降水变化和生长方式对红砂和珍珠化学计量特征的影响
2021-11-09单立山魏晓芸解婷婷
杨 洁, 单立山, 苏 铭, 魏晓芸, 马 静, 解婷婷, 李 毅
(甘肃农业大学林学院, 甘肃 兰州 730070)
第五次联合国政府间气候变化专门委员会(Intergovernmental Panel on Climate Change,IPCC)评估报告显示,全球气候变暖加剧影响全球降水量发生明显变化[1],中国西北干旱区是全球中纬度地带最干旱、对全球气候变化响应最敏感的地区[2],区域内降水量逐年减少且季节性波动加剧[3]。水分是影响植被生长发育的关键因子[4],降水变化会显著影响植物各器官生态化学计量特征。C,N是构建植物组织和表征生态化学计量特征的主要元素,研究降水变化对不同植物不同器官C,N化学计量特征及分配的影响,可为探索植物不同器官的营养分配和资源利用规律及其对环境变化响应的内在机制提供依据[5]。
降水量的改变使土壤养分含量发生变化,进而影响植物光合生理过程以及营养元素的吸收和利用,最终导致不同植物各器官中C,N含量及C/N存在较大差异[6]。有研究发现随着降水量减少,植物叶、茎中C和N含量逐渐增加,C/N降低[7-8]。但也有研究结果表明,降水量减少导致的干旱胁迫使植物叶片C和N含量降低,C/N增加[9-11]。根系作为植物运输和吸收水分、养分的主要渠道,其C,N化学计量特征也可以反映植物对养分、水分的吸收和利用情况[12]。有研究发现植物根部N含量随土壤水分的减少而减少[13]。也有研究指出适度的水分胁迫会导致植物根系N含量减少,但随干旱胁迫程度进一步加剧,植物根系N含量不再呈现下降趋势[14]。此外,还有研究表明植物根部的N含量在干旱地区会显著上升[15]。由此说明,降水变化对植物不同器官C,N化学计量特征影响的研究仍无统一定论。然而,植物各器官的C,N化学计量特征及分配不仅受降水量的影响,而且会受到物种生长方式的影响。许多研究发现不同物种混生生长能显著提高植物叶片和根系的C,N含量[16-20]。也有研究结果与此相反,发现2种植物混生会导致其中一种植物部分器官的C,N含量降低[17,21-22],或者混生后植物叶片的C,N含量无显著变化[23]。可见,生长方式对植物不同器官C,N含量的影响会因物种和试验条件等因素存在明显差异。目前有关植物不同器官C,N化学计量特征变化的研究主要集中在单一因素对森林和草原等生态系统植物的影响[24-25],由于荒漠生态系统复杂性和变异性较大,同时荒漠植物根系取样困难[26],因此,针对生长方式与降水量的交互作用对干旱区典型荒漠植物红砂(Reaumuriasoongorica)和珍珠(Salsolapasserina)影响的相关研究较少[27]。因此,结合植物生长方式,开展降水梯度对荒漠植物不同器官化学计量特征影响的研究,对探讨不同生长方式下荒漠植物应对降水变化的养分调控能力和机制具有重要意义。
红砂和珍珠是广泛分布于荒漠区的植物种,2者常互为共建种构成混生群落[28],广泛分布于荒漠草原区、典型荒漠区和极端荒漠区,分布区的年降水量自东向西递减[29]。近年来,关于2种植物混生后的生理生态变化特征的研究主要集中在光合特性[29]和荧光特性[30],干旱胁迫下2者混生后的根系变化特征也有少量报道[31]。然而针对不同降水量下混生后红砂和珍珠不同器官化学计量特征变化的研究较缺乏。因此,本研究通过人工模拟降水试验,测定不同降水量条件下单生和混生2种生长方式的红砂和珍珠不同器官的C,N含量,试图回答以下几个问题:随干旱胁迫程度的加剧,红砂和珍珠分别如何调控不同器官的C,N含量以应对干旱胁迫;与单生相比,混生后红砂和珍珠不同器官的C,N含量是否会发生变化,若发生变化,两种植物分别如何调整C,N含量在不同器官的分配以达到共存。对这些问题的回答,有利于揭示典型荒漠植物在干旱胁迫生境中的养分调控策略和共存机制,为荒漠区红砂和珍珠群落植被的保护和恢复等提供理论指导。
1 材料与方法
1.1 研究区概况
采样地甘肃省白银市景泰县(36 °43′N,104 °43′E)海拔1 627 m,属温带干旱大陆性气候,年均气温为9.1℃,无霜期在141 d左右,年均日照2 713 h,年降水量为185.6 mm,年蒸发量为1 722.8 mm,土壤类型主要是黄土、洪积灰棕荒漠土和灰钙土,其地带性植被有红砂、珍珠、盐爪爪(Kalidiumfoliatum)、画眉草(Eragrostispilosa)、白刺(Nitrariatangutorum)、芨芨草(Achnatherumsplendens)等荒漠植物。试验地甘肃农业大学(36 °09 ′N,103 °70 ′E)海拔1 572 m,属温带大陆性气候,年平均气温8.9℃,年日照时数2 476.4 h,年降水量349.9 mm,年蒸发量1 664.0 mm。
1.2 试验材料
2017年3月底4月初植物开始生长之前,在甘肃省白银市景泰县红砂和珍珠广泛分布的山前戈壁设置20 m×20 m的样方(3个重复)进行群落学特征调查,调查数据显示,该生境内红砂植株密度为1.28株·m-2,珍珠植株密度为0.86株·m-2,以该生境中红砂和珍珠的平均样株大小为标准,分别随机选取大小均一(平均株高(10±0.2) cm、平均冠幅(10±0.2) cm的一年生单生红砂、单生珍珠以及混生红砂和混生珍珠幼苗各36株。将幼苗带回甘肃农业大学校内试验地栽植到自行设计的半径为15 cm、高度为80 cm的PVC管内,管内装入取自采样地自然生境中的土壤,加入少量蛭石和珍珠岩混合避免土壤板结,土壤有机碳4.16 g·kg-1,全氮0.32 g·kg-1,全磷0.54 g·kg-1,容重1.28 g·cm-3。为防止水分流失和土壤下沉,在管子底部均放置了稍大于管子的托盘。缓苗1个月,期间为保证幼苗存活,对植株进行水分和病虫害管理。
1.3 试验设计
本研究根据红砂和珍珠的自然分布规律由东向西选取6个地点的降水量作为参照(参照点的水热条件如表1所示)。综合6个选取点的气象数据(依据中国气象局1951年01月01日—2008年12月31日近58年的数据资料),本研究将人工模拟降水量分别设置为50,100,150,200,250和300 mm,单生红砂、单生珍珠、混生红砂和混生珍珠在6个模拟降水量下的处理均为3个重复。调查研究发现,西北荒漠区降水少且不稳定,主要集中在夏秋季,占全年降水量的70%~80%[32]。因此,在模拟降水量的试验中,计算出6,7,8,9这4个月的降水量分别占全年降水量的比例并换算为相应的浇水量。降水量换算方法参考闫建成等[33]和寇祥明等[34],换算方法如下:在直径为20 cm的雨量器内,每降水31.4 mm相当于自然降水1 mm:
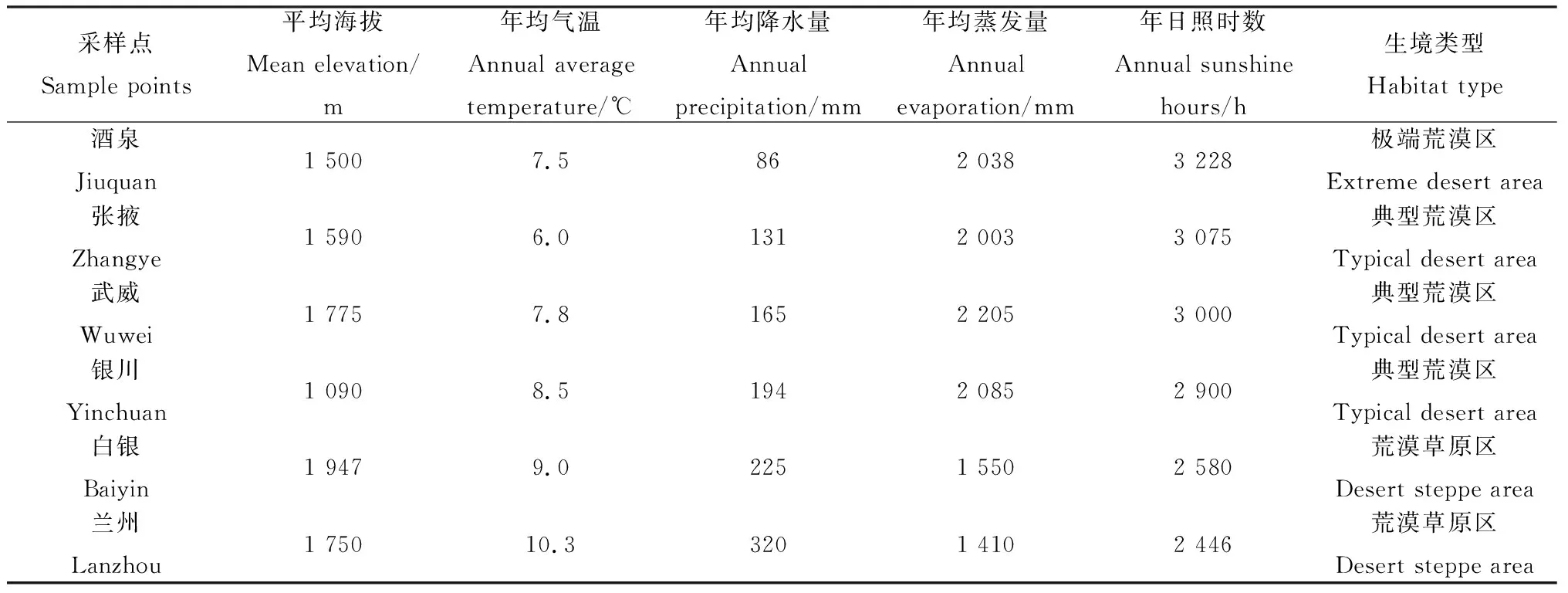
表1 试验设计参照的试验6个选取点的水热条件比较Table 1 Hydrothermal condition of six different habitats referred to experiment design
式中,A代表降水量,S代表降水面积,即在面积为S的地面上降水AmL相当于1 mm降水。单次降水量及浇水量具体量见表2,各降水处理的浇水频率分别为:模拟降水量为50 mm和100 mm的每月4次,150 mm和200 mm的每月6次,250 mm和300 mm的每月8次,时间为6月1日起至9月30日。

表2 降水量占比及浇水量换算Table 2 Proportion of monthly rainfall and conversion of irrigation volume

续表2
1.4 样品获取和指标测定
取样于10月1日进行。样品采集时,将所有植株的地上部分剪下,叶和茎分离后装入标记好的信封中,用200 μm的筛子将根系中土块和石子等杂质过滤后,将根系用蒸馏水冲洗干净装入有标记的信封。之后将根茎叶立即带回实验室,105℃杀青30 min后,80℃烘至恒重,粉碎后用于碳和氮含量测定。其中,有机碳含量测定采用重铬酸钾氧化加热法[32],全氮含量测定采用凯氏定氮法[35-36]。
1.5 数据统计和作图
本试验全部数据采用SPSS 22.0进行统计分析。在进行方差分析之前,首先根据正态直方图判断所有数据是否符合正态分布,对不符合正态分布的数据进行对数转化,使所有数据符合正态分布,其次采用Levene检验进行方差齐性检验,P大于0.05,表明数据符合方差齐性假设。在此基础上,生长方式和降水量及其交互作用对2种植物根茎叶的C,N化学计量特征指标的影响采用双因素方差分析(Two-way ANOVA),对同一生长方式不同降水量下同种植物根茎叶的C,N化学计量特征指标采用单因素方差分析(One-way ANOVA),对同一降水量不同生长方式下同种植物根茎叶的C,N化学计量特征指标进行独立样本t检验。作图采用Origin 2018,图中数值均为平均值±标准误差。
2 结果与分析
2.1 不同降水量下叶、茎、根有机碳含量及其比重比较
由表3可知,红砂和珍珠根、茎和叶的有机碳含量在不同生长方式间差异显著(P<0.05);红砂和珍珠茎有机碳含量在不同降水量间差异显著(P<0.05),珍珠根有机碳含量比重在不同降水量间差异达到极显著(P<0.01)。
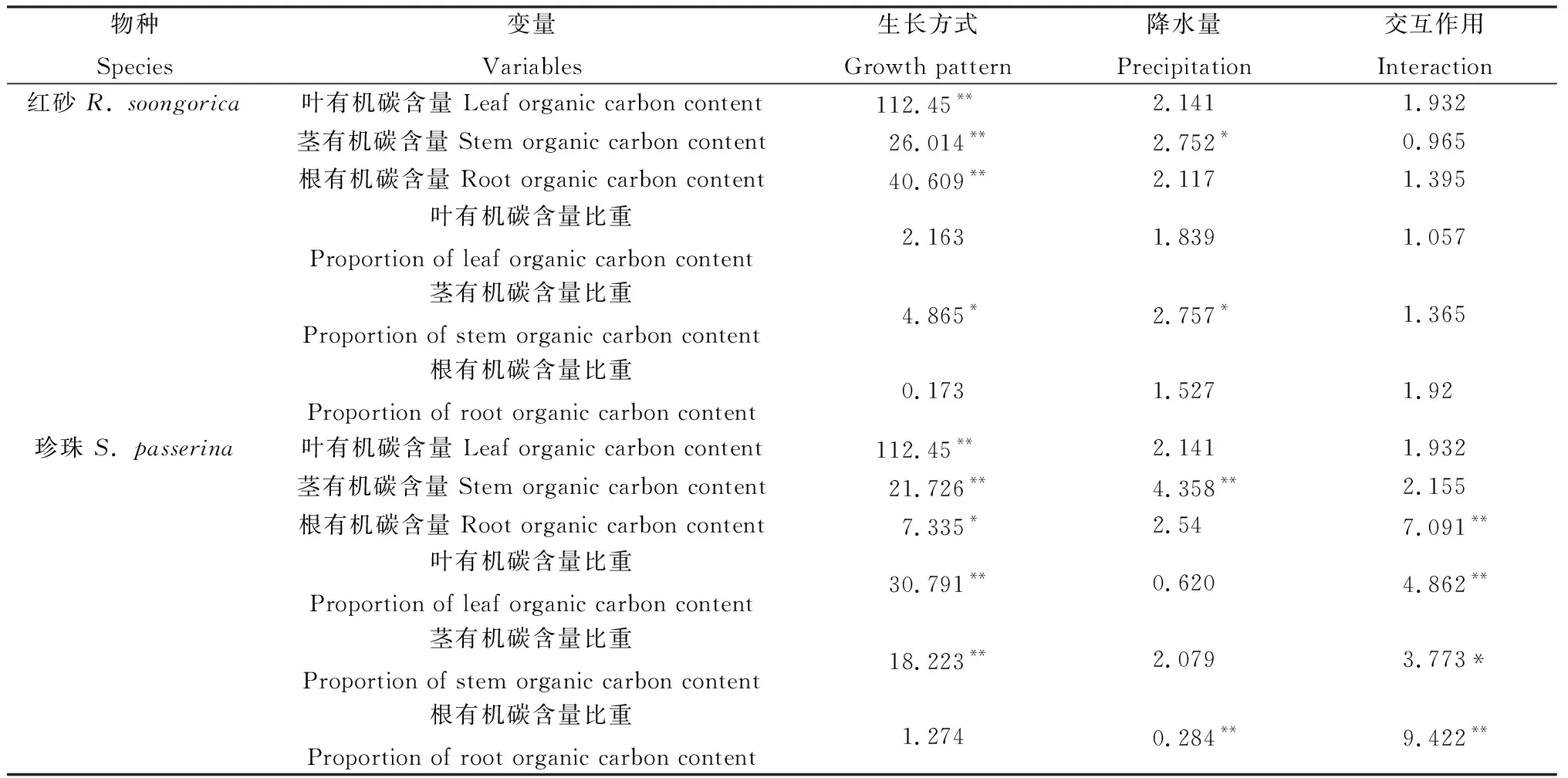
表3 不同降水量下单生、混生红砂珍珠的叶、茎、根有机碳含量及其比重的双因素方差分析(F值)Table 3 Two-way ANOVA (F-values) of the organic carbon content and its proportion in leaves,stems and roots of R. soongorica and S. passerina in monocultural and mixed conditions under six precipitation gradients
混生后,红砂根和叶有机碳含量均远大于单生,平均高40.71%和45.37%,除50 mm外,在相同降水量下,混生红砂的根部有机碳含量均显著大于单生红砂(图1a,1c);混生后红砂茎有机碳含量小于单生(除300 mm外)(图1b)。混生后,珍珠茎有机碳含量均显著小于单生(P<0.05),平均减少59.51%(图1e);混生后红砂根有机碳含量小于单生(除100 mm),50 mm降水量条件下差异显著(P<0.05),单生比混生高33.98%(图1f);叶有机碳含量大于单生(除300 mm),平均高20.75%(图1d)。
随着降水量的减少,单生红砂叶有机碳含量呈减小趋势,根有机碳含量呈先减小后增加的趋势,在250 mm降水量下达到最小值,与其他降水量下的值差异显著(P<0.05)(图1a,1c);混生红砂的根和茎有机碳含量整体均呈减小趋势,300 mm降水量下的最大值比50 mm降水量下的最小值分别高19.45%和22.09%(图1b,1c)。单生珍珠茎和根有机碳含量随降水量的减少整体呈增加趋势,均在50 mm降水量下达到最大值(图1e,1f);50~150 mm降水范围内,随着降水量的减少,混生珍珠叶有机碳含量逐渐减小,茎有机碳含量逐渐增大(图1 d)。
由图1 g可知,单生红砂有机碳在各器官的分配比重为:茎>根>叶;混生后红砂根有机碳含量逐渐增加,其分配比重为:根>茎>叶。随着降水量的减少,单生红砂根有机碳含量比重呈现先减后增的变化趋势;混生红砂根和茎有机碳含量比重均呈现减-增-减的变化趋势,且在300 mm降水量达到最大值,叶有机碳含量比重呈现增-减-增的变化趋势。由图1 h可知,单生珍珠有机碳的分配比重为:茎>根>叶;混生后珍珠根和茎有机碳含量比重较单生均降低,但仍大于叶,分配比重为根≥茎>叶。单生珍珠叶和混生珍珠茎的有机碳含量比重随着降水量的减少均呈先减后增趋势,混生珍珠叶呈现相反变化趋势(先增后减)。

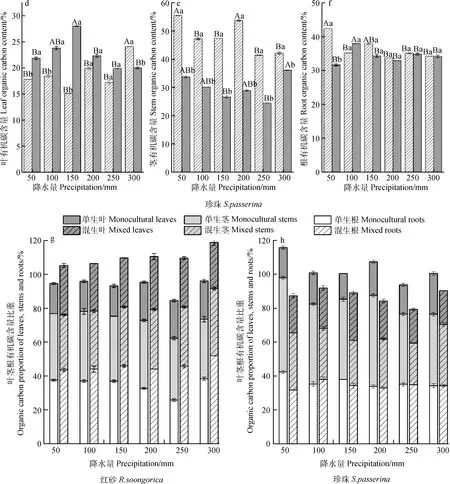
图1 不同降水量下单生、混生红砂珍珠的叶、茎、根有机碳含量及其比重Fig.1 Organic carbon and its proportion in leaves,stems and roots of R. soongorica and S. passerina in monocultural and mixed conditions under six precipitation gradients注:不同大写字母表示相同生长方式下不同降水量间差异显著(P<0.05);不同小写字母表示相同降水量下两种生长方式间差异显著(P<0.05)。下同Note:Different capital letters indicate significant differences in different precipitation under the same growth pattern (P<0.05);different lowercase letters indicate significant differences between the two growth patterns under the same precipitation (P<0.05). The same as below
2.2 不同降水量下叶、茎、根全氮含量及其比重比较
由表4可知,生长方式对红砂根全氮含量和珍珠根、茎的全氮含量及比重存在极显著影响(P<0.01);降水量及其与生长方式的交互作用对珍珠茎、叶的全氮含量有极显著影响(P<0.01),交互作用对珍珠叶全氮含量比重有显著影响(P<0.05)。

表4 不同降水量下单生、混生红砂珍珠的叶、茎、根全氮含量及其比重的双因素方差分析(F值)Table 4 Two-way ANOVA (F-values) of the total nitrogen content and its proportion in leaves,stems,and roots of R. soongorica and S. passerina in monocultural and mixed conditions under six precipitation gradients
混生红砂根全氮含量大于单生(除250 mm降水量外),平均增加23.59%(图2c);混生红砂叶和茎全氮含量与单生相比整体呈减少趋势,分别平均减少4.22%和5.38%,均在200 mm降水量下差异显著(P<0.05)(图2a,2b)。珍珠混生后的根全氮含量显著小于单生(P<0.05),平均减少 32.06%,250 mm降水量下差异最显著,单生比混生高112.88%(图2f);茎全氮含量在50 mm和100 mm降水量下表现为混生小于单生,其余降水量下混生均大于单生,整体上混生比单生平均高3.59%,250 mm降水量下差异最显著(P<0.05),混生比单生高42.82%(图2e)。
随着降水量的减少,单生红砂根和叶的全氮含量均呈现增-减-增的变化趋势(图2a,2c),茎全氮含量呈现逐渐降低趋势,300 mm降水量下的最大值比50 mm降水量下的最小值高52.39%,且差异显著(P<0.05)(图2b);混生红砂叶和茎全氮含量均在250 mm降水量下达到最大值(图2a,2b),根全氮含量在150 mm降水量下达到最大值(图2c)。单生珍珠叶全氮含量在250 mm降水量下达到最大值,与其他降水量下的值均差异显著(P<0.05)(图2 d);在100~300 mm降水范围内,混生珍珠茎和叶的全氮含量随降水量的减少均先增大后减小,在150 mm降水量下达到最大值(图2 d,2e)。
由图2 g可知,单生红砂的全氮主要分配在叶上,其分配比重均为:叶>根>茎,混生后红砂根全氮含量逐渐增加,其分配比重为:根>茎>叶。随降水量的减少,单生红砂叶和根的全氮含量比重均呈现增-减-增的变化趋势,茎全氮含量比重逐渐降低;混生红砂茎和叶全氮含量比重呈现相同变化趋势。由图2 h可知,单生珍珠的全氮分配比重为:根>叶>茎,混生珍珠根全氮比重逐渐减小,其分配比重为:茎>叶>根。混生珍珠茎全氮含量比重随降水量的减少呈现增-减-增的变化趋势,叶和根呈现先增大后减小趋势。
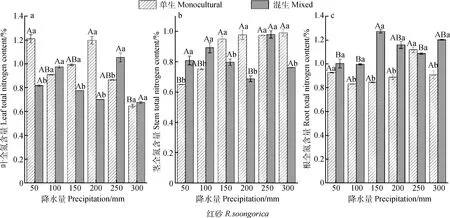
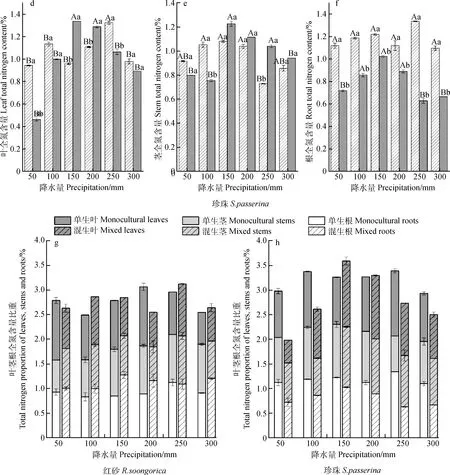
图2 不同降水量下单生、混生红砂珍珠的叶、茎、根全氮含量及其比重Fig. 2 Total nitrogen and its proportion in leaves,stems and roots of R. soongorica and S. passerina in monocultural and mixed conditions under six precipitation gradients
2.3 不同降水量下叶、茎、根碳氮比(C/N)比较
由表5可知,生长方式对红砂叶C/N和珍珠根、茎、叶的C/N均产生极显著影响(P<0.01);降水量及其与生长方式的交互作用对珍珠茎和叶的C/N的影响差异显著(P<0.05)。

表5 不同降水量下单生、混生红砂珍珠的叶、茎、根碳氮比的双因素方差分析(F值)Table 5 Two-way ANOVA (F-values) of the C/N in leaves,stems,and roots of R. soongorica and S. passerina in monocultural and mixed conditions under six precipitation gradients
混生后,红砂根和茎C/N较单生均无明显变化规律;叶C/N比单生增加,平均增长201.99%(图3a,3b)。珍珠混生后,根和叶C/N均高于单生,分别平均增长49.16%和46.08%;茎C/N小于单生,平均减少39.40%(图3c,3 d)。
单生红砂的C/N整体在根和茎上较高,其变化规律为:茎>根>叶;随着降水量的减少,单生红砂根C/N呈现减-增-减的变化趋势,茎C/N整体呈现增长趋势,叶C/N整体呈降低趋势(图3a)。在整个降水梯度范围内,混生红砂茎和叶的C/N随着降水量的减少呈现相同变化趋势;在150~200 mm降水量范围内,混生红砂叶、茎、根的C/N均呈增长趋势,变化规律为茎>叶>根(图3b)。单生珍珠根茎叶C/N分布规律较为明显,其变化为:茎>根>叶(图3c)。混生珍珠根C/N大幅度上升,其变化规律为:根>茎>叶;随着降水量的减少,混生珍珠根茎叶的C/N均整体呈现先减小后增大的变化趋势(图3 d)。

图3 不同降水量下单生、混生红砂珍珠的叶、茎、根碳氮比变化Fig. 3 C/N in leaves,stems and roots of R. soongorica and S. passerina in monocultural and mixed conditions under six precipitation gradients注:不同大写字母表示单生或混生状态下,红砂(珍珠)同一器官C/N在不同降水量之间差异显著(P<0.05);不同小写字母表示单生或混生状态下,红砂(珍珠)在相同降水量条件下不同器官间C/N差异显著(P<0.05)Note:Different capital letters indicate that the C/N of the same organ of R. soongorica (S. passerina) is significantly different in different precipitation level at the 0.05 level under mono-cultural or mixed-cultural growth conditions;different lowercase letters indicate that the C/N of R. soongorica (S. passerina) is significantly different among different organs under the same precipitation level at the 0.05 level
3 讨论
3.1 降水对红砂和珍珠不同器官C,N含量及C/N的影响
作为制约植物生长的主要因素,水分会通过影响植物的光合作用对植物体内的代谢和物质合成等动态平衡过程产生影响,同时影响植物C,N的吸收、分配和转运等过程[37],植物也会通过调控对体内C,N元素的利用以适应水分变化[38]。本研究发现,随降水量的减少,单生红砂叶片有机C含量呈降低趋势(图1a),全N含量无明显的线性变化(图2a),C/N整体呈降低趋势(图3a),这与刘晟彤等[39]对红砂和陈佳瑞等[8]对3种灌木的研究结果一致。可能的原因是:随着干旱胁迫程度的加剧(降水量逐渐减少),红砂幼苗会通过降低蒸腾作用而导致叶片气孔关闭,减小了C同化速率,因此叶片有机C含量降低[40],有机C含量的减小进一步导致叶片C/N降低。可见,红砂在水分限制、营养元素相对匮缺的荒漠生境中,经过长期的进化已形成了自身独特的生长策略和应对环境变化的机制[41-42],当生存环境中水分发生变化后,植株能够迅速对自身状态做出调整,从而与变水环境间形成主要维持在碳吸收和水分流失之间的动态平衡[39]。本研究也发现,随着降水量的减少,单生红砂茎全N含量呈现逐渐降低趋势(图2b),茎C/N整体呈现增长趋势(图3a),这与种培芳等[43]对红砂、王凯等[11]对榆树(Ulmuspumila)和陈佳瑞等[8]对黄刺玫(Rosaxanthina)的研究结果均一致。分析其原因可能是:随着干旱胁迫程度的增强,幼苗茎中逐渐形成栓塞,影响了养分运输过程,导致茎全N含量逐渐减少[44]。同时,由于干旱限制了土壤中参与C,N,P元素循环转化的酶活性减弱导致养分离子的扩散能力减缓[45],可利用N的有效性及矿化速率降低,因此植物能够吸收的N素减少导致N含量降低[13]。红砂茎在干旱胁迫下受N限制作用增强,影响N的吸收及运输,导致茎C/N整体呈现增长趋势[46]。
本研究中,单生珍珠茎和根中有机C含量随降水量的减少整体呈增加趋势(图1e,1f),这与王凯等[11]研究发现随着降水量的减少,榆树幼苗细根有机C含量逐渐升高的结果一致。这可能是因为干旱胁迫加剧导致植物生长受到抑制,生理代谢活动降低,C元素消耗减少,茎作为植物主要的C元素储存器官[47],多余的C会在植物体内积累而后储藏在茎中[44]。根系作为植物体内最先感受到土壤干旱的器官,对环境水分变化的响应极其敏感[48]。随着干旱胁迫程度的加剧,珍珠会选择适当地增加地下根系部分的C分配比例,C作为构成植物骨架的基本元素,能够为植物生理活动提供能源物质[49],从而维持根系的C同化作用[40],保证了干旱环境中植物的存活,体现了荒漠植物珍珠对干旱环境的适应策略。
3.2 混生对红砂和珍珠不同器官C,N含量及C/N的影响
资源竞争过程中,植物对地上光能吸收和地下土壤养分利用的变化会对自身化学计量特征产生影响[50],影响程度与环境胁迫和植物种类等有关[51]。植物不同器官对水分养分的需求存在差异导致不同营养器官间元素含量分配规律具有差异性,这种差异体现了植物对特定环境的适应方式[52]。本研究发现红砂和珍珠混生后,2种植物叶片有机C含量较单生均增加(图1a,1 d),但茎有机C含量比单生减小(图1b,1e),红砂根系中的有机C和全N含量比单生均增加(图1c,2c),而珍珠根系中的有机C和全N含量比单生植株均减小(图1f,2f)。说明红砂和珍珠混生后,2种植物对地上部分有机C的分配策略相同,均通过提高叶片有机C含量来降低光合速率,增强自身对外界胁迫因素的防御和抵抗能力[10]。但由于红砂地下部分根系的固C与固N能力要强于珍珠,两种植物混生后可利用资源的增加促进红砂根系竞争能力进一步加强[53]。红砂根有机C含量的增加进一步导致其根系生物量的增大和可溶性糖等抗性物质的积累增强,因而对胁迫环境的抵御能力也增强[54]。C/N是代表植物生长速率的关键生理指标,一般认为植物器官C/N越低,其生长速率越快[55]。本研究发现,与单生红砂相比,混生后红砂根和茎的C/N变化不大,而叶的C/N显著增大(图3a,3b);与单生珍珠相比,混生珍珠叶的C/N变化不大,而茎的C/N显著降低(图3c,3 d)。这可能是由于混生使红砂将无机物转化为有机物的能力提升,从而促进了植物体内非结构性碳水化合物(糖和淀粉)等物质的积累,最终导致植物体内的N含量被强烈稀释[56-57],所以导致红砂叶片C/N增加。除稀释效应外,可能导致红砂叶片全N含量减少的另一个原因是2种植物混生后,遮阴现象导致红砂叶片的气孔变小甚至关闭减慢了蒸腾速率,N的吸收和利用速度降低,因此植物体内的全N含量降低[28,39]。茎作为连接植物地上地下部分和运输有机物的重要通道,同时也是分配和消耗光合产物的器官[58],珍珠主要通过提高茎对N素的吸收能力来促进地上部分的生长,从而获得能够维持自身生长发育所需的营养,这与种培芳等[59]的研究结果一致。
本研究发现,红砂单生时各器官中有机C和全N含量的分配规律分别为茎>根>叶和叶>根>茎,混生后均为根>茎>叶(图1 g,2 g);而珍珠单生时其全N含量分配规律为根>叶>茎,混生后则为茎>叶>根(图2 h)。这说明红砂在单生时其地上部分光合速率较强,生长速率较快[60],茎作为支撑植物、输导水分和贮藏营养物质的器官,主要由富含C的多糖物质(如木质素、纤维素等)组成[61],因此单生时其茎中有机C含量最高。与珍珠混生后,深根性植物红砂倾向于将更多C,N向根系分配以利于根系向更深层土壤下扎生长[31],从而提高根系对深层土壤中营养物质的吸收利用效率[62],提高自身竞争力。而珍珠在与红砂混生后,主要依靠大气降水生长的浅根性植物珍珠更侧重于将水分养分等营养物质分配到地上部分[63],在有限的养分供应条件下植物的叶片和茎保持较高的N含量,其地上部分具有更强大的获取和充分利用资源的能力。就固N能力而言,地上部分茎叶比地下部分根系要明显强得多,固N能力越强越有益于植物光合作用能力的提高,以完成其正常的生活史[53]。可见,植物不同器官对C,N含量的分配策略存在差异,这可能与植物所处生境资源的可利用性有关,也与植物器官本身的功能特性有关[64-65]。
4 结论
综上所述,在西北荒漠区,红砂和珍珠2种典型荒漠植物均已形成了各自独特的生长策略和应对环境变化的机制。降水量变化和混生生长能够显著改变红砂和珍珠的碳,氮在不同器官中的分配模式。随着干旱胁迫程度的加剧(降水量逐渐减少),红砂茎受氮限制作用增强,影响氮的吸收及运输,导致茎碳/氮整体呈现增长趋势,但红砂会通过降低叶片有机碳含量和碳/氮以适应干旱胁迫;珍珠会选择适当增加地下部分根系的碳分配比例,并且将多余的碳储藏在茎中,保证自身在干旱环境中的存活。混生后,红砂会倾向于向根系分配更多碳,氮来提高对水分养分的竞争能力,而珍珠主要通过降低茎的碳/氮来提高地上部分生长速率,增强自身对外界不利因素的防御能力。
