端粒酶活性检测研究进展
2022-01-09娄筱叮
郭 玥,吴 霞,夏 帆,娄筱叮
(中国地质大学(武汉)生物地质与环境地质国家重点实验室,材料与化学学院,纳米矿物材料及应用教育部工程研究中心,湖北 武汉 430078)
端粒是存在于真核生物染色体末端的特殊结构,由短而多且重复的非转录序列(TTAGGG)及系列结合蛋白组成,对细胞的复制、衰老和凋亡起着重要作用。在人体的体细胞中,随着细胞分裂,端粒DNA 将缩短50~200 个核苷酸,使得细胞逐渐老化并丧失增殖能力而死亡。而端粒酶是一种核糖核蛋白酶,能以自身RNA 为模板,催化合成单链TTAGGG 重复序列于端粒的3'端,用于维持端粒长度。在正常的体细胞中,端粒酶的活性受到抑制。然而在人类肿瘤细胞中,约有85%的端粒酶活性出现上调[1]。因此,端粒酶活性的高表达与癌细胞的永生性密切相关,这使得越来越多的科学家将端粒酶作为一种新的肿瘤标志物、抗癌药物靶点、特异性肿瘤检测标志物诊断和预后指标的重要指标。如何有效地对端粒酶活性进行精准检测及临床应用成为是医学临床诊断和了解疾病机制的重要问题之一。
1 端粒酶活性检测方法
自1985年端粒酶被发现以来,其活性检测方法的研究取得了一些进展。其中,最经典的端粒酶活性检测方法是Piatyszek 等[2]于1995 年提出的依赖聚合酶链式反应(PCR)的端粒重复扩增方法(TRAP)。以后的很多关于端粒酶活性的研究方法均基于PCR 技术提出,使得该方法在灵敏度和特异性方面取得了较大的提高,但其操作繁琐且耗时,易受到诸多因素影响,导致假阴性和假阳性结果,限制了对端粒酶活性的精确量化。为了进一步优化端粒酶的活性检测,近年来,很多新的端粒酶活性检测策略如化学反应发光法、电化学法、荧光分析法、比色法等相继被开发。
1.1 化学反应发光法
化学反应发光是指由化学反应产生的能量而发出的光。化学反应发光法的分析速度快、线性范围宽且灵敏度较高。其中,过氧化物酶催化的底物过氧化氢的化学发光反应被广泛运用于端粒酶活性检测。如Zhu等[3]用鲁米诺/过氧化氢/对碘苯酚溶液代替市售荧光素酶,将端粒酶扩增的特异性与化学发光的高灵敏度相结合,开发出端粒酶活性检测方法(图1)。其机理是,端粒酶引物通过生物素-亲和素相互作用被加载到磁珠(MBs)表面,在端粒酶的催化作用下,TTAGGG 重复序列被逐步添加到引物的3'端,形成一个长的端粒DNA延伸产物,并与多个辣根过氧化物酶标记的互补DNA杂交使得辣根过氧化物酶在磁珠表面富集(多余的辣根过氧化物酶经磁分离和洗涤被洗除)。当磁珠与上述溶液混合时,磁珠上富集的辣根过氧化物酶将催化过氧化氢氧化鲁米诺的反应而发出蓝光(对碘苯酚作为增强剂),发光强度与10~1 000 个细胞范围内HeLa 细胞数的对数呈线性关系,可实现对端粒酶活性的超灵敏检测(检出限为9 个细胞),且该方法通过手持光度计可实现可视化检测,适用于疾病标志物的现场检测。
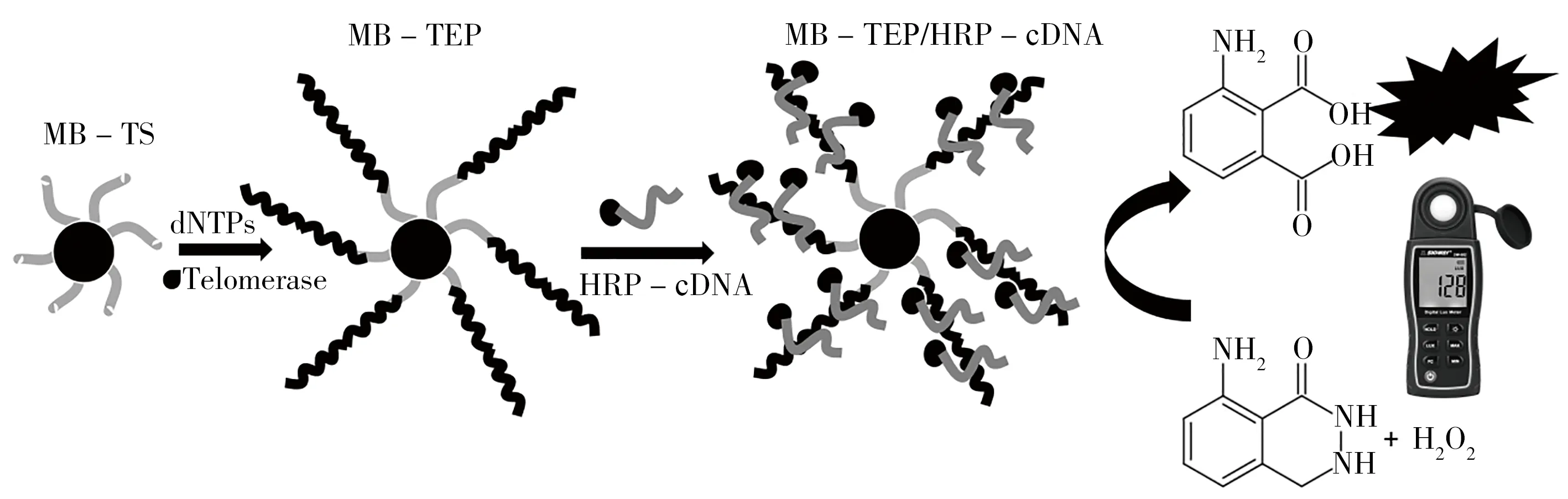
图1 基于辣根过氧化物酶催化的鲁米诺/过氧化氢氧化还原反应与手持式光度计相结合的端粒酶活性现场检测方法[3]Fig.1 Field detection of telomerase activity based on the redox reaction of luminol/hydrogen peroxide catalyzed by horseradish peroxidase and hand-held photometer[3]
1.2 电化学法
1.2.1 单信号电化学法电化学法作为一种快速发展的生物分析技术,具有简单、灵敏度高、便携、微型化的特点。常见的检测端粒酶活性的电化学方法是通过纳米粒子与端粒酶延伸产物作用,锚定电极表面并催化电化学反应引起电化学信号的改变来反映端粒酶活性,常以单一电信号形式读出。铂和金等贵金属纳米粒子具有大的比表面积,三维的大孔碳(MPC)则具有独特的大孔结构、优异的导电性、较大的孔体积和比表面积、良好的化学惰性和热稳定性,另外,环氧官能化的MPC 的表面还有许多-COOH 位点,可与DNA 进行自组装。MPC 与均匀分散的贵金属纳米粒子之间的协同作用使其具有显著的电导率和电催化性能。基于MPC 和铂纳米粒子(PtNPs)的电催化性能,Jia 等[4]设计了一种端粒酶触发催化发夹型DNA 自组装反应(CHA)的电化学检测方法(图2A)。PtNPs 与MPC 结合形成的Pt/MPCCOOH 与发夹DNA 组装成Pt/MPC-COOH-H2探针,在端粒酶的作用下,使大量Pt/MPC-COOH-H2富集于GCE 表面,催化乙酰氨基酚氧化反应,产生明显的电流信号。由于综合了探针优异的电催化性能和线性放大型CHA,该策略具有低的检出限(9.02 个宫颈癌细胞/mL)和宽的线性范围(102~107个宫颈癌细胞/mL)。类似的,酸功能化的有序介孔碳(OMC)表面也具有丰富的-COOH 位点,Wang 等[5-6]基于金纳米棒(AuNRs)的电催化性能,在端粒酶作用下,金纳米棒催化电极表面的电化学反应,根据电化学信号来反映端粒酶活性。
近年来,金属有机骨架(MOFs)的高比表面积和孔隙率、可调节的孔径和形状及良好的生物相容性使其在电化学生物传感中具有极大的潜力。将PtNPs 镶嵌在MOFs 材料表面,能阻碍PtNPs 的自聚集,提高其稳定性,获得显著的电催化性能。Ling 等[7]开发了基于cDNA@Pt@P-MOF(Fe)催化的电化学检测方法(图2B)。在端粒酶作用下,将展开大量发卡DNA(HpDNA)与cDNA@Pt@P-MOF(Fe)结合,从而催化过氧化氢分解,使电信号增强。端粒酶活性与吸附的电催化剂的量成正比,利用EXO III酶切信号循环放大作用可使检出限更低。类似的,Wang等[8]通过设计催化对乙酰氨基酚氧化的电化学生物传感方法,实现了端粒酶和其它分析物的逐步检测。Yang等[9]开发了基于MOFs材料的Zn2+和端粒酶的串联逐步识别策略。
除了利用纳米粒子对电化学反应的催化性能外,还可以通过其它电化学手段来表征端粒酶活性。比如,Liu 等[10]利用端粒酶延伸产物与链霉亲和素(SA)共同存在时,引发SA-生物素-DNA-生物素原位绝缘层的产生,通过电化学阻抗谱和扫描电化学显微镜共同监测电极表面性质的变化,实现了对端粒酶活性的灵敏检测。
1.2.2 双信号电化学法双信号比率输出克服了单个信号输出时受到的信号、背景波动的影响,检测的准确性更高。比如,Dong等[11]设计了一种亚甲蓝修饰的发夹探针,建立了用于端粒酶活性检测的双信号比率输出方法(图2C)。在端粒酶作用下,发卡探针打开,亚甲蓝远离电极表面,原电流信号降低,Au@CeMOF-cDNA 随之锚定于电极表面并催化对苯二酚(HQ)氧化为对苯醌(BQ)的反应,使得另一电流信号增加。由于双信号比率输出,该策略具有宽的线性范围和低的检出限,可用于检测单个细胞的端粒酶活性。
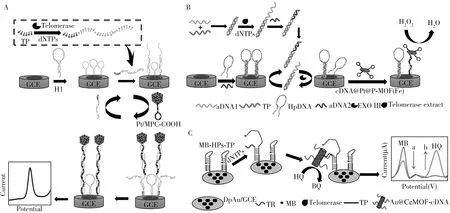
图2 一种基于Pt/MPC-COOH的端粒酶触发催化发夹DNA自组装反应(CHA)的电化学传感器(A)[4];利用cDNA@Pt@P-MOF(Fe)电催化性能检测端粒酶活性的策略(B)[7];利用AuNPs锚定MOFs后的电催化性能检测端粒酶活性的电化学生物传感器(C)[11]Fig.2 An electrochemical biosensor based on the integration of the telomerase-initiated catalyzed hairpin assembly(CHA)process with electrocatalysis by Pt/MPC-COOH(A)[4];a strategy for detecting telomerase activity using cDNA@Pt@p-MOF(Fe)electrocatalytic performance(B)[7];an electrochemical biosensor for detecting telomerase activity using the electrocatalytic performance of MOFs anchored by AuNPs(C)[11]
1.3 荧光分析法
1.3.1 传统荧光染料传统的荧光分子及其衍生物以其优异的光物理性、热稳定性、易修饰等特性被广泛用于荧光传感材料、自组装材料和纳米材料。若一个荧光基团(供体)的发射光谱和另一基团(受体)的吸收光谱之间存在重叠,当供体和受体之间的距离缩短时,荧光能量将由供体向受体转移,该现象称为荧光共振能量转移(FRET)。荧光染料和氧化石墨烯、AuNPs之间可以产生荧光共振能量转移现象。基于氧化石墨烯与单链DNA 之间的荧光共振能量转移现象,Ou 等[12]建立了一个端粒酶和miRNA-21(miR-21)同时检测的传感平台(图3A)。由于π-π 堆积相互作用,端粒酶引物和端粒酶探针、miR-21 探针可被加载到氧化石墨烯上产生荧光共振能量转移现象导致荧光猝灭,在端粒酶催化下将延伸端粒酶引物并使之与端粒酶探针互补配对,从而远离氧化石墨烯使异硫氰酸荧光素(FITC)发出荧光。由于氧化石墨烯的生物相容性好,该平台可用于同时准确检测活细胞或癌症患者组织样本中的端粒酶活性和miR-21。
三维DNA walker 可通过酶切释放被猝灭的荧光染料,从而使得荧光共振能量转移现象消失,荧光染料离开纳米金表面后发出荧光,Huang 等[13]设计了一种三维DNA walker,在端粒酶的存在下,三维DNA walker 中的引物被拉长,通过酶切割释放Cy5 荧光染料,染料远离金纳米表面而发光。基于荧光染料和AuNPs 之间的荧光共振能量转移现象,Xu 等[14]使用MnO2负载端粒酶引物进入活细胞细胞,实现了活细胞的端粒酶活性检测。Xu等[15]构建了基于金纳米粒子的三维脱氧核糖核酸酶机制,并成功应用于活细胞端粒酶的共聚焦成像识别。Yang等[16]基于量子点(QD)和荧光染料的FRET设计了一种DNA四面体纳米探针检测端粒酶活性(图3B)。该探针的其中一段序列的5'和3'端分别用荧光供体量子点和荧光受体Alexa488染料分子标记,因形成了发夹结构导致供体和受体之间的距离缩短,从而产生FRET效应。当探针进入细胞后,经过端粒酶延伸,替换下发卡结构并使其环结构打开,FRET消失。该纳米探针灵敏度高且检出限为单个细胞,可用于细胞端粒酶活性的动态监测。2019年,Ma等[17]根据2-氨基嘌呤荧光分子在分子信标中因碱基堆积作用而产生的荧光猝灭,构建了一个无猝灭剂的二次信号放大系统,能灵敏准确地检测相当于1 个肺癌细胞的端粒酶,在癌症诊断和抗癌药物发现方面具有巨大的潜力。
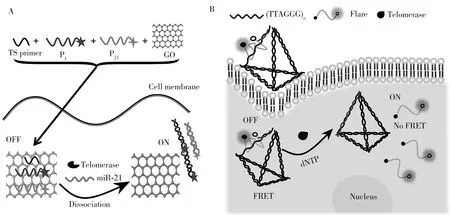
图3 端粒酶活性和miRNA-21同时检测的传感平台(A)[12];基于FRET的三维DNA纳米探针的比率传感器检测端粒酶活性的示意图(B)[16]Fig.3 A sensing platform for simultaneous detection of telomerase activity and miRNA-21(A)[12];schematic of a FRET based three-dimensional DNA nanoprobe ratio sensor for detecting telomerase activity(B)[16]
DNA四面体是一种机械刚性的纳米结构,无需转染试剂即可内化到细胞内,细胞内化效率高、结构稳定性和生物相容性好。Meng等[18]基于一种新型的DNA纳米探针建立了简单的比率传感器,该探针由端粒酶延伸引物及用于自我递送的三维DNA纳米结构构成。端粒酶可改变DNA纳米探针的构型,导致两个标记的荧光团之间的距离增加,FRET效率明显降低,通过计算两个荧光团的峰值强度即可实现端粒酶活性的比率传感,规避了由于单一信号导致的假阳性干扰的问题。该方法可用于在单细胞水平上端粒酶活性的检测。类似的,Zhang等[19]开发了一种按顺序点亮的多色DNA四面体探针。在端粒酶的作用下,探针上的分子信标可依次被打开,并在不同波长处发出荧光,依次点亮多种颜色,实现活细胞内端粒酶活性的检测。除设计受体和供体外,还可直接设计成带有自猝灭的信标,无需额外的猝灭剂。
传统的荧光染料大多基于分子信标检测端粒酶活性,需要同时标记染料基团和猝灭基团,设计复杂,合成繁琐,且具有疏水性,当分子信标在靶标上聚集时,会导致严重的荧光猝灭,即聚集荧光猝灭(ACQ)效应。故传统的荧光染料在溶液状态下的背景噪音大,光稳定性差,仅能在极稀溶液中或单分子状态下发挥作用,大大限制了传统荧光分子在端粒酶活性检测中的运用[20]。
1.3.2 聚集诱导发光荧光探针2001年,Tang等[21]首次提出“聚集诱导发光(AIE)”的概念,并且提出了限制分子内运动等机制。AIE 分子在溶液状态下具有低的背景噪音和高的光稳定性,与传统的ACQ效应截然相反,利用此类分子能大大拓宽荧光探针在生物化学传感领域中的应用[22]。
基于AIE分子的模块化组装探针可以提高细胞内化效率及检测的特异性。Wang等[23]将具有聚集诱导发光特性的四苯基乙烯(TPE)分子和DNA 结合得到两亲性的共轭聚集诱导发射探针(TPE-RDNA),用于检测细胞中MnSOD mRNA 的表达水平,实现了对肿瘤组织的成像和预后分析。Wu等[24]开发了模块化DNA 结合的聚集诱导发光探针(TPE-Py-DNA)用于甲基化转移酶的检测和成像。此外,聚集诱导发光探针还可用于化疗、光动力疗法、基因治疗、手术治疗、复合治疗等复合治疗系统[25-33]。以上结果表明AIE分子在体内、体外检测和诊疗一体化的应用中都具有很大优势。
AIE分子带正电,端粒酶引物带负电,二者可通过静电吸引相互结合。Lou等[34]设计了带正电的四苯基乙烯类AIE 分子(TPE-Z),用于检测膀胱癌患者尿液中的端粒酶活性(图4A)。在端粒酶的作用下,端粒酶引物被延伸后,带正电的TPE-Z分子吸附产生聚集诱导发光现象,荧光强度大大增强。为了进一步减低发射背景,Zhuang 等[35]利用AIE 分子和猝灭剂之间的FRET 效应,以猝灭剂修饰的端粒酶引物为探针(QP),TPE-Py分子作为荧光分子来检测端粒酶活性(图4B)。TPE-Py分子通过静电作用吸附于探针上,由于FRET现象导致荧光猝灭,在端粒酶作用下,使得越来越多的AIE分子吸附于猝灭剂修饰的端粒酶引物延长链上,逐渐远离猝灭基团,荧光信号得以增强。上述方法使用的AIE分子,展现出优异的光稳定性、低的荧光背景信号干扰以及低的生物毒性,非常适合用于细胞原位长效示踪。在上述研究基础上,Zhuang等[36]设计了猝灭剂修饰的端粒酶引物以及两种AIE分子(TPE-H和Silole-R),使得原本灵敏度不够的TPE-H 在Silole-R 的协同作用下,进一步拓宽了检测的范围,并用于活细胞成像(图4C)。此外,基于不同细胞周期的变化过程,Wu 等[37]首次建立了基于聚集诱导发光分子的上下游联动检测系统,基于聚集基元和核酸基元的定量荧光分析法,解析了细胞周期中各个时相端粒酶的逆转录蛋白的mRNA 表达水平(TERT mRNA)和端粒酶活性的动态过程(图4D)。结果说明忽略周期中的异质性对细胞群体进行端粒酶活性检测,将影响结果的准确性。因此,细胞周期同步后分析的端粒酶活性检测技术为肿瘤标志物的精准检测提供了一个新的切入点。
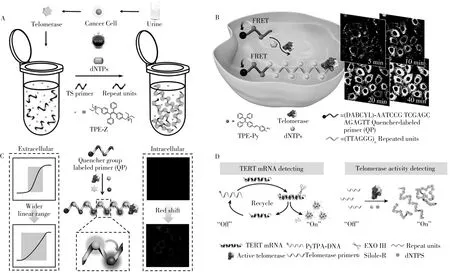
图4 基于聚集诱导发光特性的TPE-Z荧光探针用于端粒酶活性检测(A)[34];基于FRET的AIE荧光探针端粒酶活性原位检测(B)[35];双荧光信号探针用于检测原位或细胞提取物中的端粒酶活性(C)[36];基于聚集诱导发光分子的上下游联动检测系统检测细胞中的端粒酶mRNA表达水平和端粒酶活性(D)[37]Fig.4 Telomerase detection with TPE-Z fluorescent probe based on AIE(A)[34];AIE fluorescent probe used telomerase activity detection based on FRET(in situ)(B)[35];dual fluorescent signal probes were used to detect telomerase activity in situ or in cell extracts(C)[36];telomerase mRNA expression level and telomerase activity in cells were detected by the upstream and downstream linkage detection system based on AIEgens(D)[37]
1.4 端粒酶可视化检测
1.4.1 比色分析法比色分析法是通过直观颜色的变化或测定吸光度来定性或定量分析待测物含量的方法。Wang 等[38]设计了一种端粒酶活性检测的比色分析方法(图5A),将端粒酶引物(TS)固定在磁珠(MBs)上,当端粒酶和脱氧核苷三磷酸(dNTPs)存在时,端粒酶延伸产物与单链DNA 互补配对,通过生物素-链霉亲和素-生物素作用将脂质体包裹的辣根过氧化物酶连接到MBs 表面。辣根过氧化物酶催化过氧化氢和3,3',5,5'-四甲基联苯胺(TMB)之间的氧化还原反应,产生的TMB2+作为氧化剂定量蚀刻金纳米粒子。金纳米粒子具有独特的光学特性,其形态的变化和所诱导的调性转化有望表现出多种鲜艳的颜色变化和肉眼可见的高分辨率。Wang 等[39]开发了一种端粒酶活性可视化比色分析方法(图5B)。端粒酶引物(TS)被固定在磁珠表面,在端粒酶作用下,通过磁珠的洗涤和分离,被延长的端粒DNA 将富集在MBs 上,在K+和氯化血红素存在下形成许多G-四链体,高效地催化过氧化氢氧化TMB,产生蓝色产物。一个细长的TS包含几个重复单元,可以形成多个G-四链体,从而得到有效的比色读出。
1.4.2 联合设备检测联合其他设备可以实现对端粒酶活性的可视化检测,比如基于辣根过氧化物酶催化的过氧化氢发光反应检测化学发光强度,Zhu等[3]通过手持式光度计简单而灵敏地检测了肿瘤生物标志物端粒酶的活性,可应用于疾病生物标志物的现场检测(图1A)。为进一步探索端粒酶活性检测信号放大的方法以不断提高灵敏度,Liu等[40]开发了一种由葡萄糖仪读出的核酸酶切割引发双循环信号放大的端粒酶活性检测平台。利用端粒酶在引物上的扩增及核酸酶特异性切割(由Pb2+激活核酸酶),使蔗糖转化酶富集在MBs表面,能高效催化蔗糖转化为葡萄糖,用葡萄糖仪读出信号(图5C)。该方法通过双循环实现信号放大的同时,还利用金属离子及其激活的酶,提高了检测的特异性和灵敏度,检出限低至5个HeLa细胞。
此外,微芯片电泳化学发光检测在生物大分子分析中具有许多优点,能减少来自基体本身的干扰。Chen 等[41]开发了基于微芯片电泳化学发光以及T7 核酸外切酶依赖的信号放大端粒酶活性检测平台(图5D)。由辣根过氧化物酶(HRP)修饰的探针H1 和端粒酶延伸的产物互补配对后,即被T7 核酸外切酶特异性识别并切割,释放辣根过氧化氢酶和端粒酶延伸产物,释放的端粒酶延伸产物继续和新的HRP修饰的探针H1互补配对,实现信号放大。HRP修饰的探针H1和释放的HRP都能催化过氧化氢的氧化还原反应,可通过微芯片电泳技术实现二者分离,获得较高的分辨率。该方法操作简单、分析时间短、灵敏度高且不易受样品底物的干扰,可用于单细胞的端粒酶活性检测。
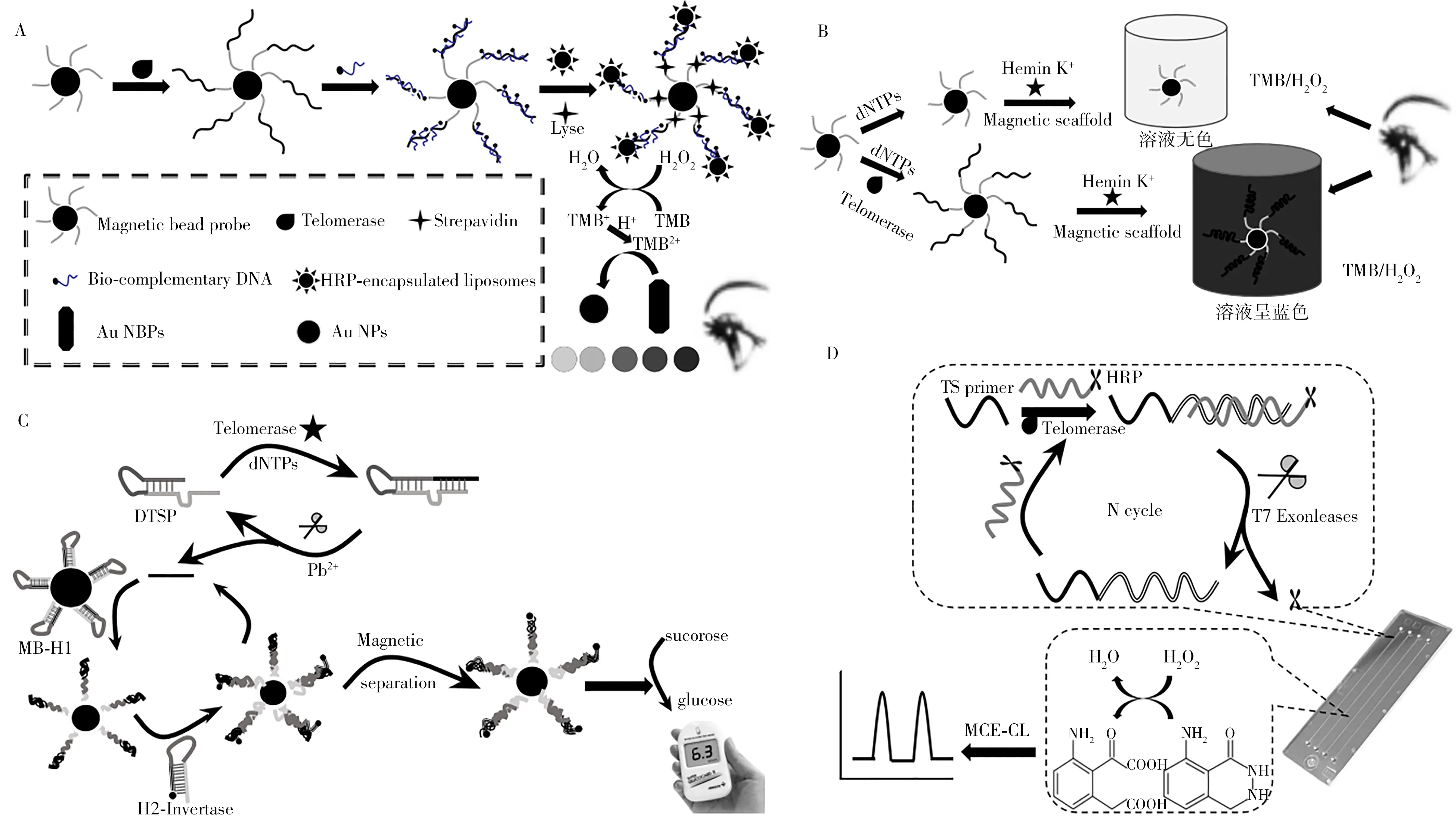
图5 基于辣根过氧化物酶催化过氧化氢和TMB之间的氧化反应产物TMB2+定量蚀刻AuNPs的端粒酶活性检测的可视化比色分析法(A)[38];基于在K+和氯化血红素存在下催化TMB/过氧化氢反应的G-四联体形成的端粒酶活性的可视化比色分析法(B)[39];结合化学反应发光和葡萄糖计检测端粒酶活性的方法(C)[40];基于微芯片电泳化学发光及微芯片电泳技术实现端粒酶活性检测的方法(D)[41]Fig.5 Based on quantitative etching of AuNPs by TMB2+,a visualized colorimetric strategy for telomerase activity detection between hydrogen peroxideand TMB catalyzed by horseradish peroxidase(A)[38];a visualized colorimetric method for detecting telomerase activity based on the formation of DNA G-quadruplex catalyzing the TMB/hydrogen peroxide reaction under the presence of K+and hemin(B)[39];a method combining chemiluminescence and glucose meter for detecting telomerase activity(C)[40];a chemiluminescence and microchip electrophoresis method for telomerase detection(D)[41]
1.5 其它检测方法
除上述几大类端粒酶的高效检测方法外,近年来还发展了一些其他检测方法。如Li等[42]使用杂交链式反应(HCR)和动态光散射(DLS)的方法,开发了通过检测人尿中端粒酶活性来诊断膀胱癌的体外检测方法。在端粒酶的作用下引发AuNPs 聚集,流体动力学直径增加,可用DLS 进行检测。Li 等[43]提出了一种表面增强拉曼散射(SERS)方法进行端粒酶活性检测,可检测单个细胞提取物的端粒酶活性。Ye 等[44]以金纳米粒子为基底,利用SERS 增强和进入癌细胞的自我递送能力,通过两个发夹DNA(H1、H2)的杂交合成了比率型双模式探针,检测了乳腺癌细胞中端粒酶的活性。在端粒酶的作用下,Rox的荧光开启、拉曼关闭,探针中Cy3 的荧光关闭、拉曼开启。与单模成像(荧光技术或SERS 技术)相比,双光谱成像方法可提供丰富的端粒酶动态信息,能更精确地检测活细胞中的端粒酶活性,提高单个活细胞端粒酶活性检测的灵敏度。这也是将具有反向变化的SERS信号比测量首次用于实时活细胞中端粒酶的检测。
2 结论与展望
端粒酶活性检测方法主要可划分为以下几大类:化学反应发光法、电化学检测法、荧光分析法和可视化检测法(比色法、联合设备检测)。在无数科研工作者的努力下,端粒酶活性检测取得了巨大的进步。但在实际应用过程中,这些方法仍然具有一定局限性,比如化学反应发光法中的酶容易降解,易受到环境的干扰,检测成本高。电化学法检测端粒酶活性存在信号探针不稳定、需要添加额外的氧化还原介质等问题。荧光分析法中的信号探针常常需要复杂的合成步骤且生物兼容性差,产生的背景噪声大大降低了检测的灵敏度。比色法的检测现象明显,但灵敏度普遍较低,难以实现原位检测,例如基于G-四链体催化性能的比色法只能实现端粒酶的体外检测。
此外,目前大多数端粒酶活性检测仍为体外检测,导致无法准确了解真实样品活体细胞中端粒酶的活性情况。因此,开发可准确定量、灵敏度高、可操作性好、检测成本低的方法来监测细胞中端粒酶的活性仍然是一个巨大的挑战。随着端粒酶活性检测的不断发展和进步,对活细胞中端粒酶活性的调节以及靶向端粒酶的特异性药物治疗手段也将成为未来几年端粒酶研究领域需要攻克的难题。
