中24h后将其转移至培养箱中培养。培养箱条件:光照/黑暗(16h/8h),温度(21℃/20℃)。培养4天后进行处理。转移至不同浓度蔗糖的平板,转移时将根竖直朝下放置,封口膜封口,然后竖立(根朝下)放置至培养箱。培养7天。
1.3 试验设计
培养基使用改良后MS营养液配制,向配制好的改良后MS营养液中添加1%琼脂和相应浓度蔗糖,蔗糖含量分别为0,1%,2%,3%。配制完成后灭菌,灭菌将培养基晾至50℃左右后在超净台中倒入50ml至方形培养皿中,在超净台中晾至培养基凝固(约25min)。将培养4天后拟南芥转移至不同浓度蔗糖的培养皿中,转移时将根竖直放置于培养基上,4株/培养皿。转移结束后,封口膜封口,然后将根竖直朝下放置至培养箱。培养7天后收获。
1.4 测量指标及方法
收获培养7天后的拟南芥幼苗,使用全自动根系扫描仪对培养皿进行扫描,用 ImageJ-Evan 测量幼苗根长和倾斜程度;用剪刀剪下主根根尖部分,I2-KI溶液染色,制片后用显微镜(x100)观察根尖中的淀粉粒,并用显微镜配置的数码照相机拍摄根尖;每2株植株作为一个样品,测量并记录其生物量。
1.5 数据处理
采用Microsoft Office Excel 2010办公软件对试验数据进行整理,利用SPSS15.0对数据进行显著性差异分析。
2 结果与分析
2.1 不同浓度蔗糖对拟南芥根系偏斜方向的影响
拟南芥根系的倾斜是琼脂表面的依赖行为,培养基中的营养物质可以影响到根系的倾斜。研究表明,无论是野生型Col-0还是NRT1.1突变体,各浓度蔗糖培养基上的根系的根尖均存在向左倾斜的现象。由表2可见,无论是野生型还是突变体拟南芥,根系偏斜角度均表现为随蔗糖浓度的增加先增加后降低,并在2%蔗糖处理达到最高,野生型Col-0,突变体chl1-5和nrt1.1-1拟南芥根系偏斜角度分别是不添加蔗糖的2.65倍,1.68倍,1.50倍。但是,当蔗糖浓度达到3%时,野生型拟南芥Col-0根系倾斜角度显著降低,而NRT1.1突变体chl1-5和nrt1.1-1根系倾斜角度与2%蔗糖处理差异不显著。在0蔗糖处理时,野生型Col-0与NRT1.1突变体拟南芥根系偏斜角度有显著性差异,NRT1.1突变体拟南芥根系偏斜角度分别是野生型Col-0拟南芥根系偏斜角度的1.34倍(chl1-5),1.71倍(nrt1.1-1)。在1%,2%蔗糖处理时,野生型拟南芥根系偏斜角度比NRT1.1突变体拟南芥根系偏斜角度大,但差异不显著。在3%蔗糖处理时,野生型Col-0与NRT1.1突变体拟南芥根系偏斜角度存在显著性差异,突变体ch11-5与突变体nrt1.1拟南芥根系偏斜角度分别是野生型Col-0拟南芥根系偏斜角度的1.80倍,2.05倍。这说明,NRT1.1突变体根系偏斜程度受蔗糖浓度影响程度小于野生型。
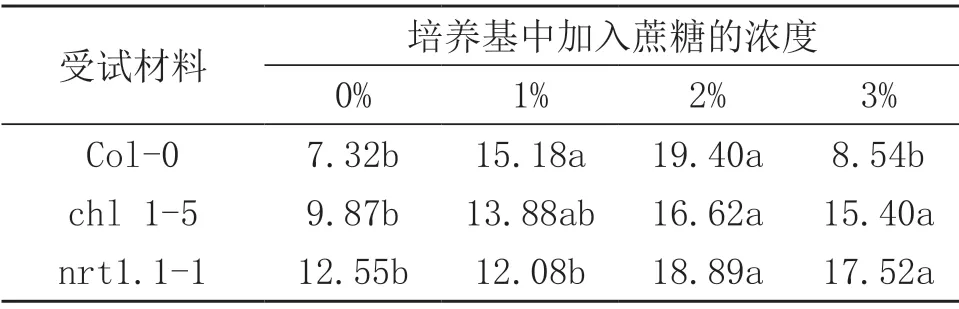
表2 不同浓度蔗糖对拟南芥根系偏斜角度的影响(单位:°)
2.2 不同浓度蔗糖对拟南芥主根长的影响
由表3可知,无论是野生型还是NRT1.1突变体拟南芥,添加蔗糖均会显著提高主根长度。添加3%蔗糖处理,野生型Col-0,突变体chl1-5和nrt1.1-1拟南芥主根长分别是不添加蔗糖的1.52倍,1.58倍和1.50倍。另外,对不同浓度下各拟南芥根长与根倾斜角度进行相关性分析发现,野生型和NRT1.1突变体的根长和根系倾斜角度不存在显著相关关系(P>0.05)。
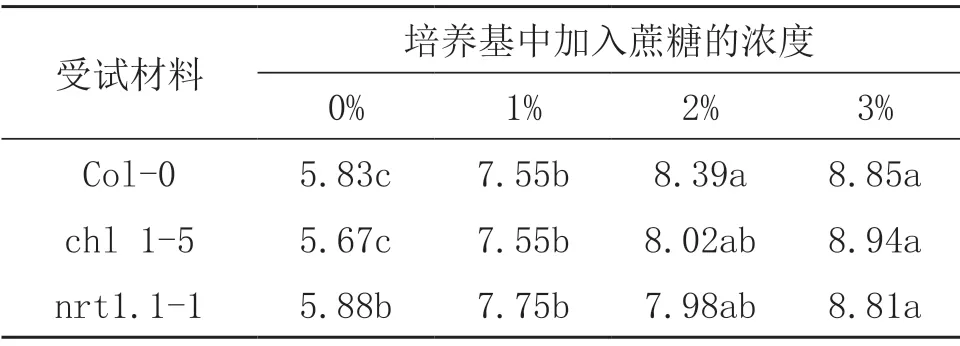
表3 不同蔗糖浓度对拟南芥主根长的影响(单位:cm)
2.3 不同浓度蔗糖对拟南芥生物量的影响
由表4可知,无论野生型还是NRT1.1突变体拟南芥,添加蔗糖均会显著提高拟南芥的生物量。添加3%蔗糖处理,野生型Col-0,突变体chl1-5和nrt1.1-1拟南芥生物量分别是不添加蔗糖的2.54倍,2.10倍和2.40倍。NRT1.1突变体生物量受蔗糖浓度影响程度小于野生型。另外,对不同浓度下各拟南芥生物量与根倾斜角度进行相关性分析发现,野生型和NRT1.1突变体的生物量与根系倾斜角度均不存在显著相关关系。(P>0.05)。
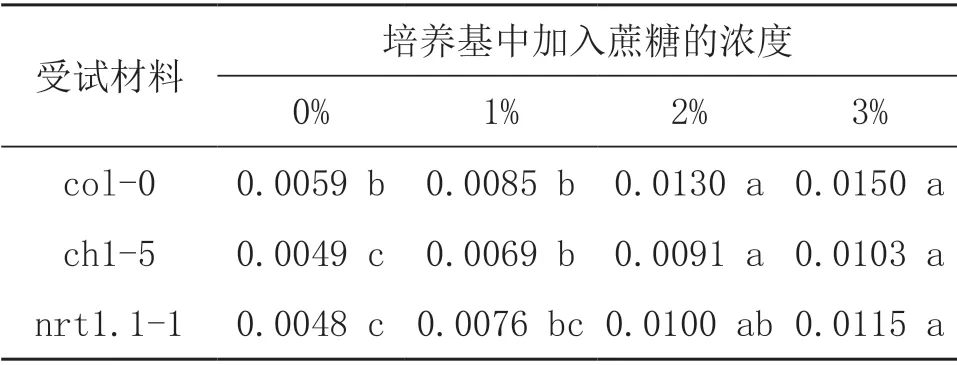
表4 不同蔗糖浓度对拟南芥生物量的影响(单位:g)
2.4 I2-KI溶液染色结果
目前,对于植物感受重力刺激机制多为淀粉-平衡石假说,该假说认为植物当重力方向发生改变时,平衡石细胞内的淀粉能够沿重力方向沉降感受重力刺激[11]。因此,根尖的淀粉粒含量直接影响植物根系对重力信号的感受。
由图1可以看出,NRT1.1突变体与野生型Col-0拟南芥根尖的淀粉粒在0%蔗糖浓度时淀粉含量最少。随着蔗糖浓度含量的增加淀粉粒的颗粒由小逐渐变大,根尖淀粉粒逐渐增多,但野生型与突变体并没有明显差异。
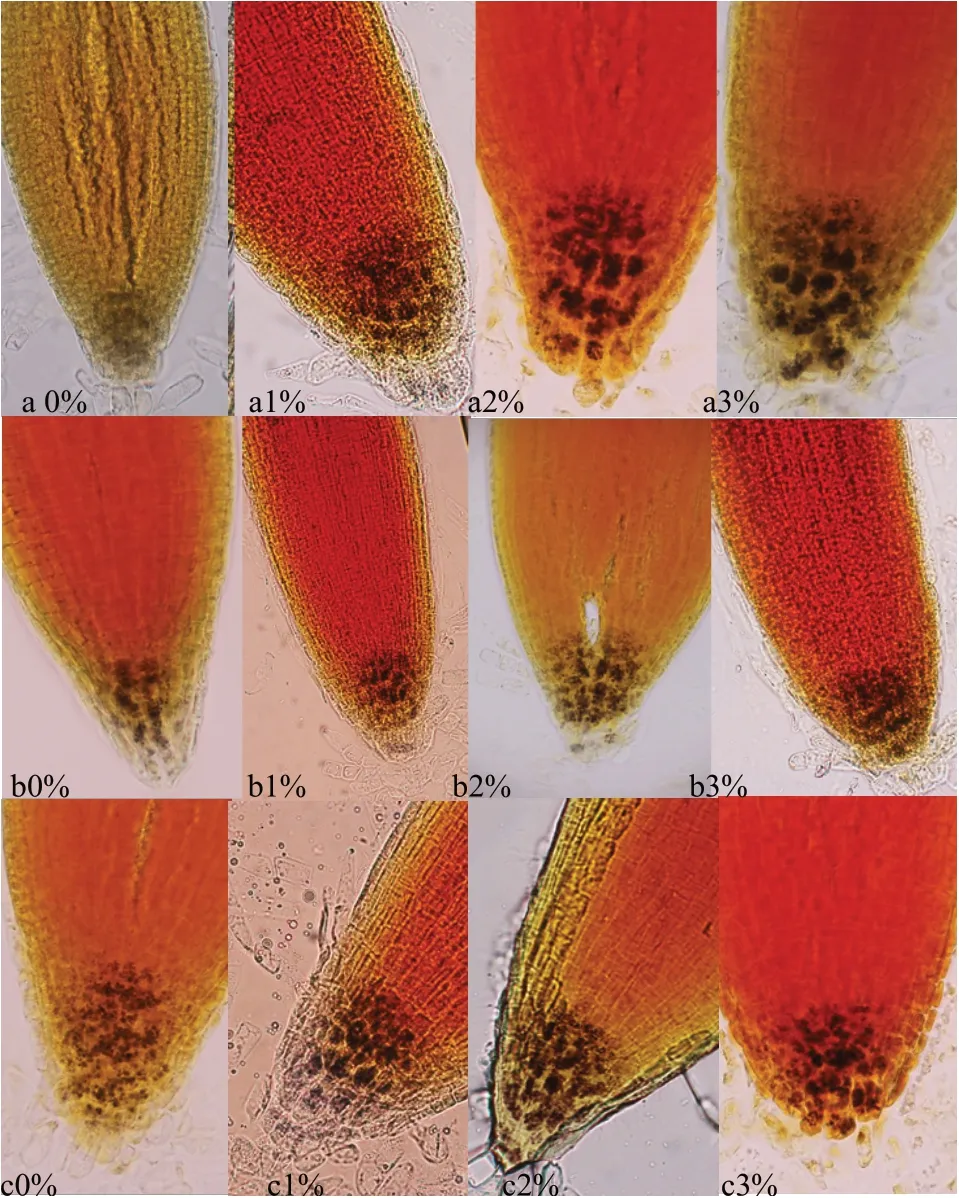
图1 蔗糖浓度不同根尖淀粉粒变化
3 讨论
前期研究发现,在添加蔗糖的培养基上培养拟南芥时,其根系会出现明显向左偏斜的行为。那么拟南芥根系的偏斜行为是否受蔗糖浓度的影响?氮的吸收转运是否参与蔗糖对根系偏斜的影响过程?本研究结果显示,蔗糖会影响根系生长的偏斜程度,无论是野生型还是突变体拟南芥,根系偏斜角度均表现为随蔗糖浓度的增加而增加,并在2%蔗糖处理达到最高,当蔗糖浓度达到3%时,野生型拟南芥Col-0根系倾斜角度显著降低,与不加蔗糖时根系倾斜角度差异不显著,而NRT1.1突变体chl1-5和nrt1.1-1根系倾斜角度无显著降低(见表2)。
在0%蔗糖处理时,野生型与突变体拟南芥的根系偏斜角度有显著性差异,突变体拟南芥根系的偏斜程度比野生型大。氮代谢与碳代谢紧密相连,光合碳代谢为氮代谢提供能量和碳架,氮代谢同碳代谢竞争光反应生成的同化力和中间代谢产物[9]。在低氮情况下进行光合作用的器官氮素降低,氮同化作用减弱,对光合作用的产物的竞争减弱[10]。研究中,NRT1.1突变体因降低了拟南芥根系对氮吸收的转运过程,因此,与野生型拟南芥相比NRT1.1突变体拟南芥对氮素同化作用减弱,对拟南芥光合作用产生的蔗糖消耗更少。1%-2%蔗糖处理时,野生型拟南芥根系偏斜角度比NRT1.1突变体拟南芥根系偏斜角度大,野生型与突变体拟南芥根系偏斜角度没有显著性差异。3%蔗糖处理,野生型与突变体拟南芥的根系偏斜角度有显著性差异,突变体拟南芥根系的偏斜程度比野生型大。综上所述,NRT1.1突变体根系偏斜程度受蔗糖浓度影响程度小于野生型,外源蔗糖可通过氮的吸收转化调控拟南芥根系的偏斜。
根系存在的向左偏斜行为可能是因为通过培养基中增加蔗糖浓度促进其根系生长,导致该现象扩大。但研究中同时对拟南芥生物量、主根长度进行相关性分析发现,与根系倾斜角度并未出现显著的相关关系。这也可能是因为根尖淀粉粒能影响植物根系对重力的感受[11],培养基中不同浓度蔗糖影响了根尖细胞淀粉粒的合成或分解,从而使根系偏斜角度增大。对根尖部位的淀粉粒进行染色观察可以发现,随着蔗糖浓度的增加,拟南芥根尖中淀粉粒是增加的。淀粉-平衡石假说中认为平衡石细胞中的淀粉体能够沿着重力方向沉降感受重力刺激,引起植物体内一系列信号分子的改变,使根的生长方向发生改变。当植物受到盐胁迫时根尖淀粉粒会被分解[11],降低根尖对重力的感知从而使根系生长方向发生改变。研究发现,培养基中增加蔗糖浓度会使根尖细胞中淀粉粒增加,但野生型与突变体并没有明显差异。这表明,外源蔗糖不是完全依靠改变根尖淀粉粒含量来感知重力,除根尖淀粉外,植物体内可能存在多种重力感受机制。
另外,已有研究表明,在根系生长发育过程中,葡萄糖可以通过调节生长素响应途径影响根系的定向响应。培养基中葡萄糖(Glc)的存在也广泛地调节了幼苗根系的生长方向[12]。葡萄糖诱导根的倾斜显著增强。葡萄糖作为糖信号与生长素、乙烯、油菜内酯素等植物激素共同相互作用控制葡萄糖诱导根倾斜[1,13-14]。故在培养基中添加不同浓度的蔗糖引起拟南芥根系倾斜变化,这可能不仅仅因为蔗糖影响了拟南芥根系中淀粉粒的形成,还可能是通过糖的渗透效应以及糖代谢引起的[10]。蔗糖被根系主动吸收后被降解为葡萄糖和果糖单体,通过葡萄糖含量的增加去调控根系的偏斜程度,但这一猜测还需进行进一步的试验验证。
4 结论
1)野生型与突变体拟南芥的根系偏斜角度均表现为随蔗糖浓度的增加而增加,并在2%蔗糖处理达到最高。
2)NRT1.1突变体根系偏斜程度受蔗糖浓度影响程度小于野生型,外源蔗糖可通过N的吸收转化调控拟南芥根系的偏斜。
3)不同浓度蔗糖处理下,无论野生型还是NRT1.1突变体拟南芥根系倾斜程度与生物量、根长均无显著正相关关系。
4)随着蔗糖浓度含量的增加,拟南芥根尖中淀粉粒逐渐增多。但野生型Col-0与NRT1.1突变体的分布和含量没有明显区别,外源蔗糖不是完全依靠改变根尖淀粉粒含量来感知重力。除根尖淀粉外,植物体内可能存在多种重力感受机制。
