黑木耳多糖对高脂饮食大鼠肠道菌群的影响
2021-11-17白云飞王雅楠刘昕桐霍宁宁时若寒李研东谭力铭李冲杨新韩雪
白云飞,王雅楠,刘昕桐,霍宁宁,时若寒,李研东,谭力铭,李冲,杨新,韩雪,3*
(1.河北科技大学生物科学与工程学院,河北 石家庄 050018;2.河北省兽药监察所,河北 石家庄 050000;3.河北省功能食品技术创新中心,河北 石家庄 050000)
黑木耳(Auricularia auricula)作为一种药食同源资源,具有丰富的营养和药用价值[1]。黑木耳多糖(Auricularia auricula polysaccharides,AAP)是黑木耳中重要的活性成分[2],作为天然活性多糖,已被证明具有广泛的生理调节功能,目前研究热点主要集中在黑木耳多糖的抗氧化[3]、抗肿瘤[4-6]、免疫调节[7]及益生作用[8]等方面。
针对黑木耳多糖益生作用的研究,通常以益生菌数量增殖效果为评价指标,而益生菌代谢产物或次级代谢产物对人体健康具有重要作用[9]。黑木耳多糖的肠道益生作用研究通常仅针对正常生理实验动物的肠道菌群种类、数量等指标进行分析[10],未充分考虑特殊生理人群和病理人群的特点。高血脂患者肠道菌群紊乱[11],黑木耳多糖及其复配物是否具有针对该类人群的调节作用有待探究。本研究在提取并鉴定黑木耳多糖的基础上,评价其对乳酸菌蛋白质类代谢产物的影响及其对高血脂人群的肠道益生作用,以期为未来高血脂人群功能性食品开发奠定理论基础。
1 材料与方法
1.1 材料、试剂与实验动物
乳酸链球菌菌株(HW-1):河北科技大学营养与功能性食品研究室保留菌种;黑木耳:中康蔬菜种植有限公司;银耳多糖、山楂黄酮:河北科技大学营养与功能性食品研究室提供,乳酸菌培养用MRS液体或固体培养基[12]。辛伐他汀片:北京万生药业有限责任公司(批号:31801043)。
谷胱甘肽过氧化物酶测试盒、超氧化物歧化酶测试盒、丙二醛测试盒、过氧化氢酶测试盒:南京建成生物科技有限公司。
DNA提取试剂盒:美国Omega生物技术公司;MiSeq测序试剂盒:美国Illumina公司;DNA凝胶回收试剂盒:美国Axygen公司;核酸纯化试剂盒:美国Beckman Coulter公司;文库定量试剂盒:美国KAPA公司。
SPF级雄性 Wistar大鼠,体质量(200±20)g,河北医科大学实验动物中心提供,许可证SCXK(冀)2018-004。
高脂饲料配方:基础饲料78.8%,猪油10%,蛋黄粉10%,胆固醇1%和胆酸盐0.2%。基础饲料由河北医科大学实验动物中心提供。基础饲料配方见表1。
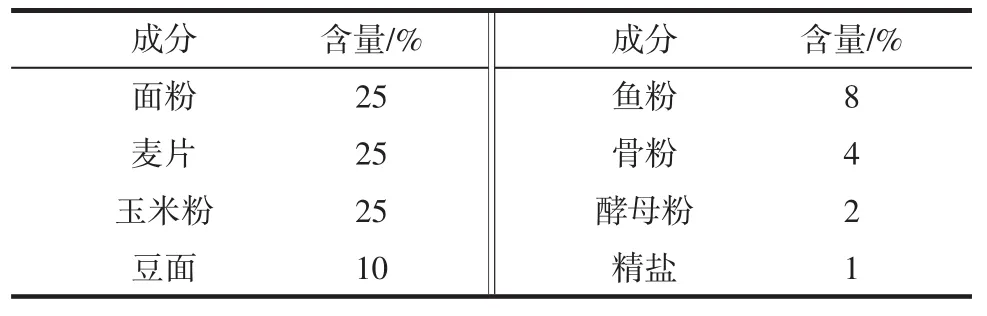
表1 基础饲料配方Table 1 The formula of basic feed
1.2 仪器与设备
高通量测序仪(PE300):美国Illumina公司;电泳仪(dyy-12):北京市六一仪器厂;凝胶成像系统(Alpha Imager HP):美国Protein Simple公司;生物分析仪(DE13806339):安捷伦科技有限公司;生物大分子分析仪(LabChip GX)、实时定量 PCR 仪(ABI 7500):美国Perkin Elmer公司。
1.3 方法
1.3.1 黑木耳多糖提取与鉴定
1.3.1.1 黑木耳多糖提取
将黑木耳用液氮冷冻法粉碎后过120目筛。采用水提醇沉法,在料液比 1∶60(g/mL)、温度 60 ℃、超声功率180 W的条件下提取20 min,于4 500 r/min离心6 min,收集上清液。重复提取3次,合并提取液干燥备用。用苯酚-硫酸法测定溶液中多糖含量[13]。
1.3.1.2 黑木耳多糖鉴定
参考莫开菊等[14]的方法,采用苯基-3-甲基-5-吡唑啉酮(1-phenyl-3-methyl-5-pyrazolone,PMP)柱前衍生化高效液相色谱(high performance liquid chromatography,HPLC)分析黑木耳多糖的单糖组成。HPLC条件:SHISEIDO C18色谱柱(4.6 mm×250 mm,5 μm),流动相A为0.1 mol/L KH2PO4(pH6.8);流动相B为乙腈;A∶B=82∶18(体积比);流速 1.0 mL/min;柱温 25 ℃;进样量10 μL;波长为245 nm。
1.3.2 黑木耳多糖对乳酸菌代谢产物影响
将对数生长期的乳酸链球菌按5%的接种量,接种到MRS液体培养基中,分别以0%、1%、5%、10%、20%、30%黑木耳多糖处理,37℃恒温培养14 h。每2 h取样并于4 200 r/min条件下离心20 min取上清液,采用考马斯亮蓝法[15]测定蛋白质含量。
1.3.3 黑木耳多糖体内益生作用分析
将36只SPF级Wistar雄性大鼠随机分为4组,分别为空白组(NC)、模型组(HFD)、辛伐他汀组(PCI)和复配物组(HDC),大鼠在自由采食高脂饲料的同时进行黑木耳多糖复配物干预,连续给药5周,具体处理方式见表2。

表2 实验设计方案(n=9)Table 2 Experimental design scheme(n=9)
1.3.4 检测样品采集
大鼠在末次灌胃12 h后,收集大鼠血清、肝脏和脑等器官,于-80℃保存备用。无菌收集结肠内粪便,置于无菌EP管中,迅速液氮冷却后置于-80℃冰箱保存备用。
1.3.5 抗氧化能力分析
根据试剂盒说明指示,依次对脑、肝脏组织中丙二醛(malondialdehyde,MDA)含量、过氧化氢酶(catalase,CAT)活性、超氧化物歧化酶(superoxide dismutase,SOD)活性和谷胱甘肽过氧化物酶(glutathione peroxidase,GSH-Px)活性进行测定。
1.3.6 Illumina高通量测序
对大鼠粪便样品进行16S rRNA基因V3~V4区域的聚合酶链式反应(polymerase chain reaction,PCR)扩展以及Illumina高通量测序。基于肠道菌群的16S rRNA基因V3~V4区域合成相关引物,进行PCR扩增,其中扩增所使用的引物序列为338F(5′-ACTCCTACGGGAGGCAGCAG-3′)和 806R(5′-GGACTACNNGGG TATCTAAT-3′)[16]。扩增的体系和扩增条件见表3与表4。

表3 16S rRNA基因V3~V4区PCR扩增体系Table 3 16S rRNA gene V3-V4 region PCR amplification system

表4 PCR扩增条件Table 4 PCR amplification condition
1.3.7 数据统计分析
数据处理采用SPSS20.0进行统计学分析。用单因素方差分析法分析组间差异的显著性,结果采用平均值±标准差表示,P<0.05表示差异具有统计学意义。
2 结果与分析
2.1 黑木耳多糖提取结果
黑木耳多糖提取结果见图1。

图1 黑木耳多糖提取结果Fig.1 Extraction results of Auricularia auricula polysaccharides
通过扫描电镜对黑木耳颗粒进行形态学观察,如图1(a)所示,冷冻粉碎处理的黑木耳样品具有较高的粒度级别,并具有良好的粒度均一性。
采用苯酚硫酸法测定多糖,绘制标准曲线,如图1(b)所示。获得回归方程为y=8.626 7x+0.140 3,相关系数R2=0.999 1。
本试验通过冷冻粉碎方法对黑木耳进行前处理,多糖的提取率达10.2%。
2.2 黑木耳多糖鉴定
单糖组成色谱图见图2。

图2 单糖组成色谱图Fig.2 HPLC chromatogram of PMP derivatives
由图2中各峰保留时间和峰面积,对黑木耳多糖的单糖组成进行分析,结果表明,黑木耳多糖中主要以葡萄糖、甘露糖为主,单糖比例葡萄糖∶甘露糖∶木糖∶岩藻糖∶半乳糖∶葡萄糖醛酸为 13.47∶8.64∶2.13∶1.95∶1.69∶1.00。
2.3 黑木耳多糖益生作用研究
黑木耳多糖对乳酸链球菌蛋白质代谢的影响结果如图3所示。

图3 黑木耳多糖对乳酸链球菌蛋白质代谢的影响Fig.3 Effects of different concentrations of polysaccharides on protein metabolism of lactic acid bacteria
当培养基中多糖含量为30%时,蛋白质含量随时间呈抛物线式变化。在8 h时,培养基中的蛋白质含量达到最大值114.48 mg/L,随后快速下降。多糖浓度0~20%时,乳酸菌的蛋白质代谢无显著变化。
2.4 体内抗氧化能力分析
黑木耳多糖复配物对大鼠血清抗氧化能力的影响见表5。

表5 黑木耳多糖复配物对大鼠血清抗氧化能力的影响Table 5 Effects of A.auricula polysaccharides complex on serum antioxidant capacity in rats
黑木耳多糖复配物具有较好的抗氧化能力。由表5可知,与NC组相比,HFD组大鼠血清CAT活力与SOD 活力显著降低(P<0.05),GSH-Px活力降低、MDA含量升高,但不存在显著差异(P>0.05)。与HFD组相比,PCI组与HDC组的GSH-Px活力、SOD活力均有不同程度的升高,MDA含量降低。其中HDC组SOD活力显著高于HFD组(P<0.05)。
黑木耳多糖复配物对大鼠肝脏抗氧化能力的影响见表6。
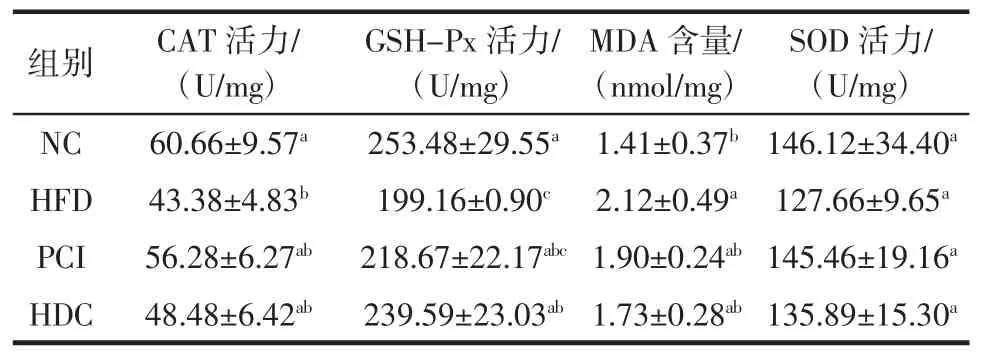
表6 黑木耳多糖复配物对大鼠肝脏抗氧化能力的影响Table 6 Effects of A.auricula polysaccharides complex on antioxidant capacity of rat liver
由表6可知,与NC组相比,HFD组大鼠肝脏CAT活力和 GSH-Px活力显著降低(P<0.05),MDA 含量显著升高(P<0.05)。与HFD组相比,HDC组GSH-Px活力显著升高。
黑木耳多糖复配物对大鼠脑抗氧化能力的影响见表7。

表7 黑木耳多糖复配物对大鼠脑抗氧化能力的影响Table 7 Effects of A.auricula polysaccharides complex on antioxidant capacity of rat brain
由表7可知,与NC组相比,HFD组大鼠脑组织GSH-Px活力显著降低(P<0.05),MDA含量显著升高(P<0.05),SOD活力虽低于NC组,但没有统计学差异(P>0.05)。与HFD组相比,HDC组MDA含量显著降低 P<0.05)。
2.5 大鼠肠道菌群物种丰度及多样性
2.5.1 分类单元聚类分析
分类单元(operational taxonomic units,OTU)交集维恩图见图4。

图4 OTU交集维恩图Fig.4 Venn diagram of OTU
试验以OTU相似性为97%对11个样本进行了OTU 分析,其中 NC 组(NC1~NC2),HFD 组(HFD1~HFD3)、PCI组(PCI1~PCI3)、HDC 组(HDC1~HDC3)。图4可以看出,4组OTU 分别为 626、692、673、656个。共有的为527个。
2.5.2 Alpha多样性分析
图5为大鼠肠道菌群Alpha多样性曲线。

图5 大鼠肠道微生物的Alpha多样性曲线Fig.5 Alpha diversity curve of intestinal microorganisms in rats
由图5(a)可以看出,随着抽样数量的增加,测得的OTU数量先增大,后趋于平缓。说明已经获得环境中绝大多数细菌序列,物种不会随着测序数量的增加产生明显的变化。从图5(b)可知,曲线逐渐趋于平缓状态,尽管增加抽样数量可以提高OTU数量,但是不会改变物种多样性。从图5(c)可以看出,随着样本量的增大,OTU数量从快速增加到趋于稳定,此时肠道菌群数量不再增多,所以此时的数据量可以用于后续实验。
图6和表8为各个样品Alpha多样性分析。

表8 肠道微生物Alpha多样性分析Table 8 Alpha diversity of gut micrabiota

图6 Alpha多样性指数Fig.6 Alpha diversity index
从图6和表8可以看出,HFD组大鼠的肠道菌群chao1指数最大,即肠道菌群丰度最高,但不存在显著性差异;HFD组大鼠肠道菌群Shannon指数显著高于NC、HDC组(P<0.05);即HFD组大鼠肠道微生态多样性明显高于NC、HDC组,原因可能是高脂饮食改变了肠道生态,有害菌过度繁殖,使肠道菌群多样性增加[17]。HDC组大鼠肠道菌群的丰富度以及多样性更接近于NC组。
2.5.3 Beta多样性分析
Beta多样性分析可以反映不同样本之间的菌群组成差异。图7为4组大鼠肠道菌群的组成,图中的点表示个体菌群,点与点之间的距离反映菌群之间的序列相似度。

图7 基于PLS-DA的4组大鼠肠道菌群组成分析Fig.7 Composition of intestinal flora of four groups of rats based on PLS-DA
从图7中可以看出,HFD组大鼠的肠道菌群构成与NC组差别较大。PCI组与HDC组大鼠的肠道菌群构成与NC组相近,其中HDC组更接近正常大鼠。说明该复配物对肠道菌群具有正向调节作用。
2.6 大鼠肠道菌群结构分析
2.6.1 门水平差异分析
大鼠肠道菌群结构在门水平的差异见图8。

图8 大鼠肠道菌群结构在门水平的差异Fig.8 Microbial community structure at the phylum level
由图8可知,在门水平上,大鼠肠道中最多的是厚壁菌门,其次是拟杆菌门。HDC组大鼠肠道内的放线菌门(16.41±1.81)%显著高于 HFD 组(8.6±1.72)%(P<0.05)。HDC组拟杆菌门丰度较NC组和HFD组有下降的趋势,但不存在显著性差异。
2.6.2 属水平差异分析
大鼠肠道菌群结构在属水平的差异见图9。

图9 大鼠肠道菌群结构在属水平的差异Fig.9 Microbial community structure at the genus lev
由图9可知,在属水平上,NC组大鼠肠道中菌属相对丰度由高到低为 Lactobacillus、Romboutsi、Rumino-coccaceae UCG-014、Ruminococcus1、Ruminococcaceae NK4A214_group等。HFD 组(10.7±4.23)%、PCI组(4.40±1.11)%的乳酸菌相对丰度较 NC组(37.71±0.63)%均极显著降低(P<0.01);与HFD相比,HDC组Alistipes、螺杆菌、脱硫弧菌属含量分别降低0.10%、0.34%、0.50%,Dorea、志贺氏菌含量均降低0.11%;双歧杆菌、乳酸菌、乳球菌含量分别增加3.06%、9.12%、0.04%。
3 讨论与结论
本实验提取黑木耳多糖的单糖组成与庄伟等[7]的研究结果基本相符。结合已有的研究,黑木耳多糖的单糖组成与黑木耳来源相关,不同品种的黑木耳的单糖组成不同[18]。黑木耳多糖作为益生元,可有效促进乳酸菌的增殖[19]。薛依婷等[20]研究发现黑木耳多糖促进了酸乳中保加利亚乳杆菌、嗜热链球菌、干酪乳杆菌的繁殖,并提高了酸乳中游离脂肪酸和胞外多糖的含量。Zhang等[21]研究发现黑木耳多糖的干预逆转了高脂血症大鼠中乳酸杆菌和Oscillibacter丰度下降的趋势。本实验将不同浓度的黑木耳多糖与乳酸菌共培养,培养过程中发现,当乳酸菌与30%黑木耳多糖共培养至8 h时,采用考马斯亮蓝法测定溶液中蛋白质浓度(114.48 mg/L)与空白组相比明显提高。考马斯亮蓝法(Bradford法)测定溶液中的蛋白质含量,其工作原理为染料考马斯亮蓝G-250与蛋白质中的碱性氨基酸或芳香族氨基酸结合,在595 nm波长下有最大光吸收[22]。因此推测,随着培养时间的延长,黑木耳多糖提高乳酸菌蛋白酶类的代谢水平,促进溶液中蛋白质的降解,使得特征氨基酸充分暴露出来并与染料结合,进而提高了其吸光度。
高脂饮食可以调节大鼠肠道菌群结构,从而对机体的健康产生影响。本研究中发现,高脂饮食大鼠肠道菌群与空白组相比产生了明显的差异,肠道中的有害菌增加,如志贺氏菌、螺杆菌、β-变形菌纲、伯克霍尔德氏菌目、梭菌属、粪球菌属等;有益菌的相对丰度降低,如乳酸杆菌、双歧杆菌等。将黑木耳多糖与银耳多糖和山楂黄酮复配后,对高脂饮食大鼠进行干预,观察到大鼠的肠道菌群结构发生变化,其结构向正常大鼠的肠道菌群结构转变,有害菌减少,乳酸菌等有益菌的增多。这与Zhang等[21]的研究结果一致。其原因可能是黑木耳多糖影响了乳酸菌的蛋白质水解系统,增强了胞外蛋白酶活性,促进了蛋白质水解为多肽。多肽通过乳酸菌的多肽转运系统进入细胞,促进了乳酸菌的代谢水平。乳酸菌进一步抑制肠道有害菌,改善肠道菌群结构,并调节宿主代谢[23-25]。另一方面,在蛋白质水解过程中还可以产生其它分子,如与乳酸菌益生作用相关的生物活性肽[26]。
综上所述,黑木耳多糖对乳酸菌的代谢产物具有一定的调节作用,并且黑木耳多糖复配物对高脂饮食造成的肠道菌群紊乱具有调节恢复作用,同时可以在一定程度上增强机体的抗氧化能力。
