葡萄Vv SUC27基因的克隆及病原响应表达分析
2021-11-12吴佳鸿黄金宝刘梅等
吴佳鸿 黄金宝 刘梅等

摘要 由葡萄座腔菌科Botryosphaeriaceae真菌引起的葡萄溃疡病(Botryosphaeriadieback)是葡萄上的主要枝干病害之一,严重影响葡萄产业的健康发展。其中,致病力最强的为可可毛色二孢Lasiodiplodia theobromae。糖转运蛋白在调节植物对病原菌的免疫过程中起着至关重要的作用,关于葡萄中糖转运蛋白参与植物免疫机制的研究尚未见报道。本试验从‘无核白葡萄叶片中克隆得到蔗糖转运蛋白VvSUC27基因,其开放阅读框全长1515bp,预测蛋白质分子量为54kD,理论等电点(pI)为9.58。生物信息学预测该蛋白含有12个跨膜结构。实时荧光定量PCR(qRTPCR)分析表明:氨基环丙烷羧酸和脱落酸处理后VvSUC27的表达水平持续上调;水杨酸和茉莉酸甲酯处理后,VvSUC27的表达呈现上调表现但无规律的变化特征;激发子chitin和flg22处理后,VvSUC27基因信号响应快速且明显;感病品种‘夏黑接种可可毛色二孢后,VvSUC27被迅速诱导,3h时表达量达到最高,而后逐渐减弱。本研究将为今后阐明可可毛色二孢与葡萄互作的分子机制提供参考依据。
关键词 葡萄溃疡病; 可可毛色二孢; 蔗糖转运蛋白基因; 基因克隆; 表达分析
中图分类号: S663.1 文献标识码: A DOI: 10.16688/j.zwbh.202016
葡萄犞犻狋犻狊狏犻狀犻犳犲狉犪集鲜食与加工于一身,是全球广泛栽培的重要经济果树之一。葡萄枝干病害(grapevinetrunkdiseases,GTDs)是由多种病原真菌引起的危害葡萄枝干的病害统称。由葡萄座腔菌科Botryosphaeriaceae真菌引起的葡萄溃疡病(Botryosphaeriadieback)是葡萄主产区最常见的枝干病害之一,病原菌主要通过伤口或自然孔口侵入,导致树干变色坏死、果实枯萎腐烂,严重的整株植株枯死。近年来该病引起我国浙江、江苏、广西、湖南、河北等地葡萄减产30%~50%。在我国葡萄产区,本研究团队对引起葡萄溃疡病的真菌进行了形态观察及分子鉴定,发现了包括葡萄座腔菌Botryo-sphaeria dothidea、Diplodia seriata、可可毛色二孢Lasiodiplodia theobromae、小新壳梭孢Neofusicoccum parvum、Lasiodiplodia、pseudo-theobromae和Neofusicoccum mangiferae在内的6种葡萄座腔菌科真菌。其中,L.theobromae和B.dothidea是主要优势种群,L.theobromae是致病力最强的一种。葡萄溃疡病已成为我国葡萄健康发展的重大障碍,但目前对该病害的致病机理还不明晰,生产上也还没有建立起对应的高效防控措施。糖类不仅是植物生长发育的必要因素,也是病原菌从宿主细胞吸收的主要碳源和能源物质。糖转运蛋白不仅介导植物细胞内、细胞间糖的摄取与释放,也介导植物病原体中糖的分配,其在植物质膜上的分配是植物生长发育和应对生物及非生物胁迫时最重要的过程之一。植物中糖转运蛋白主要分成单糖转运蛋白(MSTs)和蔗糖转运蛋白(sucrose transporters/sucrose carriers,SUTs/SUCs)两类,这两类糖转运蛋白都属于MFS蛋白超家族(Majorfacilitatorsuperfamily)。蔗糖转运蛋白(SUTs/SUCs)包含SUT1、SUT2、SUT3、SUT4和SUT55个亚族。已知的SUTs其氨基酸序列高度保守,为疏水性蛋白,包含由12个α螺旋组成的保守跨膜结构域,中间面向胞质的亲水胞质环将蛋白分为两部分,每部分含6个跨膜结构域,伸展的N 端序列也定位于胞质。SUTs基因家族成员在植物的不同生长阶段和不同部位表达,行使多种生物学功能,参与多种信号转导途径。植物能够调节糖转运蛋白,将糖从感染部位重新分配出去,改变病原菌获得营养物质的途径,消除病原体的能量来源,来限制它们的扩散。玉米黑粉菌蔗糖转运蛋白(UmSRT1)对蔗糖高度亲和,寄主植物的ZmSUT1与其竞争蔗糖,病原菌通过直接吸收蔗糖来避免利用葡萄糖,进而诱导防御机制。犝犿犛犚犜1 的缺失或ZmSUT1的上调表达都会造成病原菌毒力大大降低。马铃薯中过表达蔗糖转运蛋白SoSUT1导致丛枝菌根(arbuscularmycorrhizae,AM)真菌的定殖率增大。
迄今为止,预测葡萄基因组中有3个蔗糖转运蛋白基因:VvSUC11,VvSUC12,VvSUC27,这3个基因均可在果实中表达,并且具有转运蔗糖的功能。但对于葡萄中蔗糖转运蛋白参与免疫机制的调控目前还未见报道。前期的试验中以可可毛色二孢菌株CSS01s、高感葡萄‘夏黑为研究对象,对可可毛色二孢侵染的‘夏黑葡萄进行转录组分析,随后通过差异表达基因的比对注释,系统分析了侵染初期感病葡萄对可可毛色二孢防御反应相关基因的表达,发现蔗糖转运蛋白基因VvSUC27在感病样品中存在差异表达。本研究从葡萄的cDNA中克隆获得葡萄蔗糖转运蛋白VvSUC27基因,利用生物信息学和实时荧光定量PCR方法对其进行结构和表达分析,研究结果为进一步阐明VvSUC27基因的生物功能奠定了基础,同时也为解析其参与免疫反应的调控机制提供参考。
1材料与方法
1.1 试验材料
试验于2019年9月在北京市农林科学院植物保护环境保护研究所进行。葡萄‘夏黑枝条由北京市农林科学院林果所提供。葡萄品种‘无核白组培苗由中国农业科学院兴城果树研究所董雅凤研究员提供。
可可毛色二孢菌株CSS01s由本实验室分离并保存。LA Taq DNA 聚合酶、pMD18T 载体、SYBRPremixExTaqTM购自宝生物工程有限公司。反转录酶SuperScript Ⅲ购自Invitrogen公司。E.coli Trana5α感受态细胞购自北京全式金生物技术有限公司。其他常规化学药品均为分析纯。农杆菌EHA105菌株和SuperGFP植物表达载体由本实验室保存。flg22(鞭毛蛋白N端含保守的22个氨基酸的多肽:QRLSTGSRINSAKDDAAGLQIA,纯度>95%)由艾博抗(上海)贸易有限公司合成。几丁质chitin(C9752)购自Sigma公司。植物激素,水楊酸(SA)、脱落酸(ABA)、茉莉酸甲酯(MeJA)、氨基环丙
烷羧酸(ACC)购自SigmaAldrich公司。
1.2 试验方法
1.2.1 接种处理
葡萄‘夏黑休眠枝条种植于北京市农林科学院温室,将生根枝条以20cm×20cm 的间距移栽至25cm×25cm 的花盆中培养3 个月。接种按照Yan等方法稍作修改:将可可毛色二孢菌株CSS01s在PDA 培养基上于28℃培养2d;半木质化新生嫩枝用70%乙醇进行表面消毒后,在茎中间用打孔器制造2mm 深的伤口,取直径为4mm 的菌饼接种到伤口上并使用封口膜稍作固定,对照组接种无菌PDA 培养基块;接种后的葡萄苗置于26~28℃、光周期L∥D=12h∥12h、相对湿度为85%~95%的植物接种室中;接种0、1、3、6、12h和24h时以伤口为中心截取4cm 范围内全部韧皮组织,采集的样品在液氮中速冻,-80℃冰箱保存备用。
1.2.2 激素及激发子处理
参照Jiao等的方法,将生长情况一致的‘无核白组培苗移栽至26~28℃人工气候室,光周期L∥D=16h∥8h下培养3个月后,对材料进行激素及激发子处理:将100μmol/L水杨酸、100μmol/L脱落酸、100μmol/L茉莉酸甲酯、100μmol/L氨基环丙烷羧酸、1μmol/Lflg22和10μg/mL几丁质分别喷施在葡萄叶片上,喷施量为(1.5±0.1)mL/株,同时以等体积灭菌ddH2O处理为对照;暗培养并分别在处理0、1、3、6、12h和24h时采集叶片,每种处理设置3组重复。采集的葡萄叶片用液氮速冻后,保存于-80℃冰箱备用。
1.2.3 RNA 提取及cDNA 合成
取上述激素处理、激发子处理和接种处理后的植物组织,采用多糖多酚/复杂植物RNA 快速提取试剂盒(北京艾德莱生物科技有限公司)提取样品总RNA,琼脂糖凝胶电泳检测RNA 的质量及完整性。NanoDrop2000(Thermo)对RNA 进行定量后,利用SuperScriptⅢ反轉录试剂盒(Invitrogen)反转录合成cDNA,-20℃保存备用。
1.2.4 VvSUC27基因ORF的克隆及生物信息学分析
根据VvSUC27 序列(GenBank 登录号:AF021810.2)设计特异性引物:上游引物SUC27F:5′ATGGAGTTAGCCAAGCCTTCTTCAG3′,下游引物SUC27R:5′AGACGACGGCTGAGTCCTCTCATCC3′。以葡萄‘无核白cDNA 为模板,进行PCR扩增,反应体系(50μL):10×PCRbuffer5μL;2.5mmol/LdNTPs2μL;10μmol/L上游和下游引物各1μL;LA Taq0.4μL;cDNA 模板2μL;补水至50μL。反应程序:94℃预变性5min;94℃变性30s,60℃退火30s,72℃延伸90s,32个循环;72℃延伸10min。PCR产物经1.5%琼脂糖凝胶电泳检测后,用胶回收试剂盒(Qiagen)回收目的片段,连接到pMD18T载体上,定名为SUC27T,转化大肠杆菌Trans5α感受态细胞,单克隆鉴定为阳性后送北京擎科新业生物技术有限公司测序。采用DNAMAN6.0、ProtParam等软件对其蛋白序列进行生物信息学分析;利用MEGA7构建系统进化树。
1.2.5 基因表达分析
根据VvSUC27 基因序列设计实时荧光定量PCR特异引物,上游引物qS27F:5′ATCACCTACAGCATTCCAT3′,下游引物qS27R:5′CTACCGACACCATCATCT3′。以1.2.3制备的cDNA作为模板,每个样品设3 次重复,用VvEF1γ(AF176496)作为内参基因。参照TaKaRa说明书,采用SYBRGreenI染料法进行荧光定量PCR,反应在ABI7500 RealtimePCR 检测系统上进行。PCR 反应体系:SYBR Premix ExTaqTM10μL;10μmol/L上游和下游引物各0.5μL;ROXReferenceDyeⅡ0.4μL;cDNA 模板0.5μL;补ddH2O至20μL。反应程序为:95℃预变性30s;95℃变性5s,60℃ 退火34s,40 个循环;熔解曲线为95℃15s,60℃60s,95℃30s,60℃15s。VvSUC27基因的相对表达水平用2-△△α犆狋法计算。
2 结果与分析
2.1 葡萄VvSUC27基因的克隆
以葡萄品种‘无核白cDNA为模板,SUC27F/R为引物扩增得到1条1500bp左右的片段,测序并通过DNAMAN软件将得到的序列与‘黑比诺参考序列(PN40024)进行比对发现,其开放阅读框(ORF)序列长度一致,为1515bp;第164位核苷酸与参考序列存在差异,‘黑比诺中该位置为T,对应氨基酸为亮氨酸(Leu);‘无核白中为A,对应氨基酸为组氨酸(His)(图1)。
2.2 VvSUC27蛋白的生物信息学分析
2.2.1 VvSUC27蛋白的理化性质分析
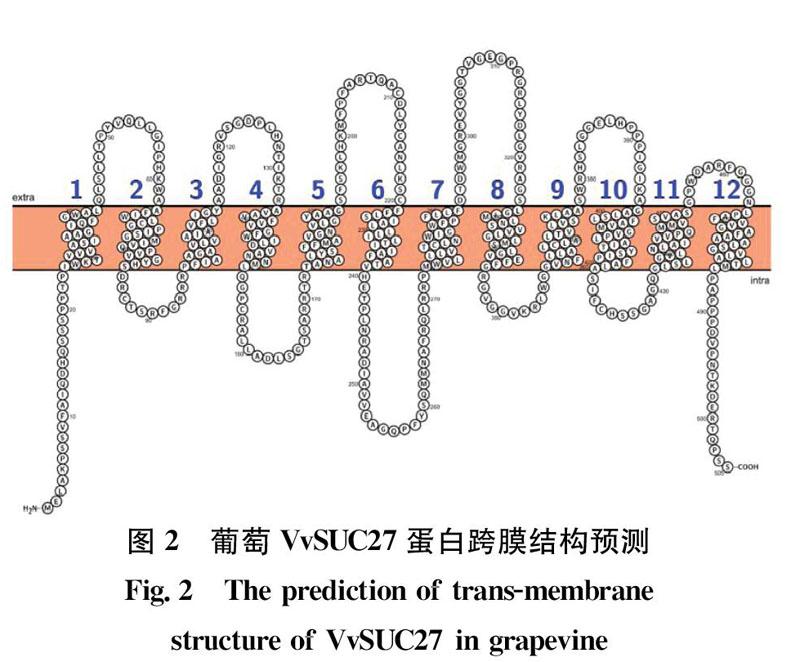
VvSUC27基因编码505个氨基酸,包含22个酸性氨基酸(Asp+Glu),36个碱性氨基酸(Arg+Lys),编码蛋白相对分子量为54kD,理论等电点(pI)为9.58。VvSUC27蛋白包含44.36%的α 螺旋(al pha helix)、14.65% 的延伸链区(extendedstrand)、4.16%的β转角(beta turn)和36.83%的无规则卷曲(randomcoil)。应用在线工具(http:∥wlab.ethz.ch/protter/start/)预测VvSUC27蛋白含有12个跨膜区(图2)。
2.2.2 VvSUC27蛋白三 级结构预测
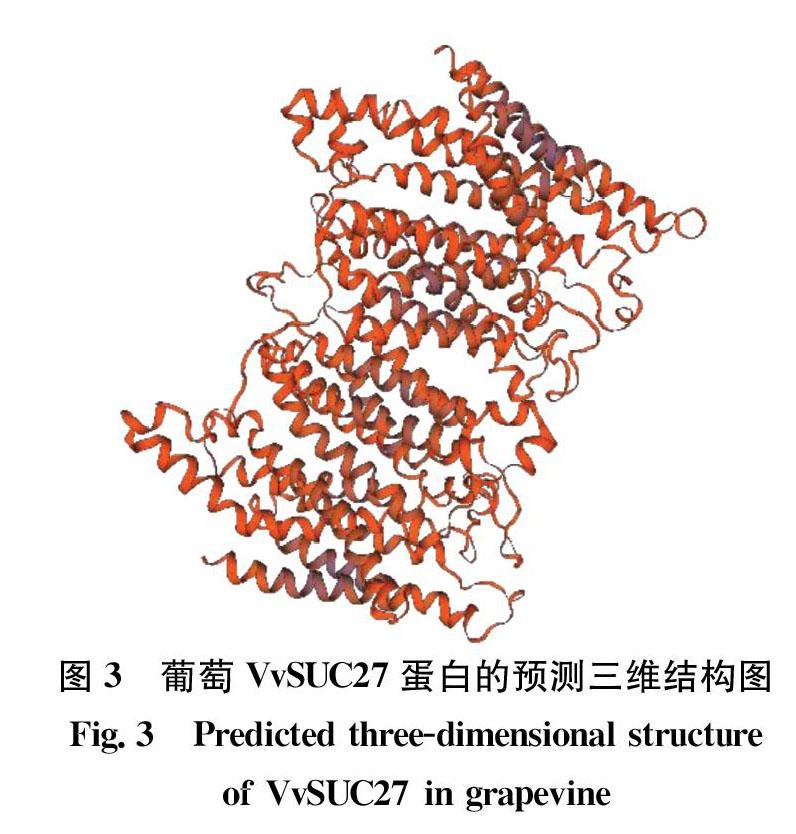
用SWISSMODEL对VvSUC27编码氨基酸的三级结构进行预测,结果如图3所示。葡萄VvSUC27蛋白的三级结构域呈现复杂弯曲的交叉螺旋状结构,有12个明显的α螺旋。
2.2.3 VvSUC27蛋白的系统进化树
通过同源性比对分析表明,葡萄VvSUC27蛋白与烟草NtSUC1a(CAA57727)蛋白和拟南芥AtSUC5(AEE35248)同源性最高分别为63.92% 和61.43%,说明其亲缘关系最近(图4)
2.3 VvSUC27基因的表达分析
激素处理后,VvSUC27基因表达量发生变化(图5):水杨酸(SA)处理后,VvSUC27基因表达量呈先上升后下降的趋势,处理12h后表达量最高,约为对照的2 倍;茉莉酸甲酯(MeJA)处理后,VvSUC27基因表达量呈先上升后下降的趋势,处理6h后表达量最高,约为对照的2倍;乙烯前体氨基环丙烷羧酸(ACC)处理后,犞狏犛犝犆27基因在1h时表达量最低,低于对照的1/2;处理12h后基因表达量上调。脱落酸(ABA)处理后,VvSUC犆27基因表达量有所下降,6h后恢复为与对照相当水平,而后不断上升,处理24h后达到峰值。
如图6所示,在chitin诱导下,VvSUC27基因信号响应十分快速且明显,表达量在3h时达到峰值,约是对照的6倍,而后表达量逐渐降低;在flg22处理下,VvSUC27基因表达量升高,其中12h时表达量最高,约为对照的4倍,处理24h后表达量则基本恢复至与对照水平相当。
接种可可毛色二孢后,VvSUC27表达量先升高,3h时达到峰值,约为对照的2.5倍;随后表达量开始下降,接种12h时最低,约为对照的2倍(图7)
3 讨论
蔗糖转运蛋白(SUTs/SUCs)是植物中特有的一类跨膜蛋白,在植物中广泛存在,负责蔗糖经韧皮部从源到库的运输以及库组织的蔗糖供给,还参与蔗糖在库组织的贮存。蔗糖转运蛋白是一个十分庞大的家族,他们的共同特点是具有12个跨膜区的蛋白质,在12个跨膜区域中,氨基酸是极为保守的,其中的疏水氨基酸几乎完全不变,而N 端与C端为高变异区。最保守的跨膜区为第1、2和11跨膜区,但不同植物或相同植物的不同基因存在功能上的差异。Davies等于1999年从葡萄果实中克隆得到了3个推测可能是葡萄蔗糖转运蛋白的基因。张雅丽等研究发现:VvSUC27在葡萄的根、幼叶以及果实等库器官中大量表达,而在叶片中表达量很少,与Davies结果一致,初步认为该基因很可能编码特异性存在于库组织中的蔗糖转运蛋白;于菲等2011年对VvSUC27启动子进行了初步的功能预测分析。本研究克隆了葡萄VvSUC27 基因,利用生物信息学方法对该蛋白的理化性质、三级结构进行了预测分析。结果表明其cDNA 全长为1515bp,编码505个氨基酸,VvSUC27蛋白具有12个跨膜结构,与烟草蔗糖转运蛋白NtSUC1a的亲缘关系最近,为63.92%,VvSUC27基因与许多已经验证功能的糖转运蛋白基因都有很高的同源性,具有糖转运蛋白的活性。
目前糖转运蛋白在生理以及非生物胁迫方面的作用已经有了较深入的研究。植物中经光合作用产生的糖分需要其运输调控以保证自身各部分的营养供应,非生物胁迫条件下糖转运蛋白也会参与以适应逆境。在玉米、小麦、马铃薯等植物中发现了参与植物与病原菌互作过程相关的糖转运蛋白基因。本研究通过对葡萄受可可毛色二孢侵染后不同时间VvSUC27基因表达量的检测,证实病菌侵染诱导寄主植株VvSUC27 基因的表達量增加,推测VvSUC27基因参与了寄主对可可毛色二孢的调控。水杨酸、脱落酸、茉莉酸以及乙烯是植物免疫相关的激素,在不同病原菌侵染过程中,其在寄主体内通过信号分子积累的改变来调节植物的免疫反应。本研究结果表明,SA 和MeJA 处理6h即可诱导VvSUC27上调表达;而ABA 和ACC 处理24h后,VvSUC27基因才出现诱导表达。在病原菌与寄主的互作过程中,寄主植物细胞表面的模式受体(patternrecognitionreceptors,PRR)通过识别病原菌保守的病原相关分子模式(pathogenassociatedmolecularpatterns,PAMPs)激发PAMPs诱导的免疫(PAMPtriggeredimmunity,PTI),其中细菌的鞭毛蛋白(flagellin)、真菌的几丁质(chitin)、肽聚糖(peptidoglycan,PGN)等都是常见的病原菌保守组分。VvSUC27受flg22和chitin的诱导表达,推测其参与寄主植物的PTI反应。
本文克隆了VvSUC27基因并分析了其在接种可可毛色二孢后以及不同激素和激发子处理下的表达情况。下一步将开展基因编辑、蛋白互作等工作,对葡萄中蔗糖转运蛋白的功能及作用机理进行深入研究。通过探索VvSUC27基因在可可毛色二孢与葡萄互作中的作用机制,为葡萄中其他蔗糖转运蛋白调控机制研究提供一个新的切入点,为葡萄的遗传改良提供理论参考。
