一株侵染草地贪夜蛾成虫的球孢白僵菌的分离鉴定
2022-02-10庞继鑫温绍海杜广祖杨航马芸杨进波付玉飞张庭发易小光李正跃陈斌
庞继鑫 温绍海 杜广祖 杨航 马芸 杨进波 付玉飞 张庭发 易小光 李正跃 陈斌


摘要
草地贪夜蛾Spodoptera frugiperda(J. E. Smith)是一种全球性重大玉米害虫,于2019年入侵我国云南。2020年8月,从云南省大理弥渡西庄村玉米田采集被虫生真菌感染的草地贪夜蛾雌成虫,经在室内分离培养,结合病原菌分生孢子与菌丝等形态及ITS序列相似性分析,确定感染草地贪夜蛾成虫的真菌为球孢白僵菌Beauveria bassiana,菌株编号为SHL2020。经生物测定, 该菌对草地贪夜蛾雌、雄成虫均具有较强的侵染致病作用。用该菌1×108个/mL 分生孢子菌悬液接种处理后第7天时,草地贪夜蛾雌、雄成虫累计死亡率分别为90.0%和86.7%。以上结果表明,从罹病草地贪夜蛾雌成虫上分离获得的球孢白僵菌SHL2020菌株对草地贪夜蛾雌雄成虫具有良好的生防潜力,该研究为草地贪夜蛾生物防治提供了理论依据和物质基础。
关键词
球孢白僵菌;草地贪夜蛾;昆虫病原真菌;致病力
中图分类号:
S476.12
文献标识码:A
DOI:10.16688/j.zwbh.2020587
Isolation and identification of an entomopathogenic fungus infecting the adult of Spodoptera frugiperda (J. E. Smith)
PANG Jixin1#,WEN Shaohai1#,DU Guangzu1,YANG Hang2,MA Yun1,YANG Jinbo2,
FU Yufei1,ZHANG Tingfa2,YI Xiaoguang2,LI Zhengyue1*,CHEN Bin1*
(1. State Key Laboratory for Conservation and Utilization of Bioresources in Yunnan,
College of Plant Protection, Yunnan Agricultural University, Kunming650201, China;
2. Yunnan Green Biotechnology Science and Technology Co. Ltd., Dali675600, China)
Abstract
Spodoptera frugiperda (J. E. Smith) is a global major pest and invaded into Yunnan province in 2019. An infected female adult of S.frugiperda was found in corn planting field in Xizhuang village in Midu county of Dali city, Yunnan province in August 2020, and was identified by combining morphology and ITS sequence analysis. The pathogenicity of the fungus to male and female adult of the S.frugiperda was determined by conidia suspension spray method. The results indicated the fungus was identified to be Beauveria bassiana, which was named as SHL2020. The fungus was pathogenic to both male and female adults, and the cumulative adjusted mortality was 86.7% and 90.0% at the 7th day after infection when the spore concentration was 1×108 spore/mL. The above results indicated that B.bassiana SHL2020 fungus isolated from the female adult S.frugiperda has good biocontrol potentiality to the male and female adults of S.frugiperda, which provide the theoretical and material base for the development of the biocontrol method for S.frugiperda.
Key words
Beauveria bassiana;Spodoptera frugiperda;entomopathogenic fungus;pathogenicity
草地貪夜蛾Spodoptera frugiperda (J.E.Smith), 也称秋黏虫,属于鳞翅目Lepidoptera,夜蛾科Noctuidae, 灰翅夜蛾属Spodoptera[1],原产于美洲热带和亚热带地区,是一种为害玉米的重大迁飞性害虫[2- 3]。 该虫具有寄主范围宽、适生区域广、迁飞能力强、扩散速度快和突发为害重等特点[4],目前,全球已有100多个国家报道了草地贪夜蛾入侵为害[5]。2019年1月,在我国云南省普洱市江城县宝藏镇首次发现草地贪夜蛾以来,该虫在我国迅速扩散,并在玉米种植区大面积发生,成为我国玉米生产的重要潜在危害因素[6 -8]。当前国内主要采用化学药剂防治
草地贪夜蛾,在农业农村部推荐的25种用于草地贪夜蛾应急防控的杀虫药剂中,有19种为化学杀虫剂[9]。但由于该虫在进入我国之前长期的化学防治已导致其对多种传统杀虫药剂产生了抗性[10 -11],而且长期使用化学农药易伤害非靶标生物,破坏生态平衡,因此化学防治受到了一定的限制。自该虫入侵我国以来,在国内已有很多有关天敌防治草地贪夜蛾的相关研究[12 -17],而对该虫具有高毒力的病原真菌的相关报道较少。
目前,在昆虫病原真菌防控草地贪夜蛾方面,郑亚强等[18]在云南省草地贪夜蛾入侵当年最先发现了田间自然罹病的草地贪夜蛾幼虫,经过室内分离鉴定菌株,确定了其为莱氏绿僵菌Metarhizium rileyi,且该菌对草地贪夜蛾幼虫具有高致病性;魏雪菲等[19]调查发现莱氏绿僵菌在昆明地区玉米田草地贪夜蛾幼虫种群中发生且流行性强。国内外已有学者研究表明金龟子绿僵菌M.anisopliae和球孢白僵菌Beauveria bassiana 对草地贪夜蛾低龄幼虫、蛹、卵都具有一定的防治效果[20 -21];雷妍圆等[22]发现玫烟色虫草Cordyceps fumosorosea对草地贪夜蛾幼虫具有较高
的致病性。已报道的菌株均是对草地贪夜蛾卵和低龄幼虫有侵染致病作用,而关于草地贪夜蛾成虫的病原真菌还未见相关报道。
白僵菌 Beauveria spp. 是最常见的昆虫病原真菌之一,在我国现有的白僵菌中,球孢白僵菌因其广谱性,是目前被用于研制真菌杀虫剂最多的虫生真菌[23],在田间害虫生物防治中发挥了极其重要的作用。自从1956年在我国首次报道了利用白僵菌防治甘薯象甲Cylas formicarius Fabricias后[24],我国在此方面的研究越来越多,已经广泛应用于马尾松毛虫Dendrolimus punctatus Walker、稻水象甲Lissorhoptrus oryzophilus Kuschel、红火蚁Solenopsis invicta Buren等害虫的防治中,并取得了较好的防治效果[25 -27]。
本研究从云南省大理弥渡西庄村玉米田采集的罹病草地贪夜蛾雌成虫上分离纯化获得1株病原真菌,通过形态学和ITS序列分析对该菌株进行了鉴定,并初步测定了该菌株对草地贪夜蛾雌、雄成虫的致病性,以期为开发真菌杀虫剂绿色防控草地贪夜蛾成虫提供备选菌株和利用昆虫病原真菌进行防治提供理论依据。
1材料与方法
1.1供试菌株与虫源
2019年8月4日,于云南省大理弥渡西庄村玉米田发现罹病草地贪夜蛾成虫,25°37′23″N,100°41′19″E,海拔1 700 m。
1.2菌株分离与纯化
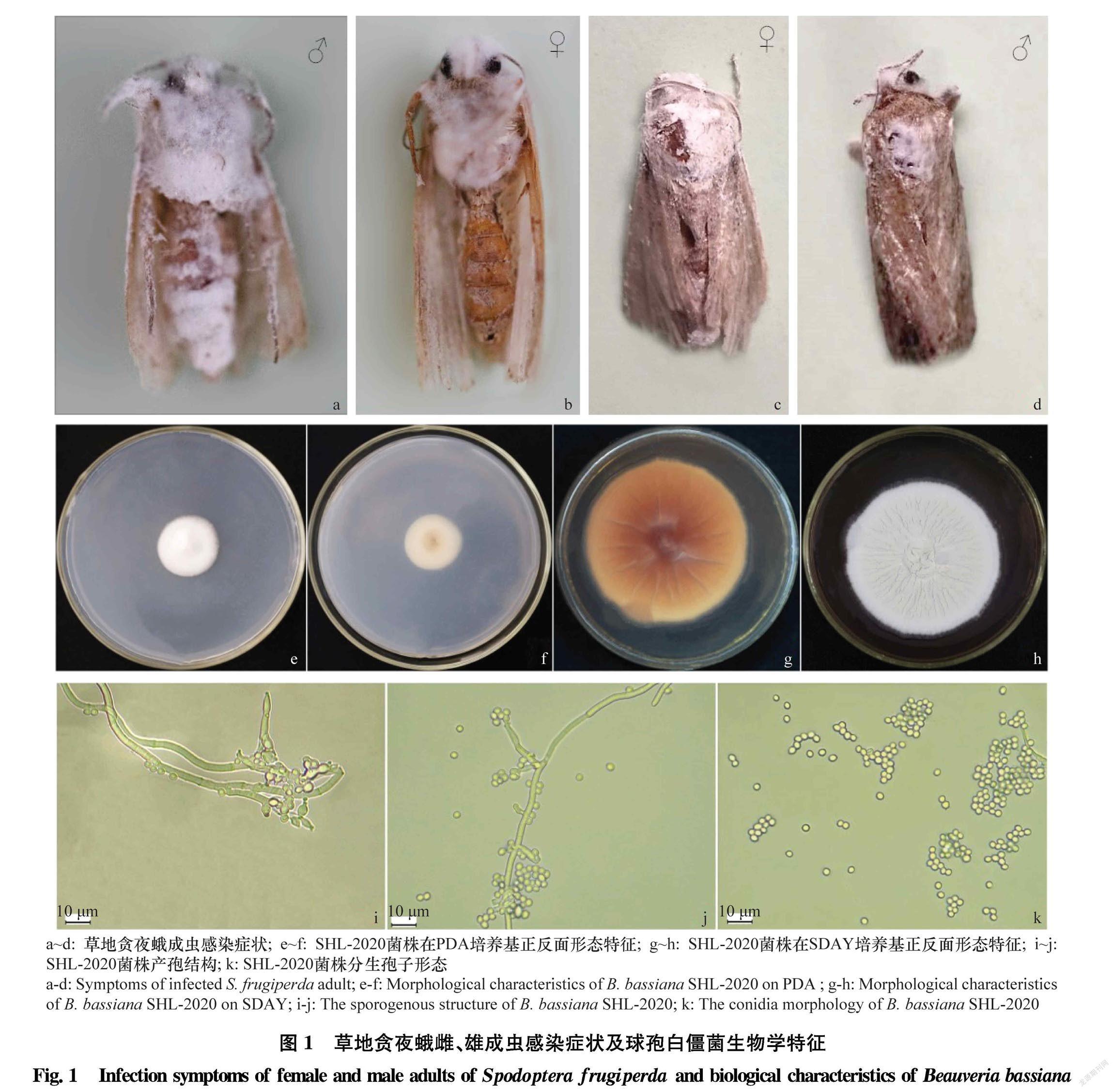
将罹病的草地贪夜蛾成虫(图1a~d)置于灭菌培养皿(d=9 cm)中,在(28±1)℃恒温培养箱中保湿培养,待其分生孢子在虫体表面明显生长后,在消毒灭菌的超净工作台中利用接种环从感病成虫虫体表面轻轻挑取少量孢子,采用抖动法将其接种于萨氏培养基SDAY(葡萄糖40 g,蛋白胨10 g,酵母浸膏粉10 g,琼脂20 g,pH自然)[28]上,于(25±1)℃,相对湿度(70±1)%,L∥D=14 h∥10 h的恒温恒湿光照培养箱中培养[29]。培养5 d后,挑取无污染的菌丝或孢子接种至新的培养基进行纯化培养,直至平板上长出的菌落形态特征一致,将该菌株编号为SHL2020。待纯化培养的菌株充分产孢后,在4℃冰箱中储存备用。
1.3菌株培养形态观察
将分离纯化后的SHL2020菌株分别接种于SDAY和PDA培养基(马铃薯200 g,葡萄糖20 g,琼脂20 g,蒸馏水1 L)中间,于(25±1)℃,相对湿度(70±1)%,L∥D=14 h∥10 h光照培养箱(宁波赛福实验仪器有限公司,型号ZRY380)中培养。在接菌后第3、7和14天时,分别测量不同培养基上菌落的直径并观察菌落的形态特征[18]。采用光学显微镜(Leica, 750)观察菌株产孢结构和分生孢子形态等显微结构。
1.4菌株分子生物学鉴定
采用李焕宇等[30]改良的CTAB法,以菌株SHL2020基因组DNA为模板,采用真菌通用引 物ITS1(5′TCCGTAGGTGAACCTGCGG3′) 和 ITS4(5′TCCTCCGCTTATTGATATGC3′)进行菌株ITS序列PCR扩增。PCR反应体系为25 μL,其中上下游引物ITS1和ITS4各1 μL,模板DNA 1~2 μL,PCR MasterMix 12.5 μL,最后ddH2O补足至25 μL。PCR反应条件:95℃预变性3 min;94℃变性1 min, 55℃退火1 min,72℃延伸1.5 min, 35个循环;72℃延伸10 min。1%琼脂糖凝胶电泳检测。PCR产物送生工生物工程(上海)股份有限公司测序。
1.5系统发育分析
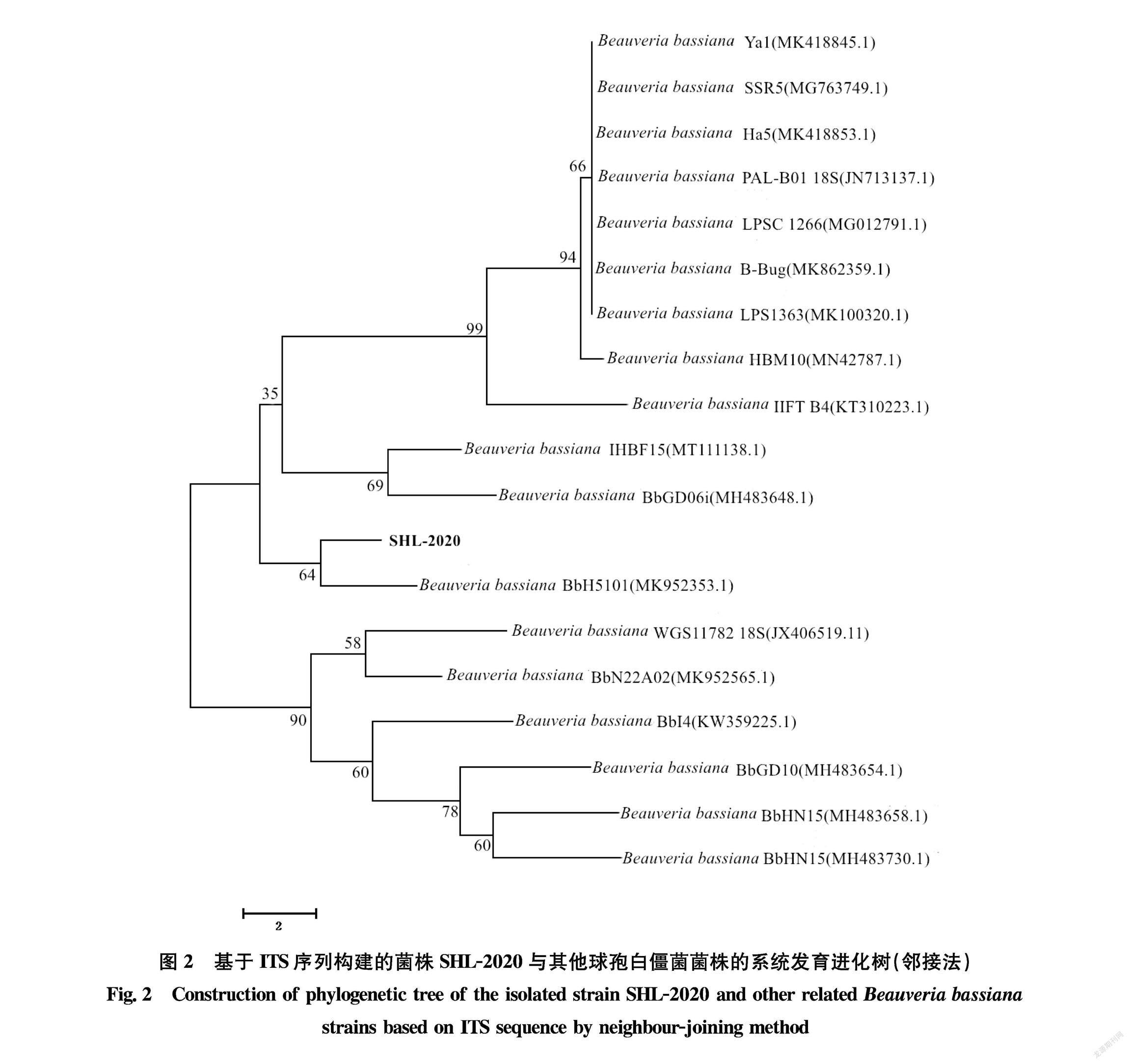
将该目的菌株的ITS序列结果提交至NCBI网站 (https:∥blast.ncbi.nlm.nih.gov/Blast.cgi),通过BLAST比对后,筛选并下载与目的菌株具有较高同源性的序列, 再使用MEGA 7.0软件, 通过邻接法(neighborjoining method, NJ),运行1 000次bootstrap验证, 构建系统发育树[31]。
1.6菌株對草地贪夜蛾成虫的侵染情况
利用无菌解剖刀刮取SHL2020菌株分生孢子,以灭菌0.05%吐温 80 (北京索来宝生物有限公司)溶液配制1×108个/mL 的孢子悬浮液,以灭菌0.05%吐温 80溶液作为对照。 挑选羽化2日龄草贪夜蛾雌、雄成虫,采用喷雾法进行接种[18],接种后将雌、雄成虫分别放置到自制的保鲜膜养虫笼中(20 cm× 20 cm×20 cm),于(25±1)℃,相对湿度(70±1)%,L∥D=16 h∥8 h的光照培养箱中饲养。 雌成虫和雄成虫每处理10头,3次重复。每天定时观察成虫感染死亡情况及感染虫体的外部特征,用灭菌镊子轻轻触碰接种处理后的成虫,无振翅与活动迹象的则认为该虫已经死亡。 发现有死亡虫体时,将其置于底部垫有灭菌滤纸的灭菌培养皿(d=9 cm)中,盖上皿盖,置于(25±1)℃,相对湿度(70±1)%,L∥D=16 h∥8 h的光照培养箱中培养,每天定时观察虫体表面菌丝或分生孢子生长情况,并镜检分生孢子及菌丝形态。
1.7數据分析
利用Excel对试验数据进行初步整理后,再使用SPSS 20.0软件进行处理分析,统计处理雌、雄成虫的累计死亡率和累计校正死亡率[32],并采用Tukey方法计算不同时间雌雄成虫累计死亡率的显著性差异。
累计死亡率= 死虫数 供试虫数 ×100%;
累计校正死亡率=(处理组死亡率 对照组死亡率)/(1-对照组死亡率)×100%。
2结果与分析
2.1菌株形态特征及分子生物学鉴定
对已分离纯化培养的菌株形态特征进行观察,结果发现培养后第3天时菌落平展,白色绒毛状,菌落边缘整齐,呈放射状生长;接种后第6天,菌落开始产孢,略变为淡黄色,菌落中间略凸起,孢子层长势均匀,菌落背面淡黄色;在SDAY培养基上培养至7 d时,菌落直径22.57 mm(图1e~h)。在光学显微镜下菌丝无色,分生孢子透明、光滑,球形或近球形,直径在2.3~2.9 μm,分生孢子梗着生于营养菌丝,光滑呈穗状(图1i~k)。
PCR扩增目的菌株SHL2020的ITS 序列,测序结果显示该菌株扩增片段为542 bp,将该序列在GenBank数据库中进行BLAST比对后,发现SHL2020菌株的ITS序列与已报道的多个球孢白僵菌菌株对应序列的相似性均达到97%以上。从中选取相关序列,使用MEGA 7.0构建系统发育树(图2)。发现SHL2020菌株与 Beauveria bassiana BbH51(MK952353.1)处于一个分支,亲缘关系最为接近,同源性最高。结合形态学和ITS序列相似性分析,最终确定该菌株为球孢白僵菌,菌株编号SHL2020。
2.2室内致病性测定
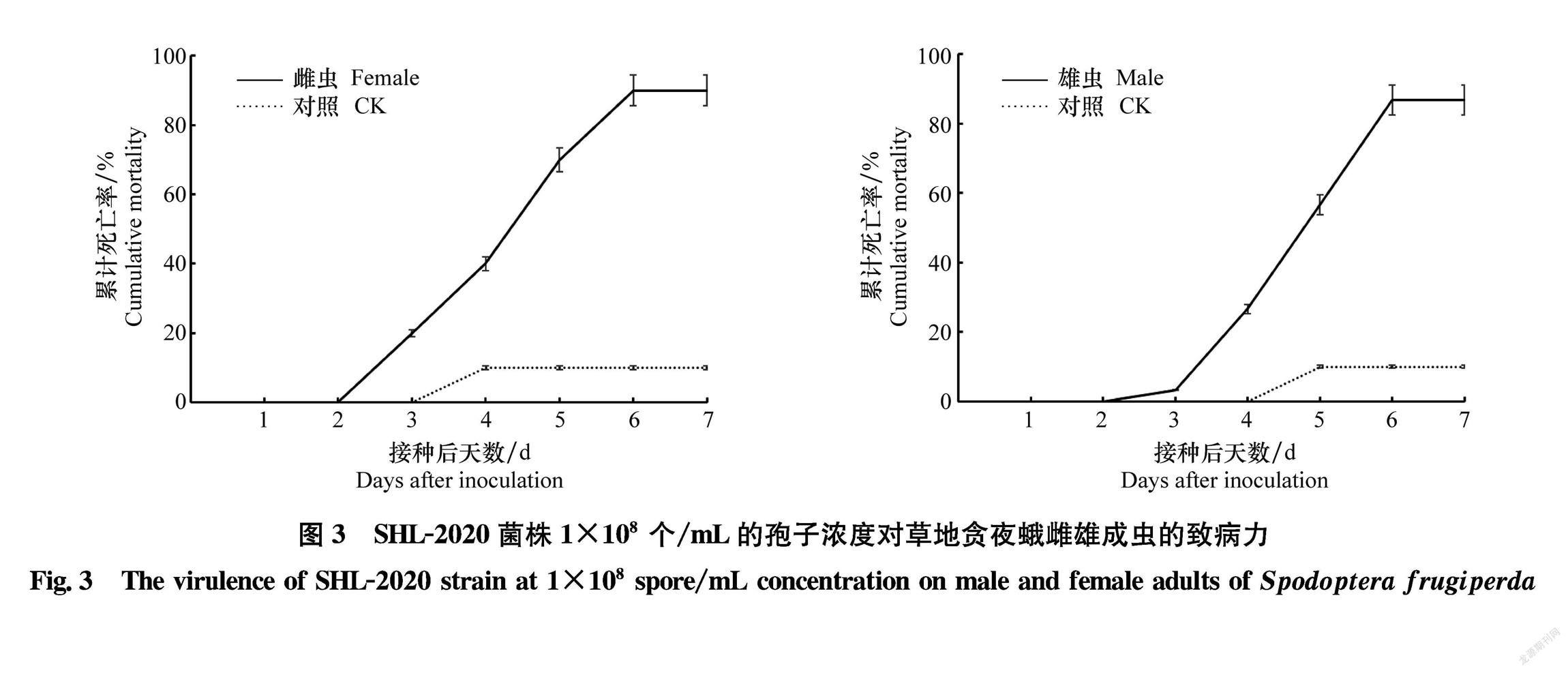
致病性测定结果表明,草地贪夜蛾雌、雄成虫都可被分离培养的SHL2020菌株感染(图1a~d)。用1×108个/mL的孢子悬浮液接种,处理后第3天供试雌、雄成虫开始出现死亡。接菌后第3天,草地贪夜蛾雌成虫累计死亡率为(18.8±2.3)%,显著高于雄成虫的(2.6±2.0)%;接菌后第6天,雌成虫累计死亡率为(89.6±0.5)%,略高于雄成虫(85.8±0.9)%(图3)。通过比较处理后不同时间雌雄成虫累计死亡率,发现除第3天雌雄成虫之间累计死亡率存在极显著差异(F=9.80,P<0.01)外,其余时间,雌雄成虫累计死亡率都无显著差异(表1)。
接种后第3天,供试成虫开始出现死亡。感染后菌丝最先从死亡成虫的头部长出;在(25±1)℃,相对湿度(70±1)%,L∥D=16 h∥8 h的光照培养箱中保湿培养2 d后,死亡虫体的胸部和足部开始有菌丝长出,第5天时成虫胸部和腹部整个长满菌丝,而翅部仅在翅脉有少量菌丝长出。
3讨论
虫生真菌生长繁殖快、易生产,能侵染多种昆虫,且害虫不易产生抗性,在防治农林害虫方面具有明显优势[33]。当前国内外关于球孢白僵菌防治草地贪夜蛾已有许多相关的研究报道[18,20 21,29, 34 -36],但目前已有的研究中多为球孢白僵菌感染草地贪夜蛾卵或者幼虫,有关该菌侵染成虫的研究还鲜有报道。本研究从罹病草地贪夜蛾成虫中分离获得1株病原真菌SHL2020,通过形态学以及ITS序列比对分析,将该菌株鉴定为球孢白僵菌。通过初步毒性测定,发现该菌株与彭国雄等[21]所用的JU435菌株相比,对草地贪夜蛾成虫具有更强的侵染致病作用。SHL2020的发现,不仅丰富了云南本地草地贪夜蛾病原真菌资源,也为草地贪夜蛾生物防治提供了生防潜力菌株。
自2019年1月草地贪夜蛾入侵我国以来,在云南已发现侵染其幼虫且流行性较强的昆虫病原真菌莱氏绿僵菌[18],但未发现有自然感染草地贪夜蛾幼虫的球孢白僵菌。本研究发现了对成虫有较强侵染作用的球孢白僵菌,其是否对草地贪夜蛾幼虫有侵染致病作用尚需进一步研究。此外,虫生真菌的发生和对害虫的侵染受湿度和温度等环境条件的影响[18],本研究发现的球孢白僵菌SHL2020在室内条件下对草地贪夜蛾雌、雄成虫均具有较强致病力,但其在田间自然条件下对草地贪夜蛾成虫的侵染致病作用,还有待研究。
参考文献
[1]郭井菲, 赵建周, 何康来, 等. 警惕危险性害虫草地贪夜蛾入侵中国[J]. 植物保护, 2018, 44(6): 1- 10.
[2]EARLY R, GONZLEZMORENO P, MURPHY S T, et al. Forecasting the global extent of invasion of the cereal pest Spodoptera frugiperda, the fall armyworm [J]. NeoBiota, 2018, 40: 25- 50.
[3]王磊, 陈科伟, 钟国华, 等. 重大入侵害虫草地贪夜蛾发生危害、防控研究进展及防控策略探讨[J]. 环境昆虫学报, 2019, 41(3): 479 487.
[4]MONTEZANO D G, SPECHT A, SOSAGMEZ D R, et al. Host plants of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas [J]. African Entomology, 2018, 26(2): 286 -301.
[5]GUI Furong, LAN Tianming, ZHAO Yue, et al. Genomic and transcriptomic analysis unveils population evolution and development of pesticide resistance in fall armyworm Spodoptera frugiperda [J/OL]. Protein & Cell, 2020, DOI: 10.1007/s13238-020-00795-7.
[6]姜玉英, 劉杰, 谢茂昌, 等. 2019年我国草地贪夜蛾扩散为害规律观测[J]. 植物保护, 2019, 45(6): 10 -19.
[7]杨学礼, 刘永昌, 罗茗钟, 等. 云南省江城县首次发现迁入我国西南地区的草地贪夜蛾[J]. 云南农业, 2019 (1): 72.
[8]魏向敏, 崔勇, 叶国浚, 等. 草地贪夜蛾寄主适应性, 种群动态特征及防控新思路展望[J]. 环境昆虫学报, 2020, 42(1): 42 -51.
[9]梁沛, 谷少华, 张雷, 等. 我国草地贪夜蛾的生物学、生态学和防治研究概况与展望[J]. 昆虫学报, 2020, 63(5): 624- 638.
[10] 吴益东, 沈慧雯, 张正, 等. 草地贪夜蛾抗药性概况及其治理对策[J]. 应用昆虫学报, 2019, 56(4): 599 -604.
[11] CARVALHO R A, OMOTO C, FIELD L M, et al. Investigating the molecular mechanisms of organophosphate and pyrethroid resistance in the fall armyworm Spodoptera frugiperda [J/OL]. PLoS ONE, 2013, 8(4): e62268. DOI:10.1371/journal.pone.0062268.
[12] 蒋骏, 张熠玚, 王文文, 等. 龟纹瓢虫对草地贪夜蛾卵和低龄幼虫的捕食作用[J]. 植物保护, 2020, 46(3): 188- 193.
[13] 范悦莉, 谷星慧, 冼继东, 等. 叉角厉蝽对草地贪夜蛾的捕食功能反应[J]. 环境昆虫学报, 2019, 41(6): 1175 -1180.
[14] 孔琳, 李玉艳, 王孟卿, 等. 多异瓢虫和异色瓢虫对草地贪夜蛾低龄幼虫的捕食能力评价[J]. 中国生物防治学报, 2019, 35(5): 709 -714.
[15] 唐继洪, 吕宝乾, 卢辉, 等. 海南草地贪夜蛾寄生蜂调查与基础生物学观察[J]. 热带作物学报, 2020, 41(6): 1189 -1195.
[16] 王燕, 王孟卿, 张红梅, 等. 益蝽成虫对草地贪夜蛾不同龄期幼虫的捕食能力[J]. 中国生物防治学报, 2019, 35(5): 691- 697.
[17] 赵英杰, 符成悦, 徐天梅, 等. 黄足肥螋成虫对草地贪夜蛾2龄幼虫的捕食功能反应[J]. 植物保护, 2019, 45(6): 35- 38.
[18] 郑亚强, 陈斌, 李正跃, 等. 草地贪夜蛾莱氏绿僵菌的分离鉴定[J]. 植物保护, 2019, 45(5): 65 -70.
[19] 魏雪菲, 郑亚强, 陈斌, 等. 昆明地区草地贪夜蛾莱氏绿僵菌研究初报[J]. 云南农业科技, 2019 (S1): 31- 34.
[20] AKUTSE K S, KIMEMIA J W, EKESI S, et al. Ovicidal effects of entomopathogenic fungal isolates on the invasive fall armyworm Spodoptera frugiperda (Lepidoptera: Noctuidae) [J]. Journal of Applied Entomology, 2019, 143(6): 626- 634.
[21] 彭国雄, 张淑玲, 夏玉先. 杀虫真菌对草地贪夜蛾不同虫态的室内活性[J]. 中国生物防治学报, 2019, 35(5): 729 -734.
[22] 雷妍圆, 吕利华, 王裕华, 等. 一株玫烟色虫草对草地贪夜蛾的致病性研究[J]. 环境昆虫学报, 2020, 42(1): 68- 75.
[23] GROGAN G J, HOLLAND H L. The biocatalytic reactions of Beauveria spp. [J]. Journal of Molecular Catalysis B Enzymatic, 2000, 9(1 3): 1-32.
[24] 赵东容. 白僵菌在农林害虫防治中的应用[J]. 湖北植保, 2018 (1): 53-55.
[25] 朱晓敏, 张强, 李锋, 等. 球孢白僵菌与苦参碱联合作用对稻水象甲成虫毒力测定[J]. 东北农业科学, 2020, 45(2): 41- 43.
[26] 王义勋, 王星冉, 陈京元, 等. 马尾松毛虫高毒力白僵菌菌株筛选[J].南方农业学报, 2016, 47(5): 662- 666.
[27] 吴志鹏, 童应华. 球孢白僵菌和金龟子绿僵菌对红火蚁工蚁的致病力测定[J]. 森林与环境学报, 2020, 40(1): 99 -105.
[28] 蒲蛰龙, 李增智. 昆虫真菌学[M]. 合肥: 安徽科学技术出版社, 1996: 553- 554.
[29] 雷妍圆, 章玉苹, 薛志洪, 等. 一株球孢白僵菌的分离鉴定及其对草地贪夜蛾的致病性[J]. 环境昆虫学报, 2020, 42(3): 593 -601.
[30] 李焕宇, 付婷婷, 张云, 等. 5种方法提取真菌基因组DNA作为PCR模板效果的比较[J]. 中国农学通报, 2017, 33(16): 28 -35.
[31] KUMAR S, TAMTRA K, NEI M. MEGA3: Integrated software for molecular evolutionary genetics analysis and sequence alignment [J]. Briefings in Bioinformatics, 2004, 5(2): 150- 163.
[32] 童应华, 马淑娟, 李万里, 等. 白僵菌与绿僵菌对甜菜夜蛾幼虫的致病力[J]. 福建农林大学学报(自然科学版), 2014, 43(4): 350 -354.
[33] 代园凤, 张承, 喻会平, 等. 一株寄生曲霉SF2对甘蔗绵蚜的感染作用及其生物学特性初步研究[J]. 云南农业大学学报:自然科学版, 2017, 32(6): 1006 -1011.
[34] CARNEIRO A A, GOMES E A, GUIMARES C T, et al. Molecular characterization and pathogenicity of isolates of Beauveria spp. to fall armyworm [J]. Pesquisa Agropecuaria Brasileira, 2008, 43(4): 513- 520.
[35] GARCIA G C, GONZALEZ M B, BAUTISTA M N. Pathogenicity of isolates of entomopathogenic fungi against Spodoptera frugiperda (Lepidoptera: Noctuidae) and Epilachna varivestis (Coleoptera: Coccinellidae) [J]. Revista Colombiana de Entomolog, 2011, 37(2): 217- 222.
[36] THOMAZONI D, FORMENTINI M A, ALVESL F A. Patogenicidadedeisoladosde fungos entomopatogênicos a Spodoptera frugiperda (Smith) (Lepidoptera: Noctuidae) [J]. Arquivos do Instituto Biológico, 2014, 81(2): 126- 133.
收稿日期:2020 -11- 04修訂日期:2021- 01 -07
基金项目:
云南省重点领域科技计划(202001BB050002); 国家重点研发计划(2019YFD0300105)
* 通信作者
Email:李正跃:kmlizhengyue@163.com;陈斌:chbins@163. com
