扇贝新品系“紫海墨”F4的生长特征和生产性能分析
2022-03-02刘志刚王春德姚高友张柯馨展建强陈麟广吕文刚钟茂程刘付少梅
张 元,刘志刚,王春德,姚高友,张柯馨,展建强,陈麟广,吕文刚,钟茂程,刘付少梅
1.广东海洋大学水产学院,广东 湛江 524088
2.广东省海产无脊椎动物科技创新中心,广东 湛江 524088
3.中国科学院烟台海岸带研究所,山东 烟台 264003
4.湛江银浪海洋生物技术有限公司,广东 湛江 524022
墨西哥湾扇贝 (Argopecten irradians concentricus) 隶属于海湾扇贝属,为海湾扇贝 (A.irradians)的亚种之一,源自美国大西洋沿岸[1]。1991年中科院海洋研究所从美国将其引进,1996年推广至广东[1],2001年获得大面积养殖成功。近20年来,累代近亲繁殖已导致种质严重退化,呈现出个体小、生长速度慢和抗逆性下降等症状[2],对其进行遗传改良迫在眉睫。与海湾扇贝同属的紫扇贝 (A.purpuratus),是原产于南太平洋的速生、多产扇贝,其在智利北部和秘鲁沿岸已进行规模化的人工养殖[3]。2008年Wang等[4]将其引进,并将其与海湾扇贝杂交,经6代选育纯化,培育出扇贝新品种“渤海红”(Scallop "Bohai Red")[5]。该品种具有壳色紫红、生长速度快的特点[4,6-7],在山东、河北和辽宁获得大面积推广[5],取得显著的经济效益。但扇贝“渤海红”96 h半致死温度为29 ℃[5],无法直接在南方北部湾海域推广养殖。基于墨西哥湾扇贝与海湾扇贝互为亚种关系,将耐33 ℃高温[8]的墨西哥湾扇贝与经济性状优良但不耐高温的扇贝“渤海红”进行杂交,有可能获得杂种优势,并通过杂交育成,使优良性状得以纯化遗传。
有关扇贝遗传改良研究已有较多的报道,如扇贝的引种复壮[9-10]、遗传育种[11-13]等,至今通过国家原良种委员会审批的扇贝新品种有10个,包括栉孔扇贝“蓬莱红2号”[14]、“中科红”扇贝[15]、“海大金贝”[16]、“中科2号”海湾扇贝[17]、华贵栉孔扇贝“南澳金贝”[18]、扇贝“渤海红”[5]、虾夷扇贝“獐子岛红”[19]、海湾扇贝“海溢丰12”[20]、虾夷扇贝“明月贝”[21]、扇贝“青农金贝”[22]等,但通过墨西哥湾扇贝遗传改良获得的新品种尚未见报道。因此,2015年广东海洋大学开展了相关研究,成功获得杂交后代。从中选育出成活率高、生长速度快且耐高温的新品系“紫海墨”,以新品系“紫海墨”为材料进行了SSR标记的开发[23]、杂种优势、遗传结构的分析[24-25]、生长性能测定和温度耐受性[26]等方面的研究。结果表明,新品系“紫海墨”拥有2个祖代基因,生长速度比墨西哥湾扇贝快,其96 h半致死温度为32.4 ℃[24]。本研究对2019年继代选育获得的新品系“紫海墨”F4(图1-a) 与墨西哥湾扇贝 (图1-b) 的经济性状进行对比,以期为新品系“紫海墨”F4的选育效果提供依据,为下一代选育提供方向。新品系“紫海墨”F4通过后续选育,有望成为又一个新品种扇贝。

图1 新品系“紫海墨”F4 (a) 和墨西哥湾扇贝 (b)Fig.1 New strain "Zihaimo" F4 (a) and A.irradians concentricus (b)
1 材料与方法
1.1 实验材料
新品系“紫海墨”F4和北部湾养殖的墨西哥湾扇贝的出池种苗同时由湛江银浪海洋生物技术有限公司提供,养殖对比实验点位于遂溪县江洪镇扇贝养殖区。实验期间每月定期测量养殖区的水温和比重,同时从养殖区随机取样3个养殖笼,当场统计该阶段各笼成活率,然后将3个养殖笼的扇贝均匀混合,随机抽样50个存活个体,用冰袋保持低温干露运输至广东海洋大学海产无脊椎动物科技创新中心实验室暂养,清除扇贝表面的附着物,洗刷干净并用纱布吸干表面的水分,进行生长数据测量。
1.2 选育和养殖方法
扇贝“渤海红”于2015年从山东引进,墨西哥湾扇贝为北部湾养殖群体,按壳长留选率5%上选,两者混交得到杂种。对杂种进行继代选育,每代选育目标首先为橙黄色个体,二选在橙黄色个体中,按壳长留选率5%上选,2016年得到选育系F1,2017年得到F2,2018年得到F3,2019年得到F4。
2019年9月11日,对新品系“紫海墨”F3按上述方法进行继代选育,于2019年10月13日 (水温为29.3 ℃) 育出壳长1~1.5 mm 新品系“紫海墨”F4和壳长1~1.5 mm墨西哥湾扇贝稚贝。两种稚贝分别采用60目网袋,按每袋3 000粒的密度下海中培20 d,中培期间水温25.9~29.3 ℃、盐度31.1~31.2、pH 8.1~8.2、透明度2.3~3.2 m;11月2日结束一级中培。将平均壳高5.2 mm新品系“紫海墨”F4和平均壳高3.7 mm墨西哥湾扇贝稚贝分别移入40目网袋进行二级中培,密度为每袋500粒;在11月22日新品系“紫海墨”F4贝苗的平均壳长为15.75 mm,即移入中培笼 (网孔8 mm、每笼8层),按每层200粒的密度进行三级中培;在12月18日墨西哥湾扇贝稚贝的平均壳长为12.93 mm,也放入同规格的笼具中进行三级中培;同期,新品系“紫海墨”F4稚贝的平均壳长达到27.65 mm,即移入养成笼 (网孔2.5 cm、每笼10层) 套网养成,密度为每层50粒,10 d后壳长大于3 cm时去掉套网进入养成。2020年1月14日墨西哥湾扇贝平均壳长达到16.93 cm,放入同样规格的笼具中养殖,30 d后贝苗平均壳长大于3 cm时去掉套网进入养成。为达到随机抽样的目的,同一群体各阶段不同大小个体均不分规格养殖。
1.3 数据测量
将壳长大于300 μm而小于3.0 mm 的稚贝在显微镜目微尺 (25×) 下进行测量,壳长大于3.0 mm的幼贝和亲贝使用电子游标卡尺测量,测量指标包括壳长、壳高和壳宽3个性状,精确到0.01 mm;使用电子天平测量扇贝的体质量、闭壳肌质量和壳质量,精确到0.01 g;成活率的测定方法是随机抽样3笼,倒出贝苗,分别统计各笼新品系“紫海墨”F4与墨西哥湾扇贝的成活率。
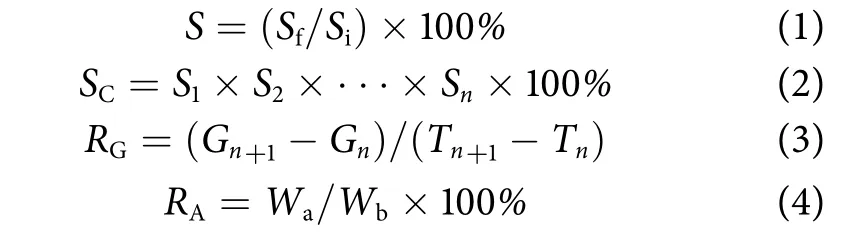
式中:S为成活率;Sf、Si分别表示实验时各阶段存活个体数、该阶段投苗总数;SC为累积成活率 (%);S1、S2分别为第一、第二阶段成活率,Sn为第n阶段成活率;RG为各性状日增长率 (μm·d−1/mg·d−1);Gn、Gn+1为该实验阶段起始、结束时各性状的数据;Tn、Tn+1为测量起始、结束的时间;n为该实验阶段序号 (n≥1);RA为肉柱率 (%),Wa为闭壳肌质量;Wb为体质量。
1.4 数据处理
采用 Excel 2010和SPSS 23.0软件进行数据处理与分析。通过直方图和Q-Q图检验方法对生长数据进行正态分布分析。用t检验法对结果进行显著性分析,P<0.05为差异显著,P<0.01为差异极显著。用Excel 2010软件作图,所有数据均以“平均值±标准差 (SD)”表示。
2 结果
2.1 成活率对比分析
2019年10月13日,将壳长1~1.5 mm的新品系“紫海墨”F4与墨西哥湾扇贝贝苗投放到遂溪县江洪镇进行对比养殖,实验期间温度情况见图2,温度介于16.7~29.3 ℃。养殖期内最低温度出现在1月 (16.7 ℃),最高出现在10月 (29.3 ℃)。
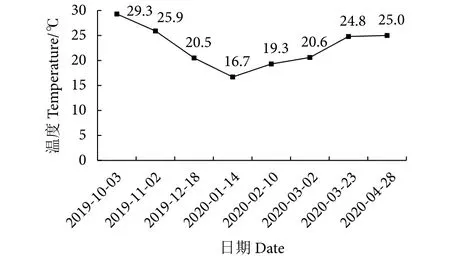
图2 养殖期间温度变化Fig.2 Variation in temperature during breeding
新品系“紫海墨”F4和墨西哥湾扇贝各阶段的成活率见图3-a。两个养殖群体同阶段的成活率有极显著差异 (P<0.01);新品系“紫海墨”F4的成活率在各时间段均高于墨西哥湾扇贝;两个养殖群体的阶段成活率最低均出现在11月的第一次中培,分别为56.2%和44.6%,这可能与刚下海的稚贝弱小、抗逆性差、淘汰较多有关;之后随着贝苗的生长,两个养殖群体的阶段成活率均明显提高,新品系“紫海墨”F4尤其显著。两个养殖群体的累积成活率变化见图3-b,随着养殖时间的延长,两个养殖群体均缓慢下降,但未出现断崖式下跌或急剧死亡,且在各个阶段新品系“紫海墨”F4的累积成活率均高于墨西哥湾扇贝,新品系“紫海墨”F4最终累积成活率为51.32%,显著高于墨西哥湾扇贝的29.18% (P<0.01)。
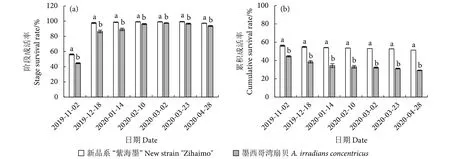
图3 新品系“紫海墨”F4和墨西哥湾扇贝各阶段成活率和累积成活率变化Fig.3 Variation in survival rate and cumulative survival rate of new strain "Zihaimo" F4 and A.irradians concentricus
2.2 新品系“紫海墨”F4与墨西哥湾扇贝生长特征比较
2.2.1 新品系“紫海墨”F4壳长的错峰生长
在相同的时间节点,新品系“紫海墨”F4的壳长均大于墨西哥湾扇贝,两个养殖群体间有显著差异 (P<0.05);两个养殖群体从2019年10月13日初始壳长1~1.5 mm开始,经过6.5个月的养殖,前者壳长达到58.33 mm,后者仅48.81 mm,前者比后者提高了19.50% (图4-a)。新品系“紫海墨”F4的壳长增长率高峰 (0.48 mm·d−1) 出现在1月14日,而墨西哥湾扇贝 (0.45 mm·d−1) 却延迟至3月2日。两个养殖群体的壳长出现了错峰生长情况,原因可能与其温度适应性有关,系祖扇贝“渤海红”的适应低温基因可能在此产生了作用。此外,新品系“紫海墨”F4在3月23日的壳长增长率最低,与后述体质量增长率较高存在不对应情况(图4-b)。这可能与此时期大量营养用于生殖腺发育和增肉有关,导致其壳长生长变缓,但墨西哥湾扇贝不存在这一情况。
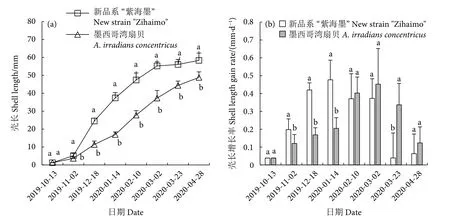
图4 新品系“紫海墨”F4和墨西哥湾扇贝壳长生长和壳长增长率Fig.4 Shell length growth and shell length gain rate of new strain "Zihaimo" F4 and A.irradians concentricus
2.2.2 新品系“紫海墨”F4壳高的错峰生长
两个养殖群体壳高的生长趋势与壳长一致(图5-a)。在相同的时间节点,新品系“紫海墨”F4的壳高均大于墨西哥湾扇贝,两个养殖群体间有显著差异 (P<0.05)。两个养殖群体从2019年10月13日初始壳高1~1.5 mm开始,经过6.5个月的养殖,新品系“紫海墨”F4的壳高为55.81 mm,后者仅46.57 mm,前者比后者提高了19.84%。新品系“紫海墨”F4的壳高增长率高峰出现在1月14日 (0.42 mm·d−1),而墨西哥湾扇贝则延迟至3月2日 (0.44 mm·d−1),两者的壳高出现错峰生长情况,与壳长的表现相同 (图5-b)。此外,新品系“紫海墨”F4在3月23日壳高增长率最低,与后述体质量增长率较高存在不对应情况,原因可能也与壳长的相同。
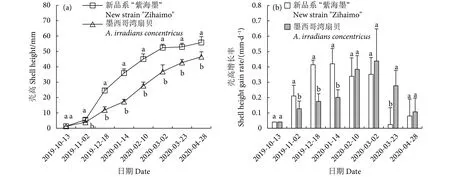
图5 新品系“紫海墨”F4和墨西哥湾扇贝壳高生长和壳高增长率Fig.5 Shell height growth and shell height gain rate of new strain "Zihaimo" F4 and A.irradians concentricus
2.2.3 新品系“紫海墨”F4壳宽的错峰生长与延缓现象
两个养殖群体壳宽的生长趋势与壳长和壳高基本一致 (图6-a)。不同的是,新品系“紫海墨”F4的壳长、壳高增长率均在1月14日达到最大值,而壳宽增长率则延迟至2月10日才达到最大值 (0.24 mm·d−1)。这与该时期扇贝开始增肉,使壳宽生长加快有关,与后述体质量增长率的变化趋势同步;而墨西哥湾扇贝的壳长、壳高、壳宽增长率均在3月2日达到最大值,不存在壳宽生长延缓的情况 (图6-b)。
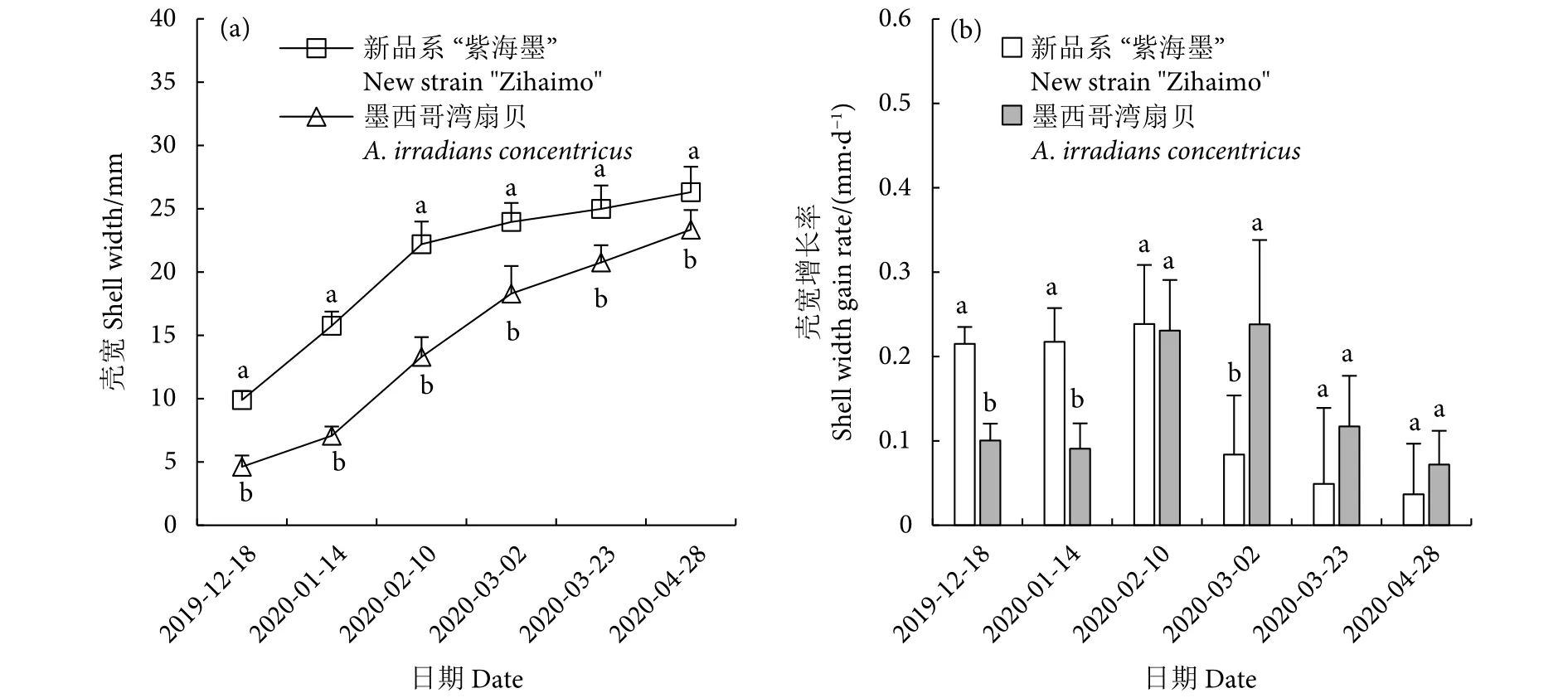
图6 新品系“紫海墨”F4和墨西哥湾扇贝壳宽生长和壳宽增长率Fig.6 Shell width growth and shell width gain rate of new strain "Zihaimo" F4 and A.irradians concentricus
2.2.4 新品系“紫海墨”F4体质量的错峰生长与延缓现象
两个养殖群体的体质量增长趋势与壳长、壳高基本一致 (图7-a)。但有一点不同,新品系“紫海墨”F4的壳长、壳高增长率均在1月14日达到最大值,而体质量增长率则延迟至2月10日才达到最大值 (0.33 g·d−1),这与此时期壳性状生长减缓而开始增肉,导致体质量增长加快有关,与上述壳宽增长率变化趋势对应;但墨西哥湾扇贝壳长、壳高、壳宽及体质量增长率均在3月2日达到最大值,不存在体质量延缓情况 (图7-b)。
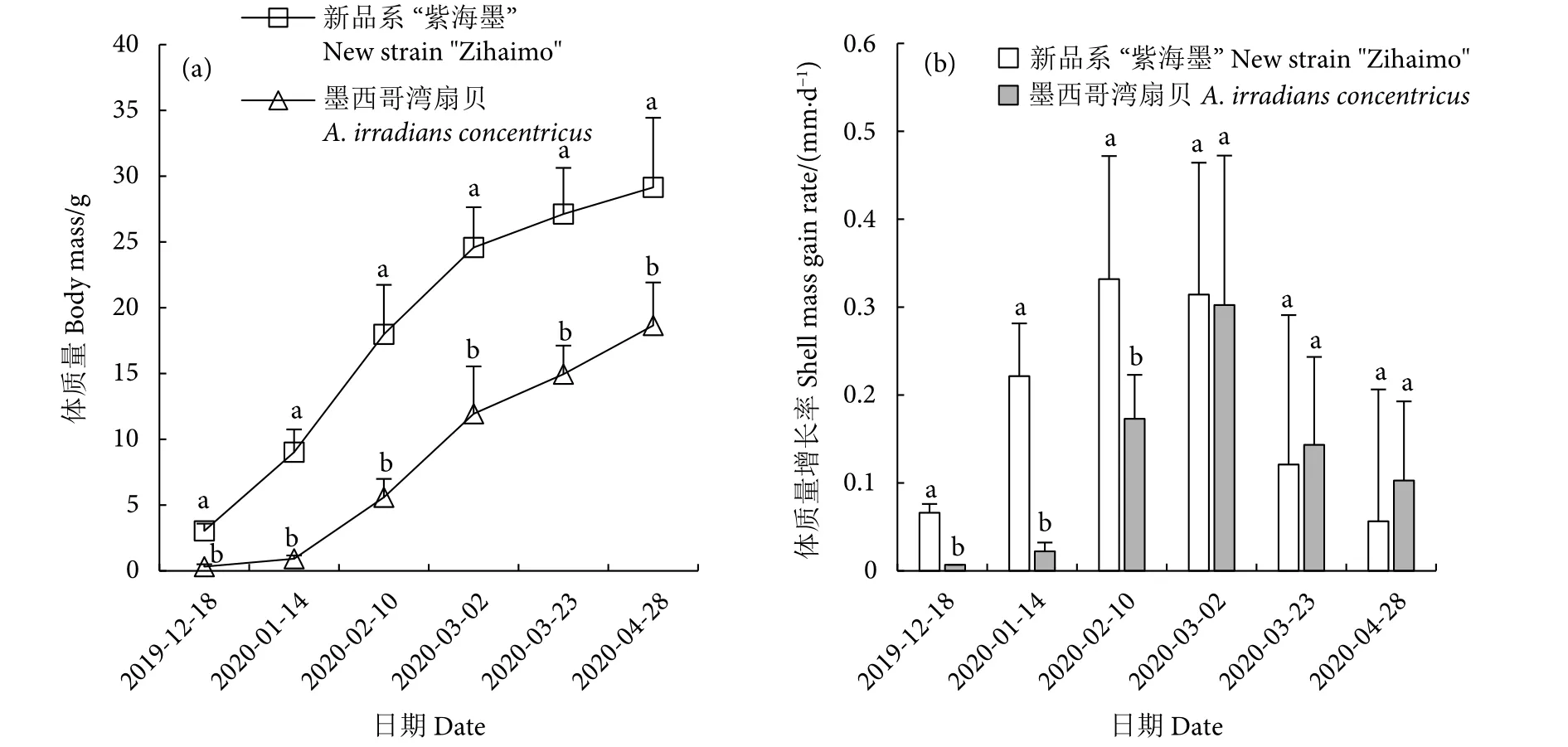
图7 新品系“紫海墨”F4和墨西哥湾扇贝体质量增长和增长率Fig.7 Shell mass growth and shell mass gain rate of new strain "Zihaimo" F4 and A.irradians concentricus
2.3 新品系“紫海墨”F4与墨西哥湾扇贝的质量性状比较
养殖141 d (4.7个月),新品系“紫海墨”F4的体质量为24.58 g,闭壳肌平均质量为3.38 g;墨西哥湾扇贝体质量为11.94 g,闭壳肌平均质量仅1.32 g。养殖198 d (6.6个月),新品系“紫海墨”F4的体质量为29.15 g,闭壳肌平均质量为3.69 g;墨西哥湾扇贝的体质量为18.65 g,闭壳肌质量为3.07 g (图8-a)。若收获规格以墨西哥湾扇贝为标准,即约25 g收获,则新品系“紫海墨”F4的养殖周期比墨西哥湾扇贝缩短约2个月。
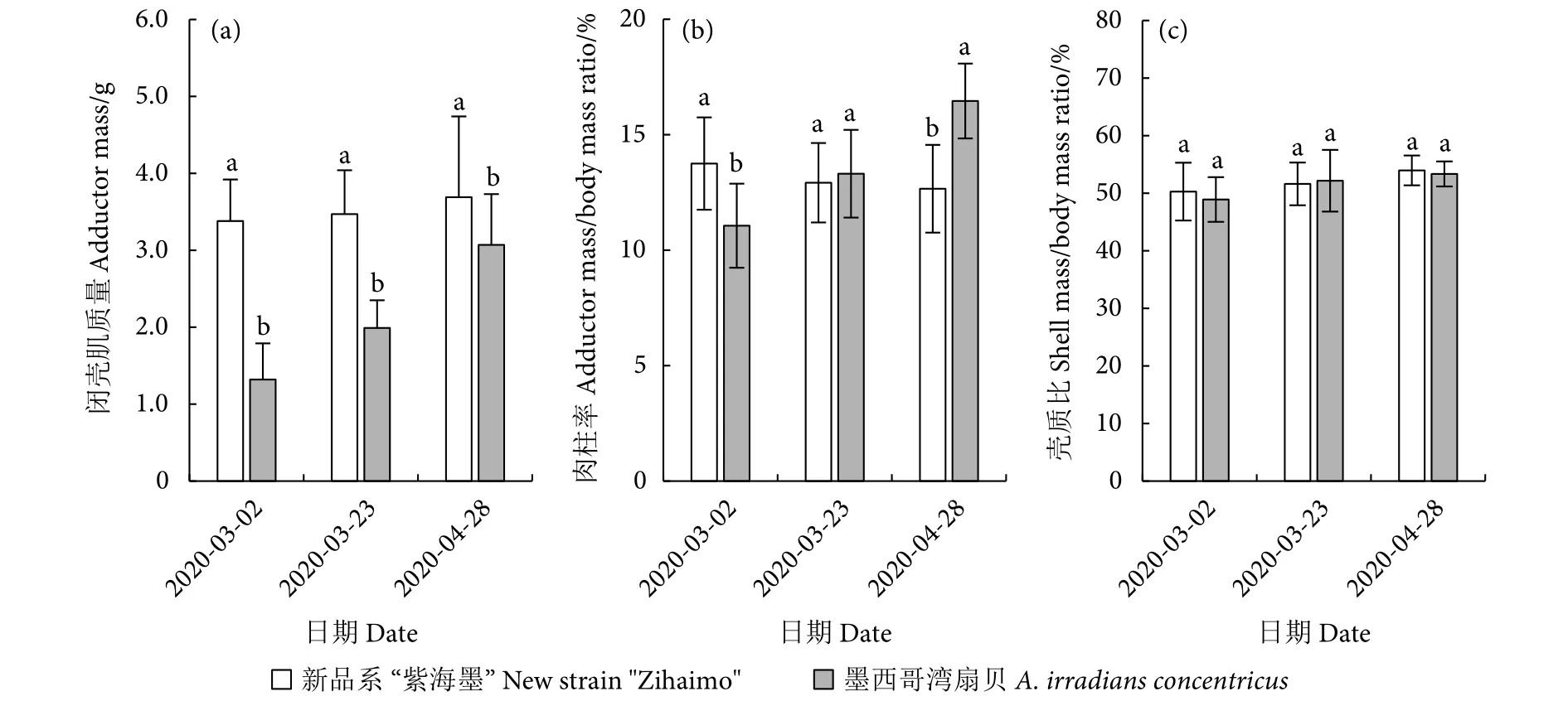
图8 新品系“紫海墨”F4和墨西哥湾扇贝闭壳肌质量、肉柱率和壳质比Fig.8 Adductor mass, adductor mass/body mass ratio and shell mass/body mass ratio of new strain "Zihaimo" F4 and A.irradians concentricus
新品系“紫海墨”F4的肉柱率在养殖第141天 (4.7个月) 时最高,后开始下降,因此以5个月内收获为佳;墨西哥湾扇贝肉柱率在养殖第198天 (6.6个月) 时才达到最高 (图8-b),表明其闭壳肌生长速度滞后于新品系“紫海墨”F4,但肉柱率却高于后者;新品系“紫海墨”F4和墨西哥湾扇贝的平均肉柱率分别为13.07%和13.61%。
新品系“紫海墨”F4的壳质比略大于墨西哥湾扇贝,表明新品系“紫海墨”F4的肉柱率略低于墨西哥湾扇贝。新品系“紫海墨”F4和墨西哥湾扇贝的壳质比均呈现上升趋势,但上升程度较缓慢,最终比值分别为53.96%和53.35% (图8-c)。
3 讨论
3.1 新品系“紫海墨”F4的成活率
本研究结果表明,新品系“紫海墨”F4在各培养阶段的成活率均显著高于墨西哥湾扇贝,且最终累积成活率 (51.32%) 极显著高于墨西哥湾扇贝(29.18%,P<0.01)。这可能与新品系“紫海墨”F4种质有关,其是来自两个不同品种杂交选育产生的后代,多样性和杂合度均高于墨西哥湾扇贝[25]。
墨西哥湾扇贝经过20多年的养殖,小群体繁殖和近亲交配退化现象严重[2],成活率下降较为突出。Zhao等[27]研究指出,双壳类的近交会使生物的适应性下降,相比于杂交其死亡率更高;张国范等[28]研究发现,海湾扇贝的近亲交配会导致发育迟缓和个体小型化现象;扇贝的子代数量足够多,一定程度上可以避免近交效应,近交效应导致种质退化,极有可能是因为出现瓶颈效应 (Bottleneck effect) 时与有效繁殖群体大小、子代个体数、交配方式和近交代数有关;并指出目前海区扇贝大规模死亡的原因为养殖环境胁迫对扇贝的生理机能造成影响,形成环境-遗传的综合效应[29]。以往研究表明,影响贝类成活率的因素涉及自身的免疫性[30-31]和对环境的适应性[32-33]。Thomas等[34]研究发现欧洲大扇贝 (Pecten maximus) 幼贝更喜欢以地板砖为附着物;Arturo等[35]研究表明缺氧对秘鲁紫扇贝的壳生长有影响,长期暴露于缺氧环境对扇贝的生长和繁殖造成影响[36];张元等[37]研究发现温度和盐度的互作效应对扇贝“渤海红”稚贝的存活有影响。本研究表明,养殖初期即11月第一次中培时的成活率较低,新品系“紫海墨”F4和墨西哥湾扇贝分别为56.2%和44.6%,这既与出苗时的机械损伤有关,也与出池稚贝个体弱小、环境适应能力低弱有关,随着养殖时间的延长,扇贝自身免疫力和对环境的适应能力均有所增强,成活率明显提高。新品系“紫海墨”F4的成活率高于北部湾养殖多年的墨西哥湾扇贝,本研究表明其已具备在当地海域进行规模化养殖的条件。
3.2 新品系“紫海墨”F4的生长性能
本研究表明新品系“紫海墨”F4的选育效果显著,其壳长、壳高、壳宽、体质量和闭壳肌质量比墨西哥湾扇贝分别提高了19.50%、19.84%、16.23%、56.30%和20.20%,且其养殖周期仅4.7个月即可达商品规格,而墨西哥湾扇贝则超过6.6个月。新品系“紫海墨”F4的生长优势与其遗传背景密切相关,它是扇贝“渤海红”与墨西哥湾扇贝杂交选育的第四代。杂交把两者的基因库融汇,汇聚了双方的优点,增加了新品系“紫海墨”F4的遗传多样性和杂合度。杂交子代经过若干代的闭锁群体选育,剔除了不适合高温环境和人类需求的基因型,保留并纯化经济性状优良的基因型,使选育后代具备了一定的形态特征和优异的经济性状,其中生长性状就是重要的经济指标。虽然杂种优势的遗传机制尚未明确[38],但已经提出了几种经典假设来解释杂种优势的遗传原因[39]。杂交育种被认为是动、植物遗传改良最有效的方法之一,被广泛运用于扇贝遗传育种中。风向标扇贝 (Patinopecten caurinus) 和虾夷扇贝 (P.yessoensis) 杂交子代的生长速度显著优于亲本[40];鳞砗磲 (Tridacna squamosa) 和圆砗磲 (T.crocea) 杂交子代的成活率、壳长和湿质量优于亲本[41];长牡蛎 (Crassostrea gigas) 和日本牡蛎 (C.nippona) 的杂交子代表现出生存与杂种优势[42];近江牡蛎 (C.ariakensis) 和香港牡蛎 (C.hongkongensis) 杂交F2子代在高盐地区比亲本香港牡蛎表现出较高的耐盐性和生长速度[43];Han等[44]研究表明3种颜色长牡蛎杂交后稚贝的成活率比亲本提高8.22%~40.91%;对墨西哥湾扇贝与华贵栉孔扇贝 (Chlamys nobilis) 进行远缘杂交,其D形幼虫的壳长、壳高的生长速度明显高于亲本,具有明显的杂交优势[45]。经过 5个世代的针对性选育,吉富罗非鱼 (Oreochromis niloticus) 的抗病和生长性能均获得明显改善,选育效果显著[46];李振通等[47]研究显示驼背鲈 (Chromileptes altivelis♀)×蓝身大斑石斑鱼 (Epinephelus tukula♂) 远缘杂交组合是可行的,且杂交子代表现出生长速度快的杂交优势。通过高通量测序的方式,分析杂交子代与亲本的差异表达基因,表明上调基因参与了线粒体的形成和能量代谢过程,推测其极有可能与提高能量促进杂交贝的生长有关[13]。
3.3 两个养殖群体生长的温度错位效应
新品系“紫海墨”F4的壳长、壳高增长率高峰均出现在1月 (16.7 ℃),而墨西哥湾扇贝则延迟至3月 (20.6 ℃),这可能与两者的温度适应性和遗传背景有关。新品系“紫海墨”F4的遗传背景一方来自北方养殖的新品种扇贝“渤海红”,其半致死温度为29.0 ℃,另一方来自北部湾养殖的墨西哥湾扇贝,其小贝的半致死温度为32.67 ℃[48];新品系“紫海墨”F3的最适生长温度为22 ℃[24],墨西哥湾扇贝为24~28 ℃[49]。新品系“紫海墨”F4在16.7~25 ℃内生长速度更快,生长适温更倾向于扇贝“渤海红”,而墨西哥湾扇贝则在19.3~25 ℃内生长迅速。此外,新品系“紫海墨”F4的壳宽、体质量增长率则延迟至2月10日才达到最大值,这与此时期开始增肉且体质量增长加快有关,体质量增长导致其体积膨胀,并促使壳宽快速生长以适应软体质量增长的需要。这一结果与王辉等[50]和刘志刚等[51]报道的墨西哥湾扇贝体质量增长规律相同。程操[52]通过新品种“明月贝”与对照组虾夷扇贝在室内高温模拟与夏季室外验证的方式,得出新品种“明月贝”有一定的高温耐受性。
4 小结
本研究表明,经过6.5个月同一海区的养殖,新品系“紫海墨”F4选育效果显著。新品系“紫海墨”F4和墨西哥湾扇贝的累积成活率分别为51.32%和29.18%,前者比后者提高了75.87%;壳长、壳高、壳宽、体质量和闭壳肌质量分别比墨西哥湾扇贝分别提高了19.50%、19.84%、16.23%、56.30%和20.20%;墨西哥湾扇贝生长明显滞后于新品系“紫海墨”F4。在接下来的选育工作中,将把新品系“紫海墨”F4于不同海区进行养殖,同时探讨其基因与环境的互作效应,以便检验新品系“紫海墨”在南方海域不同养殖地点的适应性及推广优势。
