伊拉肉兔和美系獭兔FGF5基因外显子及部分内含子多态性研究
2021-09-14牛华锋张成东杨雪娇任战军王淑辉赵永攀杨凌职业技术学院动物工程分院陕西杨凌700西北农林科技大学动物科技学院陕西杨凌7000陕西省畜牧产业试验示范中心陕西泾阳770
牛华锋,张成东,杨雪娇,任战军,王淑辉,赵永攀(.杨凌职业技术学院 动物工程分院,陕西 杨凌,700;. 西北农林科技大学 动物科技学院,陕西 杨凌,7000; . 陕西省畜牧产业试验示范中心,陕西 泾阳770)
兔毛生长发育过程与毛囊(毛乳头)关系密切[1]。成纤维细胞生长因子5(Fibroblast growth factor 5,FGF5)是唯一通过影响毛囊生长期的长短而影响被毛长度的调控因子[2]。因此,研究该基因的序列结构和遗传分布对家兔被毛性状的选育具有重要意义。FGF广泛存在于各组织中,迄今为止共鉴定包括FGF5在内的23种FGFs。Hebert等[3]1994年利用基因打靶技术,通过胚胎干细胞基因敲除证实go基因就是FGF5基因。Sundberg等[4]进一步揭示,FGF5基因大约2 kb核苷酸的缺失导致安哥拉鼠躯干被毛过度增长。Mulsant等[5]于2003年扩增出兔FGF5外显子Ⅰ和Ⅲ,发现外显子Ⅰ存在一个TCT插入突变,外显子Ⅲ存在T-C错义突变。夏胜荣等[6]采用PCR-RFLP法 对5个家兔群体FGF5基因部分外显子1的遗传多样性分析发现外显子1的220~222位点存在TCT缺失。冯凯等[7]分析了5个家兔品种FGF5 CDS区序列,发现外显子Ⅰ285~287位点存在TCT缺失,外显子Ⅲ存在T-C突变,但是没有得到外显子Ⅲ的基因型条带。李春笑等[8]采用PCR产物直接测序法检测荥经长毛兔、天府黑兔以及加利福尼亚兔,发现FGF5外显子Ⅰ的217位点存在TCT插入突变,检测出2种基因型,FGF5外显子Ⅲ的59位点和3位点存在T-C错义突变和T-C同义突变。由此可见,对家兔FGF5基因的多态性研究还不尽完善,不同兔品种间FGF5基因多态性存在差异,有必要对更多的兔品种进行多态性研究,为家兔分子育种提供更多的理论依据。
伊拉肉兔是法国欧洲兔业公司在20世纪70年代末由9个原始品种经不同杂交组合选育而成,具有生长发育快、出肉率高、肉质鲜嫩,但被毛品质差。美系獭兔是从美国引进,被毛品质好,粗毛率低,被毛密度大,制裘价值很高。本试验旨在研究伊拉肉兔和美系獭兔FGF5基因外显子Ⅰ、外显子Ⅱ和外显子Ⅲ的单核苷酸多态性,寻找有价值的多态位点以及突变位点,探索与毛质性状有关的遗传标记,为家兔分子选育提供理论依据。
1 材料与方法
1.1 试验动物
在陕西天鑫食品有限公司选择伊拉肉兔和美系獭兔各70只(公母各35只),心脏采血10 mL放入已添加2.5 mL灭菌ACD抗凝剂的离心管中,用冰盒迅速带回实验室,-80 ℃保存。采用酚-氯仿法提取基因组DNA,-20 ℃保存。
1.2 试验方法
1.2.1 引物设计 参考http://asia.ensembl.org/ 基因库中兔FGF5基因外显子Ⅰ、Ⅱ和Ⅲ序列,用oligo 6设计引物(表1)。引物由生工生物工程(上海)股份有限公司合成。
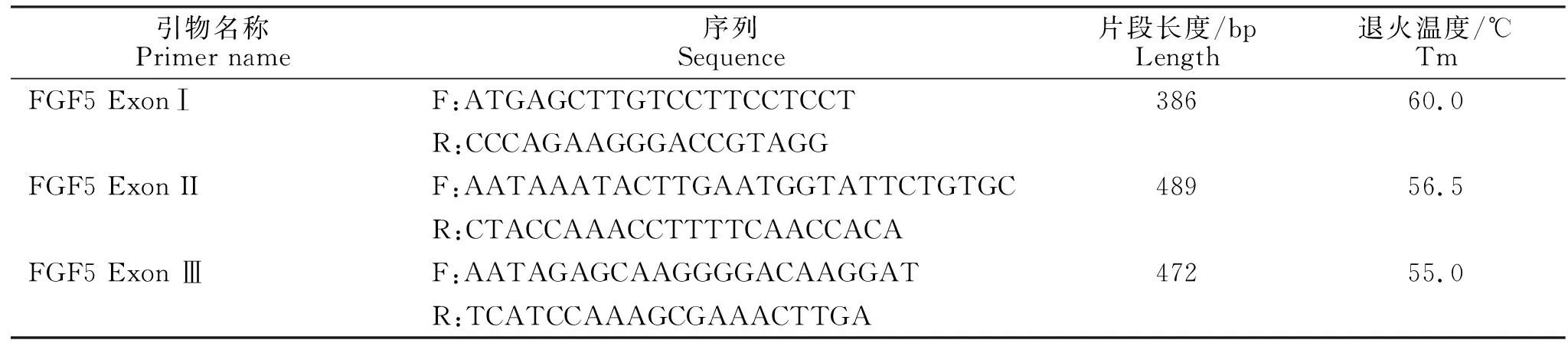
表1 FGF5基因外显子Ⅰ、Ⅱ和Ⅲ的引物序列信息Table 1 Primer sequence information of FGF5 ExonⅠ, Ⅱ and Ⅲ
1.2.2 PCR扩增条件 PCR体系为50 μL : 2×Taq Master Mix来自康威世纪,由Taq DNA polymerase、PCR Buffer、Mg2+、dNTPs、蓝色染料以及PCR稳定剂和增强剂组成,共25 μL;上游和下游引物各2 μL(浓度为10 pmol/μL),DNA模板1μL(浓度为100 ng/μL),dd H2O 20 μL。PCR扩增程序为:95 ℃预变性5 min;94 ℃变性30 s,FGF5基因 ExonⅠ、Ⅱ和Ⅲ的退火分别为60 ℃、56.5 ℃、 55 ℃,均为40 s,72 ℃延伸40 s,34个循环;72 ℃延伸5 min,4 ℃保存。取5 μL扩增产物用1.5%的琼脂糖凝胶电泳检测,并用BIO-RAD凝胶成像系统检测扩增结果。
1.2.3 PCR产物测序 各样本均取30 μL扩增产物和20 μL上下游引物,送往生工生物工程(上海)股份有限公司进行纯化、反向测序。
1.2.4 数据统计 采用MegAlign、Chromas Application 2.3.0.0软件对测序结果进行序列结构分析,通过序列比对和序列峰值校正,筛选出SNP位点,判断突变位点的基因型。用PopGene32软件进行Hardy-Weinberg平衡检验,计算基因型频率、等位基因频率、杂合度、有效等位基因数等遗传参数,用PIC软件计算多态信息含量。
2 结果与分析
2.1 PCR扩增结果
FGF5外显子Ⅰ、Ⅱ和Ⅲ的PCR产物琼脂糖凝胶电泳结果见图1,产物特异性良好,片段长度符合预期。
图1 FGF5 外显子Ⅰ(A)、Ⅱ(B)和Ⅲ(C) PCR产物电泳检测结果 M. D2000标准分子量;1~5. FGF5外显子ⅠPCR产物;6~10. FGF5外显子ⅡPCR产物;11~15. FGF5外显子ⅢPCR产物Fig.1 Electrophoresis results of FGF5 Exon Ⅰ (A) ,Ⅱ (B) and Ⅲ (C) PCR amplification products M. D2000 DNA ladder;1~5. products of FGF5 Exon Ⅰ; 6~10. products of FGF5 Exon Ⅱ;11~15. products of FGF5 Exon Ⅲ
2.2 PCR产物测序结果
将家兔FGF5外显子Ⅰ、Ⅱ和Ⅲ的PCR产物送往生工生物工程(上海)股份有限公司进行纯化、双向测序。结果显示FGF5外显子Ⅰ和Ⅲ均存在多态性,FGF5外显子Ⅱ不存在多态性。
2.2.1FGF5外显子Ⅰ 测序结果发现,在FGF5外显子Ⅰ的168~169位点间(记作FGF5外显子ⅠA位点)存在AGA 3碱基插入突变,将该位点存在AGA插入的等位基因命名为A1,不存在AGA插入的等位基因命名为A2,共发现3种基因型:基因型A1A1(突变型)为引物169~171位点存在AGA 3碱基插入(反向);基因型A2A2(野生型)为 169位点不存在AGA 3碱基插入(反向);基因型A1A2(杂合子)为 169位点等位基因存在AGA 3碱基插入造成测序结果产生移码突变(反向)(见图2)。
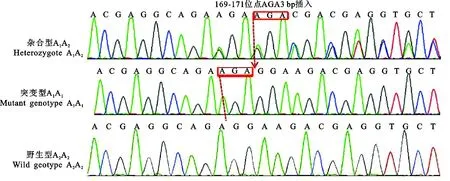
图2 FGF5外显子Ⅰ的3种基因型Fig.2 Three genotypes of FGF5 Exon I
2.2.2FGF5外显子ⅡFGF5外显子Ⅱ的测序结果显示,外显子Ⅱ全部序列(104 bp)以及内含子Ⅰ的7 686~7 852 bp和内含子Ⅱ的1~218 bp片段未发现多态位点(图3)。

图3 外显子Ⅱ基因序列(反向)Fig.3 Exon II gene sequence(Reverse)
2.2.3FGF5外显子ⅢFGF5外显子Ⅲ的135位点(记作FGF5外显子ⅢB位点)存在C-G 单碱基突变,将含C碱基的等位基因命名为B1,含G碱基的等位基因命名为B2,共检测出2种基因型,分别为B1B1和B1B2,见图4。

图4 FGF5外显子Ⅲ 两种基因型Fig.4 Two genotypes of FGF5 Exon Ⅲ
2.3 遗传多态性分析
由表2可知,在所检测的伊拉肉兔和美系獭兔群体中,FGF5外显子ⅠA位点均存在3种基因型,在伊拉肉兔中A2A2基因型频率明显高于A1A1和A1A2基因型频率,A2A2基因型为优势基因型;A2等位基因频率明显高于A1等位基因频率,等位基因A2为优势等位基因。在美系獭兔中A1A1基因型频率明显高于A1A2和A2A2基因型频率,A1A1基因型为优势基因型;A1等位基因频率明显高于A2等位基因频率,等位基因A1为优势等位基因。FGF5外显子ⅠA位点基因型频率在伊拉肉兔群体中分布不符合hardy-weinberg平衡定律(P<0.05),在美系獭兔群体中分布符合Hardy-Weinberg平衡定律(P>0.05)。
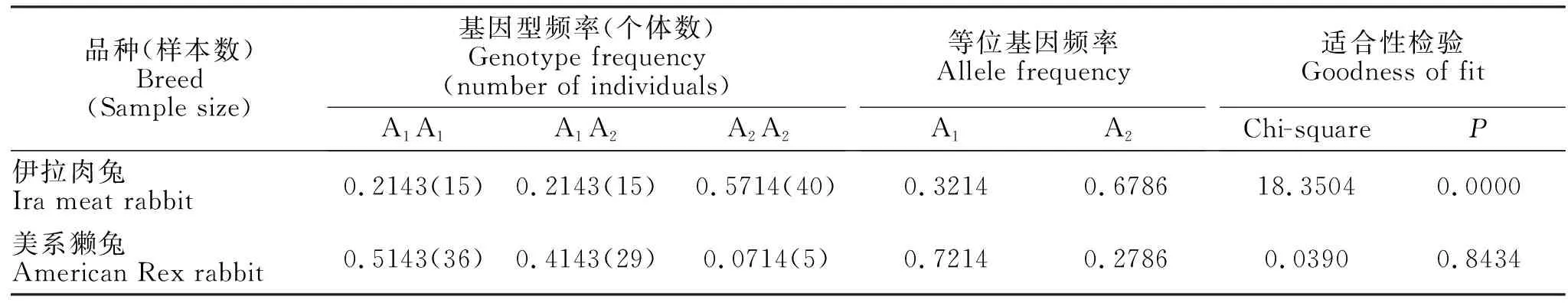
表2 FGF5外显子Ⅰ不同基因型在伊拉肉兔和美系獭兔中的遗传分布Table 2 Genetic distribution of FGF5 Exon I genotypes in Ira meat rabbit and American Rex rabbit
由表3可知,在所检测的伊拉肉兔和美系獭兔群体中,FGF5外显子ⅢB位点都只存在B1B1和B1B2两种基因型,且基因型B1B1的频率分别为0.6429和0.8857均明显高于B1B2基因型频率,基因型B1B1为优势基因型;B1等位基因频率明显高于B2等位基因频率,等位基因B1为优势基因。FGF5外显子ⅢB位点基因型频率在伊拉肉兔和美系獭兔群体中分布均符合Hardy-Weinberg平衡定律(P>0.05)。

表3 FGF5外显子Ⅲ不同基因型在伊拉肉兔和美系獭兔中的遗传分布Table 3 Genetic distribution of FGF5 exon Ⅲ genotypes in Ira meat rabbit and Amercian Rex rabbit
杂合度和多态信息含量是衡量群体等位基因片段多态性的重要参数,杂合度和多态信息含量越低,表明基因突变变异程度越低,遗传一致性越高。由表4可知,伊拉肉兔和美系獭兔FGF5外显子Ⅲ的B位点观测杂合度,期望杂合度和有效基因数均较低,而它们FGF5外显子Ⅰ的A位点这些指标相对较高。在伊拉肉兔和美系獭兔FGF5外显子Ⅰ的A位点以及伊拉肉兔FGF5外显子Ⅲ的B位点均存在中度多态(0.25 表4 伊拉肉兔和美系獭兔FGF5基因各多态位点的遗传参数Table 4 Genetic parameters of FGF5 gene polymorphisms in Ira meat rabbit and American Rex rabbit 本试验采用PCR产物直接测序法,对伊拉肉兔和美系獭兔两个品种进行基因多态性分析,与PCR-SSCP法和PCR-RFLP法相比,不仅简化了试验操作,而且提高了对PCR产物突变位点的检测精度,PCR产物直接测序法寻找突变位点的准确率达到了100%。 伊拉肉兔和美系獭兔是目前我国饲养量较多的家兔品种,美系獭兔毛绒品质优良,被毛品质好,粗毛率低,被毛密度较大,皮板富有弹性;伊拉肉兔被毛品质与美系獭兔差异较大,因此,本研究选择这2个品种分析研究其毛绒品质相关分子遗传标记。本研究发现在伊拉肉兔和美系獭兔2群体中FGF5基因外显子Ⅰ和Ⅲ均存在多态性,FGF5外显子Ⅱ不存在多态性。在FGF5外显子Ⅰ中存在1个TCT插入突变,与Mulsant等[5]和李春笑等[8]的研究结果一致,但是各基因型在不同兔品种间的分布有所不同,本研究结果显示A1等位基因为美系獭兔的优势基因,A2等位基因为伊拉肉兔的优势基因,而冯凯等[7]发现A1等位基因为闽西南兔、海狸色獭兔、白色獭兔、九疑山兔和皖江长毛兔的优势基因,皖系长毛兔群体中未发现A2等位基因;李春笑等[8]研究显示A1等位基因在荥经长毛兔中占绝对优势,A2等位基因在天府黑、加利福尼亚兔中占绝对优势。刘琳玲等[9]研究发现FGF5基因T376C位点为水貂毛长性状的主控位点。FGF5在哺乳动物毛发生长过程中具备关键性的调节作用,FGF5基因外显子Ⅰ发现的A1等位基因和A2等位基因在伊拉肉兔和美系獭兔的差异性与品种选育有关,美系獭兔在培育的过程中绒毛品质是关键性状,以皮用为主,伊拉肉兔在培育过程中肉用性能为关键性状,兼顾绒毛品质。由此可以证明FGF5外显子ⅠA位点A1等位基因决定了家兔的长毛的表型,是影响被毛生长的重要因子,对家兔品种培育及绒毛资源利用都具有极高的应用价值。 FGF5基因是多态信息含量是衡量等位基因片段多态性的理想指标,是衡量基因突变变异程度高低的指标[10], 本研究伊拉肉兔和美系獭兔FGF5外显子Ⅰ的A位点以及伊拉肉兔外显子Ⅲ的B位点多态信息含量0.25 FGF5外显子ⅢB位点基因型频率在伊拉肉兔和美系獭兔群体中均处于Hardy-Weinberg平衡状态(P>0.05)。FGF5外显子ⅠA位点基因型频率在美系獭兔群体中也处于Hardy-Weinberg平衡状态(P>0.05),而在在伊拉肉兔群体中处于Hardy-Weinberg非平衡状态(P<0.05)。高爱琴[11]研究发现FGF5外显子SNP位点 在 内 蒙古绒山羊、辽宁绒山羊品的基因频率处于Hardy-Weinberg不平衡状态,而在文登奶山羊群体处于Hardy-Weinberg 平衡状态。李新海等[12]研究内蒙古绒山羊在该位点基因频率均处于Hardy-Weinberg平衡状态。美系獭兔在FGF5外显子ⅠA位点和外显子ⅢB位点基因频率处于平衡状态的原因说明其群体在品种选育、遗传漂变等作用下,这些位点基因均处于动态平衡中。造成伊拉肉兔FGF5外显子ⅠA位点基因频率不平衡的原因可能与其对经济性状的选择、样本数量等因素有关,其对毛绒品质的调控还需进一步的研究和探索。 本研究发现FGF5外显子Ⅲ引物的135位点存在G-C的突变,但未发现冯凯等[7]、李春笑等[8]、Mulsant等[5]所发现的T-C错义突变。对于突变是否影响以及如何影响家兔的产毛性能还需要进一步的研究。 家兔作为一种重要的皮用经济动物,毛绒品质至关重要,为了提高其生产性能,本研究对伊拉肉兔和美系獭兔2品种FGF5基因外显子及部分内含子多态性研究发现在伊拉肉兔和美系獭兔2群体中FGF5基因外显子Ⅰ和Ⅲ均存在多态性,FGF5外显子Ⅱ不存在多态性;FGF5外显子ⅠA位点A1等位基因为美系獭兔的优势基因,A2等位基因为伊拉肉兔的优势基因。由此推断FGF5可以作为调控家兔毛绒品质的候选基因,FGF5外显子ⅠA位点A1等位基因是影响被毛生长的重要因子,对家兔品种培育及绒毛资源利用都具有极高的应用价值。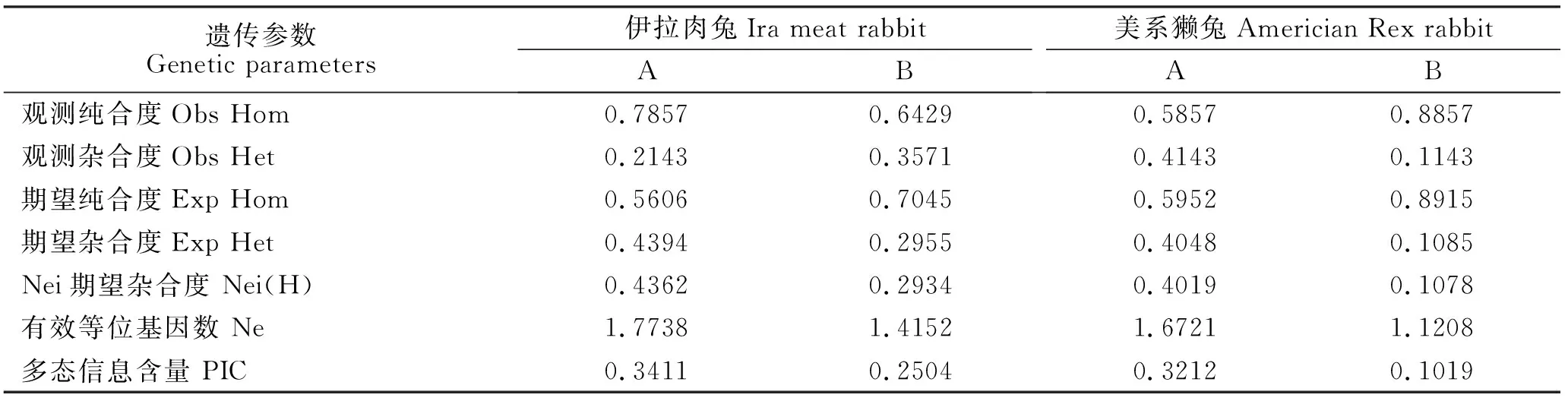
3 讨 论
4 结 论
