公路沿线不同植物叶际微生物群落结构及重金属污染比较
2021-09-28周之栋徐建华
周之栋 徐建华

摘要:公路沿线由于汽车尾气和扬尘导致重金属污染严重,道路绿化应兼顾植被对重金属污染物的净化作用。选择公路两侧4种树种为研究对象,即海滨木槿(Hibiscus hamabo)、金边胡秃子(Elaeagnus pungens)、山核桃(Carya cathayensis)和中山杉(Taxodium ‘Zhongshansha )118,测定其叶际微生物群落结构组成和叶际重金属、根际土壤重金属含量。结果显示,不同植物叶际微生物群落结构和物种组成显著不同;中山杉和海滨木槿具有更高的叶际微生物多样性(PD_whole_tree、Shannon、Simpson),而中山杉叶际微生物的丰富度(OTUs、Chao1)最高;中山杉和金边胡秃子叶片Cr(铬)、Cu(铜)、Zn(锌)、Cd(镉)和Pb(铅)含量较低,而对于植被林下土壤重金属含量,海滨木槿较低,中山杉显著高于其余树种;叶片Cr、Cu、Zn以及重金属总量对叶际微生物群落结构具有显著性影响(P<0.05)。综合以上结果,不同植物对不同重金属元素的净化能力不同,其中中山杉总体上对公路交通扬尘阻隔和净化效果更好。
关键词:公路;重金属;汽车尾气;叶际;微生物
中图分类号:X173 文献标志码: A
文章编号:1002-1302(2021)18-0215-07
收稿日期:2020-12-12
基金项目:江苏省中科院植物研究所博士人才科研启动基金(编号:JSPKLB202027)。
作者简介:周之栋(1990—),男,江苏南京人,博士,助理研究员,主要从事微生物生态学研究。E-mail:1289597338@qq.com。
公路交通中汽车汽油、柴油的燃烧尾气,轮胎、刹车片的磨损,道路路面沥青磨损等是公路沿线重金属污染的主要来源[1-2]。这些污染物主要以汽车扬尘的形式进入公路沿线两侧空气和土壤中,其次通过自然沉降、降雨沉降等形式迁移进入路域土壤-植物系统中,给周围农田、居民造成污染[3]。以往关于公路沿线重金属污染的研究主要集中在重金属在路域范围的空间分布格局、植物对汽车扬尘的滞沉效果的研究[4-5]。研究表明,植物对公路交通引起的重金属污染具有很好的阻隔、净化效果,其净化效应主要表现为植物叶片对重金属的吸收、吸附,植物地上部分的直接物理阻挡作用,根系对路域土壤重金属污染物的吸收等形式[6-7]。叶际微生物作为叶片表面参与植物生理生化活动的活跃成分,对植物生理特性具有显著影响,且对环境变化敏感[8],而路域环境下不同植物叶际微生物的群落组成异同以及对植物叶片吸收富集重金属元素影响的研究鲜有涉及。
叶际微生物已被证明与植物叶际微环境变化、促植物生长物质的产生、叶际环境污染物的降解等有密切关系[9]。环境条件的改变会影响叶际微生物的群落结构组成,影响植物形态、生长和生理生态过程,进而影响植物对重金属污染物的吸收富集。叶际微生物可通过吸附、氧化还原重金属以及矿化固结重金属离子来辅助植物叶片对交通粉尘的净化[10]。因此,本研究通过比较公路沿线不同植物叶际微生物群落结构组成、叶片重金属含量和土壤重金属含量,并结合生物统计分析,以期揭示不同植物对公路交通扬尘重金属污染的净化规律及效果。
1 材料与方法
1.1 样品采集
样品于2020年9月采集。采样地点位于江苏省省道S334公路两侧(南通市如皋市搬经镇段,120°23′43″E,32°17′40″N)。选取公路两侧4种主要绿化树种为研究对象,即海滨木槿、金边胡秃子、山核桃和中山杉118(以下文中称为中山杉)。均匀采集植物各部位叶片(约100张),混合作为1个样品,每种树采集3个重复。采集每种植被林下0~20 cm土层土壤,围绕植株取3个点,混合作为1个土壤样品,每种树采集3个重复。将采集的部分叶片样品(用于叶际微生物群落结构测定)置于冰盒中,迅速带回实验室进行后续分析。
1.2 叶片和土壤重金属含量测定
叶片样品置于105 ℃烘箱中杀青2 h,之后 85 ℃ 烘干至恒质量。烘干后的叶片样品研磨粉碎。土壤样品进行风干处理,并研磨过0.149 mm筛。样品处理过程中避免接触金属制品,以避免对重金属测定的影响。样品消解采用微波消解仪(JUPITER,SINEO)。重金属Cr、Cu、Zn、Cd和Pb含量的测定采用电感耦合等离子体质谱仪(ICP-MS,ThermoFisher,USA)。
1.3 叶际微生物测定
取新鲜叶片20 g置于装有100 mL灭菌水的三角瓶中,超声波(40 kHz)振荡处理10 min。转移浸提液于离心管中,离心(11 000 r/min,4 ℃)获取沉淀物用于后续叶际微生物DNA提取。采用E.Z.N.A. Soil DNA Kit (Omega Bio-tek,Norcross,Georgia,USA)試剂盒提取沉淀物中微生物总的DNA。利用琼脂糖凝胶电泳和Qubit 3.0 荧光计(Life Technologies,USA)进行DNA质量检测和定量。针对16S rRNA的V4-V5区域进行PCR(聚合酶链式反应)扩增。所用引物为515F(5′-GTGCCAGCMGCCGCGG-3′)和907R(5′-CCGTCAATTCMTTTRAGTTT-3′)。扩增程序设定为:95 ℃ 2 min;95 ℃ 2 min,55 ℃ 30 s,72 ℃ 30 s,25个循环;最终 72 ℃ 延伸5 min。PCR 20 μL反应体系包括:4 μL 5×FastPfu缓冲液,2 μL 2.5 mmol/L dNTPs,0.8 μL引物(5 μmol/L),0.4 μL FastPfu聚合酶和10 ng模板DNA。扩增子采用2%琼脂糖凝胶电泳进行提取,利用AxyPrep DNA Gel Extraction Kit 试剂盒进行纯化(Axygen Biosciences,Union City,CA,USA),并用Qubit 3.0 荧光计进行定量。测序文库按照标准流程在Illumina HiSeq平台进行双末端测序(2×250)(南京集思慧远生物科技有限公司)。
原始fastq格式测序文件经过QIIME软件(version 1.17)进行多路分解和质量过滤处理。序列处理遵循以下标准:(1)250 bp序列在10 bp 滑窗中的平均质量得分小于20的任意碱基位点处被截断;(2)条码序列的任何错配,引物序列的2个核苷酸的错配以及模糊的序列被剔除;(3)大于10 bp重叠的条带进行组装。采用UPARSE(version 7.1 http://drive5.com/uparse/)依据97%的相似性阈值进行OTU(operational units)划分,采用UCHIME进行嵌合体序列的识别和去除。利用uclust程序对比于silva数据库对16S rRNA序列进行系统发育关系归类,置信阈值控制在50%。
1.4 统计分析
物种数量稀释性曲线采用Mothur软件分析。叶际微生物多样性指数Chao1、PD_whole_tree、Shannon、Simpson采用R语言(vegan包)软件进行计算。Venn图、柱状图、热图、聚类分析、主坐标分析(基于weighted unifrac距离)、冗余分析采用R语言绘制(vegan包和ggplot2包)。采用Adonis算法对比不同样本间叶际微生物群落结构的差异。采用单因素方差分析结合Duncans多重分析比较不同样本间重金属含量的统计学差异。LEfSe分析采用网页分析器http://huttenhower.sph.harvard.edu/galaxy,以LDA=3判别分值作为划分各样本特有生物标记物(biomarker)的阈值。
2 结果与分析
2.1 不同植物叶际微生物群落结构组成
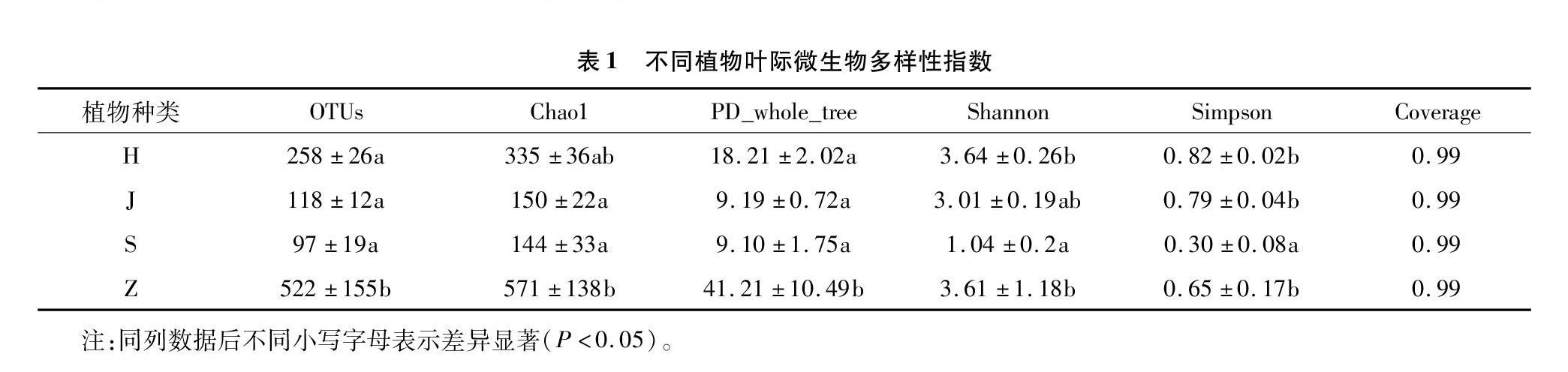
微生物种类数量稀释性曲线随测序深度增加趋于平缓,测序深度达到0.99,表明本试验测序量足够支持后续分析(图1)。中山杉和海滨木槿叶际微生物多样性较高,各多样性指数较之于其他树种分别高出98.15%~352.85%(PD_whole_tree)、19.93%~250.00%(Shannon)、116.67%~173.33%(Simpson);其中中山杉具有最大的物种丰富度指数,OTUs指数高于其他树种102.33%~438.14%、Chao1指数高出70.45%~296.53%(表1)。不同树种间叶际微生物物种组成差异较大,其中中山杉具有最多的特有微生物种类(557个OTU),其次为海滨木槿(38个OTU)(图2)。
在微生物门水平上,不同树种变形菌门(Proteobacteria)占比最大(52.02%~99.50%)。在門水平上,山核桃具有较丰富的Proteobacteria,海滨木槿具有较丰富的蓝藻门(Cyanobacteria)、放线菌门(Actinobacteria),金边胡秃子具有较丰富的厚壁菌门(Firmicutes),中山杉具有较丰富的拟杆菌门(Bacteroidetes)、Actinobacteria、酸杆菌门(Acidobacteria)、浮霉菌门(Planctomycetes)、绿弯菌门(Chloroflexi)、疣微菌门(Verrucomicrobia)(图3-a)。在属水平上,山核桃具有较丰富的泛菌属(Pantoea)、乳球菌属(Lactococcus),海滨木槿具有较丰富的马赛菌属(Massilia)、假单胞菌属(Pseudomonas)、鞘氨醇单胞菌属(Sphingomonas)、不动杆菌属(Acinetobacter)、短小杆菌属(Curtobacterium),金边胡秃子具有较丰富的狭义梭菌属1(Clostridium sensu stricto 1)、Kosakonia、芽孢杆菌属(Bacillus)、狭义梭菌属5(Clostridium sensu stricto 5),中山杉具有较丰富的Sphingomonas、Curvibacter、Hafnia-Obesumbacterium、Methylobacterium(图3-b)。不同植物叶际微生物群落结构组成具有显著差异(P<0.05)(图3-c)。
LEfSe分析显示不同植物的叶际微生物具有各自特有的生物标记物,即特有的丰富物种(图4)。中山杉和海滨木槿具有最多的生物标记物,分别为39和32个。山核桃生物标记物为Proteobacteria,海滨木槿生物标记物主要为Actinobacteria、Oxyphotobacteria、Micrococcales、Chloroplast、Acetobacterales、Caulobacterales、立克次氏体目(Rickettsiales)、假单胞菌目(Pseudomonadales)、Xanthomonadales,金边胡秃子生物标记物为Clostridia、Clostridiales、Clostridiaceae 1、芽孢杆菌目(Bacillales)、芽孢杆菌科(Bacillaceae),中山杉生物标记物主要是Actinobacteria、Bacteroidetes、Chloroflexi、拟杆菌纲(Bacteroidia)、Alphaproteobacteria、Deltaproteobacteria、Planctomycetacia、Propionibacteriales、Micromonosporales、Kineosporiales、Corynebacteriales、Cytophagales、Sphingomonadales、红细菌目(Rhodobacterales)、Rhizobiales、Myxococcales。
2.2 叶片和土壤重金属含量
不同植物叶片重金属含量存在显著差异(图5)。海滨木槿叶片对Cr、Cu、Pb的富集能力较强;山核桃对Zn、Cd、Pb的富集较多,中山杉和金边胡秃子对几种重金属的叶片富集能力较弱。对于几种重金属的整体富集能力,海滨木槿和山核桃表现更好。
不同植物林下土壤重金属含量表现出显著性差异(图6)。中山杉林下土壤重金属含量较高,显著高于其他树种。海滨木槿林下土壤重金属含量最低。山核桃和金边胡秃子林下土壤重金属含量无显著性差异。
对土壤和植物叶片重金属浓度进行比值计算,除Pb之外,其余重金属浓度比值集中在0.76~4.69(表2)。海滨木槿和山核桃比值较低,中山杉和金边胡秃子比值较高。通过比值计算,说明植物通过叶片截留和沉降是公路沿线两侧树木对汽车尾气扬尘污染阻隔的主要途径。
2.3 重金属含量与微生物群落结构组成的相关性分析
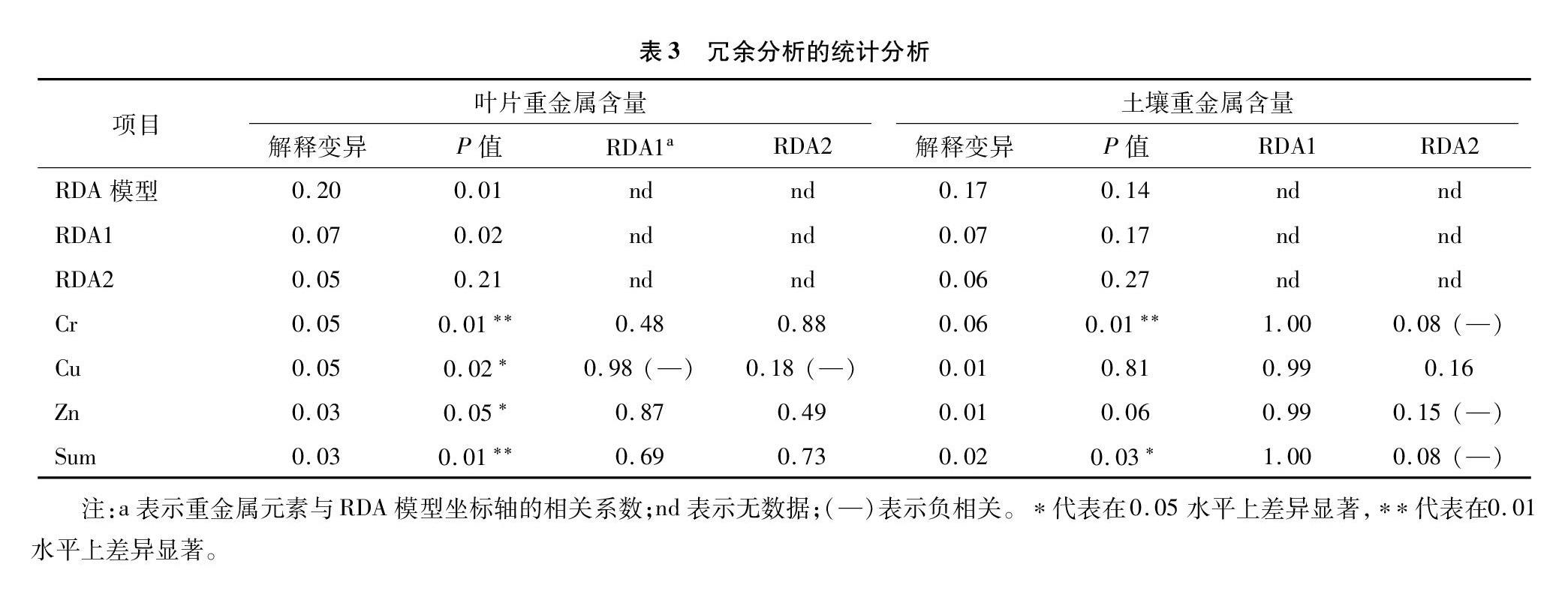
冗余分析模型(RDA)前2个坐标轴解释了63.63%(叶片重金属)和74.74%(土壤重金属)的变异量,说明该模型可以很好地解释微生物群落结构与重金属含量之间的相关关系。在OTU水平上的冗余分析表明,叶片重金属Cr、Cu、Zn、Sum对叶际微生物群落结构具有显著性影响(图7,表3)。土壤重金属含量与叶际微生物群落结构的相关性较弱。RDA模型将不同植物的叶际微生物群落显著区分开。分析重金属含量与RDA坐标轴的相关性,叶片重金属Cu和Pb含量与Cr、Cd、Zn和Sum含量之间呈现负相关性;土壤重金属含量之间呈现正相关性。RDA模型中OTU1、OTU2、OTU3、OTU7、OTU9、OTU55(Proteobacteria),OTU4(Cyanobacteria),OTU5、OTU6、OTU8、OTU11(Firmicutes)与叶际微生物群落结构和重金属含量具有较强的相关性。
3 讨论与结论
由物种数量稀释性曲线结合测序深度(Coverage)可知测序条带数量已经充分覆盖叶际微生物种类。稀释性曲线结果与Chao1丰富度指数一致,即中山杉具有较高的物种丰富度,而山核桃最少,这说明中山杉作为针叶乔木树种,相比于其他阔叶乔木及灌木,其叶际具有更为复杂的微生物群落结构组成。中山杉叶际相较于其余树种具有极为丰富的特有微生物物种组成,如venn图分析显示其具有557种特有OTU,LEfSe分析其特有生物标记物为39种,这些指标均远高于其余树种。分析可能的原因是中山杉叶片特有的分泌物或叶面结构[11],提供给环境中微生物适宜的生存场所,从而招募更多样化的微生物类群寄居在其叶片表面,这有待于后续更深入探索其中的机理。根据微生物对重金属净化机理的研究,本研究分析中山杉叶际多样化的微生物种类对公路扬尘中的重金属元素进行初步的生化反应转化,一部分重金属元素固持在微生物体内,一部分重金属元素活化后可增加叶片对其的吸收积累[12-14]。叶际微生物与植物叶片重金属的吸附作用的显著关系通过冗余分析结果也得以体现,这进一步说明叶际微生物在参与植物净化路域重金属污染中的作用。微生物类群Proteobacteria作为不同树种叶际微生物的优势菌群,反映出其对环境的较广的适应能力[9,15],本研究中RDA模型同样发现叶片和土壤重金属元素含量与该微生物类群的显著相关关系,说明该微生物种群在叶际微环境中发挥了重要作用。在微生物属水平上的优势类群Pantoea、Massilia、Pseudomonas也隶属于该菌类。
公路交通带来的汽车尾气、部件摩擦等产生的携带重金属颗粒的扬尘是引起沿线路域重金属污染的主要来源。扬尘经过自然沉降或降雨等形式沉降到植物叶片表面和林下土壤中,因此作为直接暴露在此大气扬尘污染环境中的植物叶片,其直接对重金属的吸附是净化重金属的重要途径[16-18]。植物从土壤中吸收重金属主要集中于根部,而短时间内从土壤到叶片的富集量较少,因此路域植物的叶片直接吸附吸收重金属是一条重要途径[19-20]。本研究發现,路域叶片重金属浓度与土壤重金属含量比值集中在0.76~4.69(Pb除外,这说明植物对Pb吸附净化能力较之其余元素弱),这进一步说明由大气环境中扬尘带来的重金属污染通过叶片的吸附、吸收。本研究还发现,中山杉叶片中重金属含量相比于其他树种浓度含量较低,可能解释是多样性的中山杉叶际微生物通过吸收固定重金属粉尘阻隔了进一步进入中山杉叶片内部。通过RDA模型发现,叶片重金属含量(Cr、Cu、Zn和Sum)与叶际微生物群落结构具有显著相关关系。本研究未考虑以浓度进行比较的局限性。之前研究也从叶片的组织结构、分泌物以及叶型等角度探讨叶面对空气中粉尘污染物附着以及吸收的影响[21-22],但也有研究讨论了降雨以及风力作用可以比较容易移走大部分聚集在叶面的附着粉尘从而使其最终沉降到林下土壤[16]。因此,考量公路绿化植被下土壤中重金属元素的含量,可以从一个侧面反映植物对扬尘的阻隔作用。中山杉林下土壤重金属含量较高,说明其较强的扬尘阻隔能力,将交通扬尘阻隔于林下土壤中,阻止其进一步向公路两侧农田和居住区的扩散。这可能与中山杉树形有关:其植株呈宝塔形,主干下部萌生较多枝条,加之叶型为针叶且茂密,可以很好地遮挡交通扬尘。作为灌木的海滨木槿和金边胡秃子可能由于树形低矮,而山核桃枝叶稀疏,致使它们对扬尘的阻隔效应并不明显。
参考文献:
[1]杜孔昌,张景光,冯 丽. 路域土壤-植物系统重金属污染研究综述[J]. 中国沙漠,2014,34(6):1598-1604.
[2]Al-Taani A A,Nazzal Y,Howari F M. Assessment of heavy metals in roadside dust along the Abu Dhabi-Al Ain National Highway,UAE[J]. Environmental Earth Sciences,2019,78(14):1-13.
[3]Huber M,Welker A,Helmreich B. Critical review of heavy metal pollution of traffic area runoff:Occurrence,influencing factors,and partitioning[J]. Science of the Total Environment,2016,541:895-919.
[4]智颖飙,王再岚,王中生,等. 公路绿化植物油松(Pinus tabulaeformis)和小叶杨(Populus simonii)对重金属元素的吸收与积累[J]. 生态学报,2007,27(5):1863-1872.
[5]王再岚,何 江,智颖飙,等. 公路旁侧土壤植物系统中的重金属分布特征[J]. 南京林业大学学报(自然科学版),2006,30(4):15-20.
[6]Papa S,Bartoli G,Nacca F,et al. Trace metals,peroxidase activity,PAHs contents and ecophysiological changes in Quercus ilex leaves in the urban area of Caserta (Italy)[J]. Journal of Environmental Management,2012,113:501-509.
[7]冯效毅,刘春阳. 公路周边土壤重金属污染及植物修复[J]. 污染防治技术,2011,24(5):17-21,66.
[8]Shakir S,Zaidi S S,de Vries F T,et al. Plant genetic networks shaping phyllosphere microbial community[J]. Trends in Genetics,2021,37(4):306-316.
[9]韓筱璇. 庞庄矿区飞灰对小麦幼苗及其叶际微环境的影响研究[D]. 徐州:中国矿业大学,2019:1-6.
[10]Rahman Z. An overview on heavy metal resistant microorganisms for simultaneous treatment of multiple chemical pollutants at co-contaminated sites,and their multipurpose application[J]. Journal of Hazardous Materials,2020,396:122682.
[11]Kniskern J M,Traw M B,Bergelson J. Salicylic acid and jasmonic acid signaling defense pathways reduce natural bacterial diversity on Arabidopsis thaliana[J]. Molecular Plant-Microbe Interactions,2007,20(12):1512-1522.
[12]钱春香,王明明,许燕波. 土壤重金属污染现状及微生物修复技术研究进展[J]. 东南大学学报(自然科学版),2013,43(3):669-674.
[13]Yin K,Wang Q N,Lv M,et al. Microorganism remediation strategies towards heavy metals[J]. Chemical Engineering Journal,2019,360:1553-1563.
[14]王瑞宏. 铜尾矿区白羊草叶际和根际微生物群落特征研究[D]. 太原:山西大学,2019:1-5.
[15]Dang C C,Xie G J,Liu B F,et al. Heavy metal reduction coupled to methane oxidation:Mechanisms,recent advances and future perspectives[J]. Journal of Hazardous Materials,2021,405:124076.
[16]张 军,梁青芳,高 煜. 宝鸡市绿植叶片重金属空间分布及污染特征[J]. 环境科学,2020,41(10):4504-4513.
[17]林星宇,李海梅,李彦华,等. 灌木滞尘能力与重金属含量间的关系[J]. 江苏农业科学,2019,47(15):180-183.
[18]林星宇,李海梅,李彦华,等. 8种乔木滞尘量与重金属含量的关系[J]. 林业资源管理,2018(5):63-69.
[19]杨淏舟,李艳梅,陈奇伯,等. 昆明市区18种常见绿化树种叶片重金属富集特征[J]. 福建农林大学学报(自然科学版),2017,46(5):584-589.
[20]李 果. 浙江农林大学衣锦校区绿化植物滞尘效应分析[D]. 杭州:浙江农林大学,2019:1-12.
[21]Perini K,Ottelé M,Giulini S,et al. Quantification of fine dust deposition on different plant species in a vertical greening system[J]. Ecological Engineering,2017,100:268-276.
[22]Han D H,Shen H L,Duan W B,et al. A review on particulate matter removal capacity by urban forests at different scales[J]. Urban Forestry & Urban Greening,2020,48:126565.
