利用CRISPR/Cas9技术创制早熟甘蓝型油菜材料
2022-01-05陈娜娜韦世豪朱彦涛张彦锋穆建新
乔 幸,安 然,陈娜娜,韦世豪,朱彦涛,陈 静,张彦锋*,穆建新*
(1.陕西省杂交油菜研究中心,陕西杨凌 712100;2.绵阳市农业农村局,四川绵阳 621010)
当前,我国食用植物油供不应求、严重依赖进口。甘蓝型油菜作为我国主要的油料作物之一,具有重要的经济价值。开花是植物从营养生长进入生殖生长的重要阶段,油菜经过春化处理才能开花成熟,开花的早晚与早熟性密切相关。开花性状由复杂的多基因控制,拟南芥已经报道的与开花相关的基因有120多个,并且控制开花的调控网络也逐渐明[1]。FLC和FRI是开花途径中2个关键基因,FLC基因编码MADS-box转录因子,是开花时间抑制因子[2]。FRI通过促进FLC的表达可抑制油菜开花[3]。对甘蓝型油菜的5个FLC同源基因(BnFLC1-BnFLC5)在拟南芥中分别进行过量表达,结果显示BnFLC1-BnFLC5都能延迟拟南芥开花,BnFLC1、BnFLC2和BnFLC3抑制开花的程度明显高于BnFLC4和BnFLC5[4]。甘蓝型油菜有4个FRI同源基因,分别是Bna A3.FRI、Bna A10.FRI、Bna C3.FRI和 Bna C9.FRI,其中Bna A3.FRI与N3连锁群上效应最大的QTL共定位,因此Bna A3.FRI是控制甘蓝型油菜开花期的主要因子之一[5]。
CRISPR/Cas9基因编辑技术,能够对靶向基因进行精准敲除、序列替换和定点突变,快速创制目标突变体,已成为作物种质资源创制的重要手段。目前,CRISPR/Cas9系统已在拟南芥、烟草、水稻、高粱和小麦等农作物中成功应用[6-10],在油菜中也已有报道[11-12]。
本研究通过CRISPR/Cas9基因编辑同时敲除甘蓝型油菜BnaFLC和BnaFRI,成功创制了BnaFLC和BnaFRI多基因敲除的油菜早熟突变体,该突变体不需春化,从播种到开花只需59 d,表现出明显极早熟特征。该研究结果为早熟材料的创制探索了一条新的途径,为进一步研究早熟性状调控机理以及油菜早熟育种创制了宝贵的种质材料。
1 材料和方法
1.1 材料
甘蓝型油菜(Brassica napus L.,2n=38,AACC)半冬性品种浙油50(浙江省农科院培育)。
1.2 方法
1.2.1 BnFLC及BnFRI基因敲除靶位点序列设计
利用CRISPR/Cas9靶位点在线设计网站(CRISPR DESINGER),结合 Brassicadatabase(http://brassicadb.org/brad/)和NCBI提供的序列信息,对甘蓝油菜基因BnFLC和BnFRI分别进行比对分析,分别设计FLC和FRI的靶位点。
1.2.2 载体的构建
CRISPR/Cas9基因编辑载体pHSE401由中国农业大学陈其军教授提供,构建方法详见参考文献[13],油菜FLC和FRI基因靶向位点引物序列见表1。

表1 油菜FLC和FRI基因靶向位点引物序列Table 1 Primer sequences for targeting sites of BnFRI and BnFLC genes in Brassica napus
1.2.3 甘蓝型油菜的遗传转化
浙油50种子用75%乙醇消毒1 min,加入15%次氯酸钠(200 mL次氯酸钠加入吐温 200 μL),消毒15 min。ddH2O漂洗4~5次,点播于1/2 MS培养基上,25℃暗培养5~7 d。切取8~10 mm左右的下胚轴作为外植体,用含有重组质粒的农杆菌菌液(OD=0.4~0.6)浸染外植体15 min,在M1培养基上共培养2 d,转入潮霉素(浓度50 mg/mL)M2培养基中继续培养20 d后,再转入潮霉素(浓度50 mg/mL)M3培养基中继续培养,切取幼芽,转入M4生根培养基,生根后移入基质培养,炼苗后转至气候室继续培养,收获种子。
1.2.4 阳性植株靶位点检测
选取具有明显早花表型的基因编辑植株提取DNA(SDS法),用载体序列特异性扩增引物U6-26p-F、CRIS-JD-R-all(表2)进行遗传转化真实性检测。
1.2.5 CRISPR/Cas9敲除位点检测
分别设计BnaFLC和BnaFRI靶位点侧翼特异引物(表2),PCR扩增含有靶位点的目标序列,构建克隆载体,测序,解析早花材料BnaFLC和BnaFRI基因的突变位点。

表2 CRISPR/Cas9敲除位点检测引物Table 2 Detection primers of CRISPR/Cas9 knockout sites
1.2.6 T1代植株靶位点检测
将基因敲除后的早熟表型植株套袋自交,提取子代明显早花植株的DNA,用载体序列特异性扩增引物U6-26p-F、CRIS-JD-R-all筛选不含转基因成分的植株,并用含有靶位点的目标序列引物(表2),检测靶位点突变序列,以确定具有非转基因特性的早熟性状的突变材料,用于后续早熟油菜品种的培育。
2 结果与分析
2.1 靶位点设计及载体的构建
2.1.1 靶位点设计
对比已公布的甘蓝油菜基因BnFLC和BnFRI基因序列,利用在线设计软件设计,选择BnFLC基因上3个靶点,BnFRI基因上4个靶点。
BnFLC基因在甘蓝型油菜中有9个拷贝,分别位于 A2(BnaA02g00370D)、A3(BnaA03g02820D、BnaA03g13630D)、A10(BnaA10g22080D)、C2(BnaC0-2g00490D)、C3(BnaC03g04170D、BnaC03g16530D)和 C9(BnaC09g46500D、BnaC09g46540D)染色体上。设计的靶位点T1位于BnFLC基因A2或C2染色体的第6个外显子区域,可以同时编辑A2和C2染色体的2个同源拷贝。靶位点T2位于BnFLC基因A10或C9染色体的第1个外显子区域,可以同时编辑A10和C9染色体的3个同源拷贝。靶位点T3位于BnFLC基因A10或C9染色体的第3个外显子区域,可以同时编辑A10和C9染色体的3个同源拷贝(图1)。
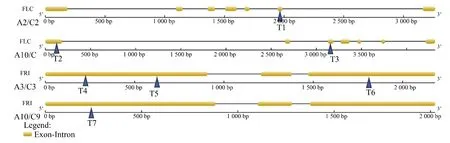
图1 甘蓝型油菜BnFLC和BnFRI基因敲除位置信息图Figure 1 BnFLC and BnFRI knockout location
BnFRI基因在甘蓝型油菜中有4个同源基因,分别位于 A3(BnaA03g13320D)、A10(BnaA10g058-50D)、C3(BnaC03g16130D)和 C9(BnaC09g27290D)染色体上。设计的靶位点T4和T5位于BnFRI基因A3或C3染色体的第1个外显子区域,可以同时编辑A3和C3染色体的2个同源拷贝。靶位点T6位于BnFRI基因A3或C3染色体的第3个外显子区域,可以同时编辑A3和C3染色体的2个同源拷贝。靶位点T7位于BnFRI基因A10或C9染色体的第1个外显子区域,可以同时编辑A10和C9染色体的2个同源拷贝(图1)。
靶位点T6、T7(表1)为BnFRI基因的单靶点,为提高靶点效率,设计5个双靶位点,可以同时敲除BnFRI和 BnFLC 2 个基因,分别是 T2+T5、T1+T4、T1+T6、T3+T4和T1+T5(表1),具体靶位点序列见表 1。
2.2 油菜阳性植株靶位点检测
使用农杆菌介导法侵染甘蓝型油菜下胚轴,获得一株具有明显早熟表型的植株GM13,该阳性植株比正常植株(浙油50)提早抽薹,提早开花40 d左右。利用靶点检测引物U6-26p-F、CRIS-JD-R-all(表2)进行PCR检测,获得500 bp的单靶点阳性片段以及1 000 bp的双靶点阳性片段,验证早熟植株GM13被成功转化。阳性片段克隆测序,对比靶位点序列。验证该植株靶位点T1-T7(表1)均被成功转化。
2.3 阳性转化苗靶位点编辑结果检测
扩增阳性植株靶位点所在区域片段,并对PCR产物进行克隆测序分析,发现所有靶位点T1-T7中,BnaFLC基因中存在2种编辑情况,即BnaFLCA2(BnaA02g00370D)染色体的靶位点T1存在一个碱基C的插入;C2染色体缺失7个碱基(AAGAGAA);BnaFRI基因中存在1种编辑情况,即C9染色体的T7靶位点有一个碱基T的插入,其余靶位点均未被编辑(表3)。

表3 CRISPR/Cas9敲除结果Table 3 Result of CRISPR/Cas9 knockout
2.4 T1代植株的突变是否可稳定遗传
早熟表型植株GM13的T1代植株与对照浙油50相比(图2),有明显早熟性状,表现为提早抽薹开花。选取3株T1代植株提取植物DNA为扩增模板,首先用载体序列特异性扩增引物U6-26p-F、CRIS-JD-R-all筛选不含转基因成分的植株,再对每个T1代株系PCR扩增后送2个克隆测序,结果表明T1代植株的编辑位点序列与T0代植株的靶位点编辑情况完全一致(表4)。表明突变位点可稳定遗传到下一代。

图2 基因编辑早熟材料T1代植株与ZY50开花时间比较Figure 2 Flowering time of T1 generation plants compared to ZY50

表4 T1代植株突变位点情况Table 4 Result of CRISPR/Cas9 knockout in T1 generation plants
3 讨论
CRISPR/Cas9技术基因编辑可以对基因组进行定点修饰和定点突变,现已被广泛应用于拟南芥、水稻、玉米和烟草中,目前在油菜中已有报道利用该技术创制高油酸甘蓝型油菜新种质[14]、创建甘蓝型油菜多室突变体[15]。甘蓝型油菜为异源多倍体植物,基因组高度重复,基因存在多个拷贝,但拷贝之间又不完全相同,基因定向编辑难度较二倍体植物高,虽然转基因阳性率可以高达92.5%,但多靶点CRISPR/Cas9的T0代突变效率为12.9%~59.5%,单靶点平均编辑效率为6.5%~14.9%[15]。高谢旺等[14]对两个高油酸基因BnaFAD2和BnaFAE1进行基因编辑,T0代均未发生基因编辑,在T1代检测到了编辑的发生。本研究设计的7个靶位点均被成功转化,但T0代被成功编辑的靶位点仅有2个。说明基因编辑在多倍体植物中效率不高,如果采取单个靶点遗传转化,鉴定工作量大,费时费力。为提高基因转化效率和编辑效率,获得基因编辑的早花甘蓝型油菜材料,我们在遗传转化中将含有多个靶位点的载体混合遗传转化,转化后获得表型植株再检测验证植株靶位点的编辑情况,通过表型筛选植株减少了直接鉴定阳性植株的盲目性并且更加可靠。
FLC和FRI是油菜春化途径的两个关键基因,甘蓝型油菜中的9个FLC同源基因中,BnaFLC.-A02、BnaFLC.C02、BnaFLC.A10 与油菜早晚熟开花有密切关系。代书桃[16]将BnFLC.A10基因转入春油菜品种Westar,发现在春油菜环境或者无春化处理条件下阳性株系花期延迟。魏大勇[17]对甘蓝型油菜开花期进行了GWAS分析,检测到位于A02染色体上的已知开花基因BnaA02g00370D在冬性油菜和春性油菜中表达量差异达到极显著水平。陈磊[18]通过转基因验证试验证实BnFLC.A2导致双亲材料开花期差异,BnFLC.C2导致晚花亲本R15和早花亲本R11开花期差异,因此BnFLC.A2和BnFLC.C2基因在花期调控中起着重要的作用。本研究中FLC基因位于染色体A10和C9的同源基因并未编辑成功,而A2、C2染色体上的同源基因均被成功编辑,推测相较BnaFLC.A10和BnaFLC.C9,BnaFLC.-A02、BnaFLC.C02是影响开花时间的主要因子。
FRI是FLC上游调控因子,促进FLC基因的表达以此来抑制开花,甘蓝型油菜中分离的4个FRI同源基因中[5],BnaA3.FRI可能是决定油菜春化的主要因子,其余3个基因与甘蓝型油菜春化以及开花的关系尚需进一步研究[19]。本研究中,FRI基因C9的碱基T插入,导致氨基酸发生变化及基因功能突变,但该反应对开花的影响有待于进一步剖析。T1代植株1中,虽未检测到BnFRI基因靶位点被编辑,但植株具有明显的早熟性状,相比对照浙油50早抽薹,早开花。这可能是由于T1代的克隆测序程度不够,或BnFLC基因靶位点被编辑后对早熟性状的贡献更大导致的。
