有机肥对退化红壤中细菌群落功能组成影响的PICRUSt基因预测分析
2021-09-11吴照祥刘巧丽李辉虎钟永达叶昌炎余发新
吴照祥 刘巧丽 李辉虎 钟永达 叶昌炎 余发新
摘要:为探讨不同修复措施对南方典型退化红壤细菌群落功能组成的影响,设置单施生物炭(B)、微生物有机肥(OF)、化学肥料(CF)、生物炭配施微生物有机肥(BO)、生物炭配施化学肥料(BC)以及空白对照(CK)等6种修复处理方式并进行车前草的盆栽试验,采用细菌16S rRNA基因的高通量测序技术并结合PICRUSt功能预测分析,研究不同修复处理方式下退化红壤细菌群落结构及其功能组成的变化。结果表明,微生物有机肥和化学肥料对车前草根际土壤细菌群落结构与功能组成具有显著的影响(F2,15=25.55,R2=0.773 1,P<0.001;F2,15=17.22,R2=0.696 6,P<0.001),而生物炭的作用不显著(P>0.05);微生物有机肥处理车前草根际土壤细菌物种多样性显著增加,增幅达到17.30%,但是化学肥料显著降低细菌物种多样性,降幅达21.25%;微生物有机肥增加生物代谢途径和氮循环途径功能基因丰度,促进氮异化还原、反硝化作用和固氮作用过程,有利于土壤氮素的维持和土壤质量的改善,而化学肥料施用则相反。另外,根际土壤细菌物种多样性跟土壤的pH值显著正相关,跟碱解氮和有效磷等土壤养分含量显著负相关。本研究明确了退化红壤微生物群落功能组成以及氮循环相关功能基因对修复措施的响应,为南方退化红壤的生态修复与农业生产提供理论依据。
关键词:PICRUSt;退化红壤;生态修复;细菌群落;功能基因预测
中图分类号:S154.38+1 文献标志码: A 文章编号:1002-1302(2021)16-0060-07
土壤退化是一个全球性的问题,主要包括土壤盐渍化、酸化、板结、重金属污染、营养元素失衡、有机质和微生物多样性减少等。在过去40年里,世界上1/3的可耕地因为各种退化原因而无法耕种。红壤是我国南方土壤的重要组成部分,占全国土地面积的21%以上[1]。江西省地处南方亚热带中部,是我国重要的红壤分布区,全省71%的土地面积都是红壤[2]。南方红壤地区水、热条件优越,光照充足,是我国农林业生产的重要区域,其中福建省和江西省森林覆盖率位列全国之首,但是早期森林的过度采伐、毁林垦地,以及过度放牧、化肥农药的过度施用和高强度多频次耕作等不合理的土地利用[3],导致红壤肥力下降、水土流失、地力衰退等严重的土壤退化问题,也引发了一系列的民生、经济、社会和生态问题。
土壤退化问题自1971年由联合国粮农组织(FAO)提出以来,一直备受各界的广泛关注,世界各国也都开展了土壤退化现状、成因以及应对措施方面的研究。我国早在20世纪50年代就开始对华南退化坡地土壤侵蚀进行长期的观测,并进行植被恢复的研究[4]。经过幾十年的不懈努力,我国在土壤退化及其恢复理论与应用技术方面取得了丰硕的成果,针对不同区域、不同土壤、不同退化类型和程度的生态系统,探索并总结了包括物理、化学和生物的退化土壤修复技术体系。在南方红壤区这一独特的生态系统,也开展了以恢复植被、控制水土流失为目标的荒山绿化、水土流失综合治理研究并取得显著成效[5]。
虽然不同学者对土壤退化定义的论述具有差异性,但是都着重强调土壤质量的降低,土壤退化和土壤质量具有千丝万缕的联系[6]。微生物是土壤生态系统的重要组成部分,几乎所有的土壤过程都直接或间接与其有关,越来越受到科学家的密切关注[4,6]。土壤是微生物的重要栖息地,其理化性质和养分含量对微生物群落具有重要影响,一定程度上决定了微生物的种类、数量和多样性,而土壤微生物的结构和多样性的变化也可以反映土壤质量的变化趋势。微生物活动对健康和肥沃的土壤至关重要,是土壤质量和退化土壤修复评价的一项重要指标[7]。
随着生物学技术的进步和退化土壤修复研究的深入,人们对退化土壤微生物群落组成与多样性以及修复措施对土壤微生物的影响等方面开展了广泛的研究,但是对退化土壤中的微生物群落功能组成的系统研究还相对较少,多集中在微生物硝化作用、反硝化作用以及固氮能力等单一功能的研究。土壤微生物的分类学多样性与功能多样性有密切联系,但并没有显著相关性[8-9],所以土壤微生物群落结构与功能组成的系统研究有助于土壤微生态过程的全面揭示。因此,本研究通过室内盆栽模拟野外退化土壤的修复,通过对细菌16S rRNA进行高通量测序并借助细菌PICRUSt基因功能预测技术,研究稻壳生物炭、微生物有机肥以及化学肥料施用等措施对南方退化红壤土壤微生物功能组成的影响。
1 材料与方法
1.1 试验材料
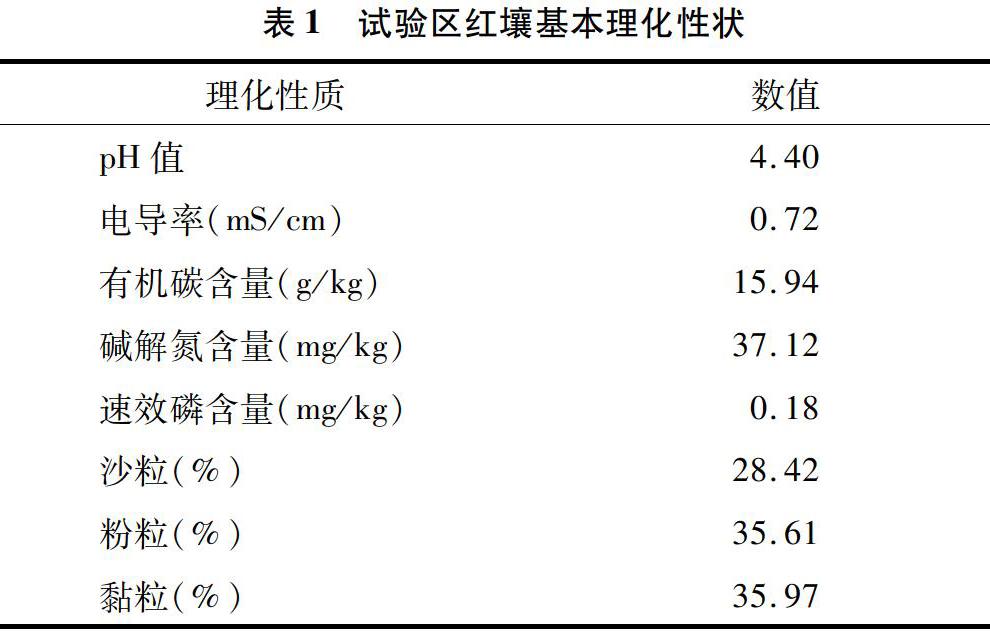
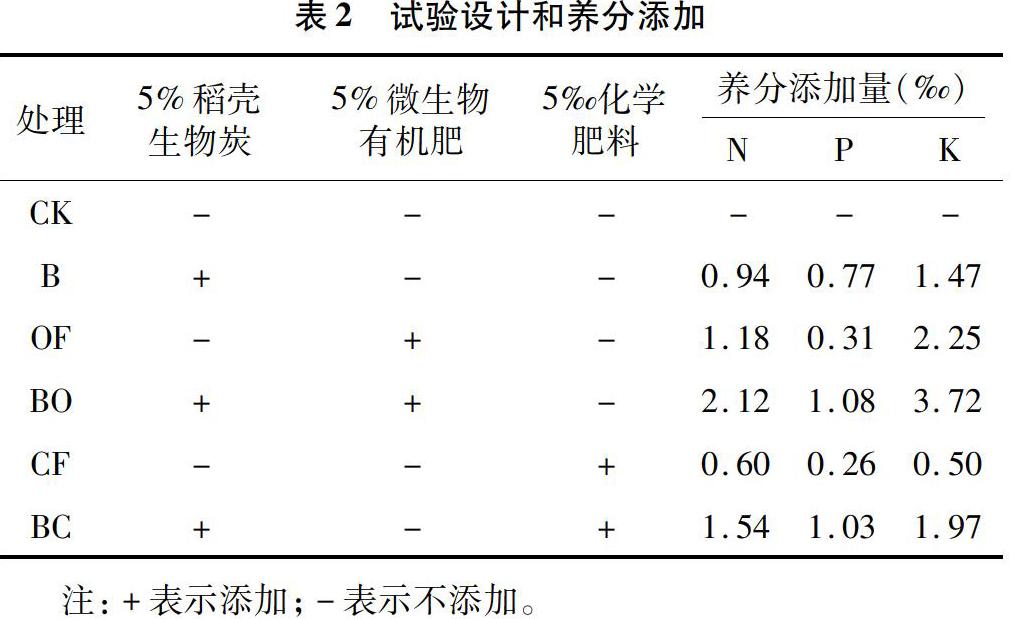
南昌市黄马乡试验基地位于116°1′3.57″E、28°22′28.28″N,土壤成土母质是第四纪红黏土,由于长期过度利用,造成水土流失、土壤板结、养分短缺等问题,土壤退化严重。挖取0~20 cm的表层土壤,过2 mm筛后风干备用,其基本理化性状见表1。稻壳生物炭以干燥稻壳为原料,通过炭化炉在 500 ℃、无氧状态下制备而成,pH值为8.36,养分包括48.59% C、1.87% N、1.53% P、2.94% K,含灰分11.43%。微生物有机肥是将畜禽粪便发酵完全后与多黏芽孢杆菌复合而成,化学肥料为史丹利农业集团股份有限公司生产的硫酸铵型复合肥,均购买于当地农资市场。微生物有机肥含44.20%的有机质和2.35% N、0.62% P、4.49% K,多黏芽孢杆菌活菌数为3×108 CFU/g,化学复合肥中N、P、K含量分别为12.0%、5.24%、9.96%。本试验采用室内模拟盆栽方式进行,分别添加稻壳生物炭(B)、微生物有机肥(OF)以及化学肥料(CF)、生物炭配施微生物有机肥(BO)、生物炭配施化学肥料(BC)等方式对退化红壤进行修复,以空白作为对照(CK),共设置6个试验处理,详细设计方案和养分添加见表2,每个处理重复3次。2019年8月25日以塑料花盆(口径15 cm×高12 cm,底部有孔)作为栽培容器,装填1.0 kg混合均匀的基质后移栽2株1月龄健壮车前幼苗。试验在江西省科学院智能人工气候室中进行。试验期间保持室内温度为20~25 ℃,空气湿度为60%。植物生长期间定期浇灌去离子水,称质量法保持土壤含水量在20%(最大田间持水量60%)。植物生长90 d后,移除植物地上部和根系后,收集土壤样品,过2 mm土壤筛,混匀后分成2份,一份自然风干用于土壤理化性状测定,一份真空冷冻干燥后置于-80 ℃超低温保藏,用于土壤细菌分子生态学分析。
1.2 土壤理化性状测定
土壤pH值和电导率采用PHS-3C型酸度计测定(测定pH值的水土比为2.5 ∶ 1,测定电导率的水土比为 5 ∶ 1,FE20-Five Easy PlusTM,Switzerland);土壤有机碳含量采用重铬酸钾氧化还原滴定法[10]测定;土壤可溶解有机碳含量采用沸水浸提法[11]测定;采用硼酸吸收盐酸滴定法测定土壤碱解氮含量[12];土壤有效磷含量采用氟化铵-盐酸溶液浸提,钼蓝比色法测定[13];土壤速效钾含量采用中性醋酸铵溶液浸提,火焰光度计法测定。
1.3 高通量测序
每个土壤样品准确称取冷冻干燥后的土壤 500 mg。使用试剂盒Fast DNA SPIN Kit for Soil(Q BIOgene Inc.,Carlsbad,CA,USA)提取土壤总DNA,按照试剂盒内的操作流程进行。提取的DNA经0.8%琼脂糖凝胶电泳和Nanodrop ND-1000 UV-Vis Spectrophotometer(NanoDrop Technologies)检测后保存于-20 ℃冰箱中备用。
合成带有barcode标签序列(用于区分不同的样品)及接头序列的特异引物515F/806R(前引物515F:5′-GTGCCAGCMGCCGCGGTAA-3′;后引物806R:5′-GGACTACVSGGGTATCTAAT-3′),采用PCR扩增技术对土壤细菌16S rRNA基因高变区的V4-V5进行扩增。PCR扩增采用25 μL反应体系,包括1 μL的模板DNA(10 ng/L)、1 μL上下游引物[10 pmol/μL,生工生物工程(上海)股份有限公司]、12.5 μL 2×PCR缓冲液(TaKaRa公司,大连)、0.5 μL DNA Taq 聚合酶(2.5 U/μL,TaKaRa公司),最后加灭菌超纯水至25 μL。反应条件为:95 ℃预变性3 min;95 ℃变性30 s,58 ℃退火30 s,72 ℃延伸 1 min,35个循环;72 ℃再延伸10 min。每个样品3个重复,将同一样品的PCR产物混合后使用QIAEXDNA(QIAGEN Inc,USA)胶纯化试剂盒回收纯化PCR产物,并用Nanodrop ND-1000 UV-Vis Spectrophotometer(NanoDrop Technologies)对回收产物进行检测。按照每个样品的测序量要求,进行相应比例混合并构建MiSeq文库,在Illumina MiSeq (PE×300)平台上进行高通量测序。
1.4 數据分析
对MiSeq测序得到的数据进行拼接,同时对序列的质量和拼接效果进行质控过滤,并根据序列末端的box序列校正序列方向。拼接后的数据用QIIME (version 1.8.0)软件[14]进一步质控:序列长度小于200 bp,连续5个碱基平均测序质量小于30,含有模糊碱基,引物碱基中有超过1个的错配,以及无法被任何barcode 识别的序列,所有包含这些条件的序列都会被丢弃。采用UCHIME[15]借助16S核糖体基因序列的Silva数据库(version 119)[16]检测并去除嵌合体序列;采用Usearch对所有序列参照Greengenes数据库以97%的相似度进行OTU聚类分析,得到OTU列表并对其进行序列数量标准化,借助PICRUSt软件对细菌16S rRNA基因功能进行预测。每个土壤样品的细菌群落丰富度是通过计算每个样品的OTU获得的。细菌物种数和土壤理化性状之间的相关关系采用线性回归方法进行分析。拟合适合度采用调整的R2和P值进行判定。基于Bray-Curtis 相似性矩阵的PCoA 分析用于分析土壤细菌群落组成和预测基因功能多样性的变异,并用多元的相似性分析方法ADONIS(Adonis test,Vegan package in R)检验变异的显著程度[17]。应用统计分析软件SPSS 18.0计算微生物多样性指数,预测功能基因丰度平均值和标准差,并进行独立样本t检验,确定各处理样品间微生物多样性和预测功能基因差异显著性。
2 结果与分析
2.1 土壤细菌多样性
本研究包括18个土壤样品,测序下机数据通过拼接、质控后共计得到1 201 631条有效序列,根据97%的相似度划分为3 669个OTU。平均每个样品获得66 757条序列,每个样品的序列数在43 548~74 917条之间。经过比对、注释和分类统计得到32个细菌门分类单元,其中优势分类门(>细菌群落总丰度的1%)分别为变形菌门(Proteobacteria)、放线菌门(Actinobacteria)、绿弯菌门(Chloroflexi)、Patescibacteria、WPS-2菌门、酸杆菌门(Acidobacteria)、芽单胞菌门(Gemmatimonadetes)、厚壁菌门(Firmicutes)、浮霉菌门(Planctomycetes)、拟杆菌门(Bacteroidetes),占总序列数的95%以上。优势分类门在不同土壤修复措施下具有显著差异,其中变化最大的为厚壁菌门。
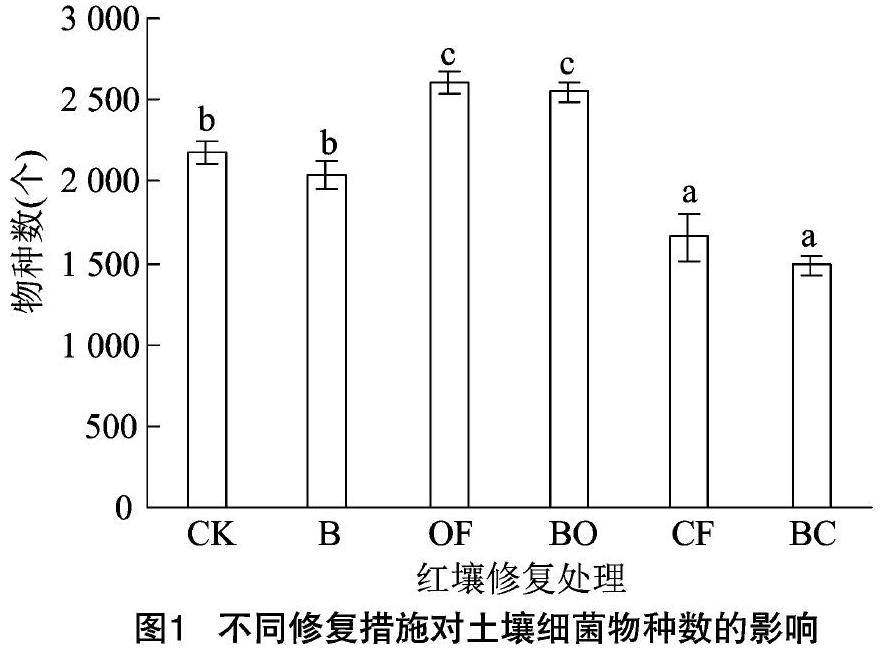
对不同修复措施下土壤细菌物种数的统计分析发现,生物有机肥的施用显著提高了土壤细菌物种数,化学复合肥料的施用显著降低了土壤细菌物种数,而稻壳生物炭的施用没有产生显著的影响(图1)。进一步分析发现,土壤细菌物种数(OTU)与土壤pH值呈正相关,而与土壤碱解氮和有效磷含量呈负相关(图2),都达到了极显著的水平(P<0.01)。
2.2 土壤细菌群落结构变化
基于Bray-Curtis距离的细菌群落组成,在PCoA分析的第一、第二排序轴上明显区分成3个大的区域(图3),分别为不施肥组、施用微生物有机肥组、施用化学复合肥组(Adonis F2,15=25.55,R2=0.773 1,P<0.001),解释率达到68.7%。各大组均包括施用生物炭和不施用生物炭,但是无法根据生物炭施用与否进行进一步的划分(P>0.05)。
2.3 土壤细菌功能组成
基于KEGG数据对比样品中的细菌一级功能分类,各个修复处理间细菌一级功能基因丰度差异见图4。已知的六大类生物代谢通路中,代谢(Metabolism)相关基因丰度最高,其次是遗传信息处理和环境信息处理,而细胞过程、有机系统和人类疾病相关基因丰度很低。统计分析结果表明,化学肥料(CF和BC)施用显著降低代谢和环境信息处理相关基因丰度(P<0.05),有机肥(OF和BO)的施用对其丰度提升不显著。有机肥和化学肥料的施用对遗传信息处理、人类疾病和细胞过程相关基因丰度的影响不显著(P>0.05)。另外,化学肥料的施用还显著降低了有机系统相关基因丰度。稻壳生物炭单独或与肥料混合施用都对细菌一级功能基因丰度无显著影响。
上述生物代谢通路可以进一步细分为二级功能层,共包含40种子功能,根据Bray-Curtis距离计算出预测功能基因β多样性,通过PCoA分析发现,所研究的18个土壤样品的区分不明显(图5),大致可以区分为对照组、化学肥料组以及其他(Adonis F2,15=17.22,R2=0.6966,P<0.001)。
2.4 土壤细菌氮循环
进一步研究不同类型肥料施用下车前根际土壤细菌参与氮循环途径(三级功能层,ko00910)的基因差异,发现有机肥施用显著提高氮循环途径基因丰度,化学肥料施用降低其丰度,而稻壳生物炭单独施用或与肥料混合施用都没有表现出明显的影响(图6)。
结果表明,本试验中稻壳生物炭对土壤细菌群落结构与功能组成无显著性影响,所以接下来对氮循环5个阶段(氮异化还原、氮同化还原、反硝化作用、固氮作用和硝化作用)相关功能基因丰度分析只在施用有机肥(OF)、化学肥料(CF)的修复处理和空白对照(CK)之间进行。总体而言,退化红壤车前根际细菌的氮异化还原、氮同化还原以及反硝化作用相关基因丰度最高,固氮作用次之,硝化作用最低(图7)。在车前根际土壤细菌氮异化还原过程中,亚硝酸还原酶基因nirB、硝酸还原酶基因narL和异化还原酶基因nrfA对各修复处理的响应基本一致,有机肥显著提升其丰度,而化学肥料处理其丰度显著降低或降低不显著。化学肥料施用显著降低亚硝酸还原酶基因nirD丰度,而有机肥无显著影响。氮同化还原过程中,硝酸还原酶基因nasA和nirA丰度高于nasB和narB,nasA和nirA丰度均表现为CK>OF>CF。反硝化作用过程中,化学肥料施用显著增加亚硝酸还原酶基因nirK和一氧化氮还原酶基因norB丰度,降低一氧化氮还原酶基因narC、亚硝酸还原酶基因narH以及硝酸还原酶基因napA和napB丰度,而有机肥施用显著增加亚硝酸还原酶基因narH和氧化亚氮还原酶基因nosZ丰度。固氮作用过程的固氮酶基因nifD、nifH、anfG和nifK丰度大小均表现为OF>CK>CF处理。硝化作用过程相关基因丰度明显低于其他氮循环过程,其中羟胺脱氢酶基因hao丰度表现为CK>OF>CF处理,其他基因丰度无显著差异。
3 讨论与结论
植物根际土壤微生物是根部最活跃的部分,能够敏感地反映出土壤生态系统的变化[4,6,18]。本研究发现,有机肥施用显著增加退化红壤车前根际细菌物种数,提升代谢和环境信息处理通路基因丰度,特别是土壤氮循环途径基因丰度,调控土壤氮循环过程。化学肥料的施用基本上发挥了相反的作用,而稻壳生物炭的影响不明显。根际土壤细菌物种数以及生物代谢途径基因丰度,反映了土壤同化和矿化的能力及方向,在土壤养分循环和能量转化过程中起着重要作用,是土壤生态系统肥力的重要生物学指标[19],表明有机肥施用能对退化红壤起到很好的修复作用,提升土壤质量和土地生产力。
相关研究表明,生物炭和肥料的单独施用都对土壤微生物群落组成表现出较强的调节作用[20-21],有效提升土壤质量。本研究将生物炭与微生物有机肥以及化学肥料相结合,并应用于退化红壤的生态修复,结果表明,微生物有机肥能显著增加细菌群落物种多样性,而化学肥料表现出相反的作用,与前人研究结果[22]一致。施用微生物有机肥向土壤补充活泼的碳源,促进异养微生物群落发展壮大,增强其代谢活性[23],这可能是微生物有机肥提升土壤微生物群落的重要原因。然而,稻壳生物炭对细菌群落多样性没有显著的影响。细菌群落主成分(PCoA)分析结果也显示,所有样品可以根据肥料类型划分为3类,包括微生物有机肥相关修复、化学肥料相关修复以及不施肥对照,生物炭施用与否区分不明显。
稻壳生物炭施用对退化红壤中细菌群落结构和多样性没有表现出显著的影响,这也体现在土壤微生物群落功能组成上,包括基于KEGG的生物代谢通路功能基因丰度、氮循环途径功能基因丰度以及氮循环过程相关功能基因丰度。已有研究发现,多种类型生物炭的施用对土壤微生物群落具有很好的调节作用[20,24-25],这可能是由生物炭自身特性及剂量、土壤类型、环境条件与微生物表征方法等差异引起的,本试驗中生物炭施用对土壤微生物群落没有发挥显著作用的原因还有待进一步深入研究。
基于16S rRNA基因的高通量测序,比对KEGG数据库对细菌群落功能组成进行预测分析,结果表明,施用不同类型肥料对微生物群落功能组成具有显著的影响,施用微生物有机肥能显著增加生物代谢通路功能基因丰度,提升代谢途径中的氮循环途径功能基因丰度(ko00910),而化学肥料的施用基本表现出相反的作用。肥料施用对细菌群落功能组成的影响与其对细菌群落结构组成的影响基本一致,说明基于肥料施用的退化红壤的修复效应是深远的,微生物有机肥的施用能够明显改善退化土壤质量,也将显著提升土地的生产力。
对氮循环过程相关的功能基因分析结果表明,微生物有机肥增加氮异化还原、氮固定以及部分反硝化作用相关的功能基因丰度,究其原因可能是反硝化、固氮等微生物多为异养微生物,有机物料可以为土壤提供更均衡和稳定的营养供应,促进它们生长繁殖[26],这有利于促进土壤中氮素养分的维持[27],而化学肥料表现出显著的抑制作用。大量研究表明,农田土壤中施加氮肥可以显著提高参与反硝化作用过程的nirK和norB基因丰度[28-29],与本研究结果一致。nirK和norB是反硝化作用过程的2个关键基因[27],化学肥料的施用通过增加它们的丰度,促进土壤氮素向空气中扩散,这不利于环境保护和土壤质量。
本研究结果显示,结合细菌16S RNA基因高通量测序和PICRUSt功能基因预测,分析并比较了稻壳生物炭、微生物有机肥和化学肥料在典型南方退化红壤修复中对土壤微生物群落结构和功能组成的影响。微生物有机肥对退化红壤的修复增加车前根际土壤细菌群落多样性,有效调节细菌的微生物群落结构,而化学肥料施用则表现出相反的作用。微生物有机肥施用显著改善了土壤微生物群落功能组成,提高生物代谢途径和氮循环途径功能基因丰度。另外,微生物有机肥的施用还调节了氮循环过程相关基因丰度,有利于土壤氮素的维持,改善土壤质量,而化学肥料施用则相反。稻壳生物炭单独施用或与肥料混合施用对微生物群落结构和多样性无显著影响,表现在土壤微生物群落功能组成上。
参考文献:
[1]李庆逵. 中国红壤[M]. 北京:科学出版社,1983.
[2]江西土地資源管理局. 江西土壤[M]. 北京:中国农业科学技术出版社,1991.
[3]吴 卓,戴尔阜,林媚珍. 气候变化和人类活动对南方红壤丘陵区森林生态系统影响模拟研究——以江西泰和县为例[J]. 地理研究,2018,37(11):2141-2152.
[4]余作岳,彭少麟,丁明懋. 热带亚热带退化生态系统植被恢复生态学研究[M]. 广州:广东科技出版社,1996.
[5]黄小春,宋小妹. 江西低丘红壤水土流失区植被恢复策略及理论探讨[J]. 江西林业科技,2008(6):42-47.
[6]Doran J W. Defining soil quality for sustainable environment[M]. Wisconsin:Soil Science Society of America,1994.
[7]Cruz-Paredes C,Wallander H,Kjller R,et al. Using community trait-distributions to assign microbial responses to pH changes and Cd in forest soils treated with wood ash[J]. Soil Biology and Biochemistry,2017,112:153-164.
[8]Harsch M A,Hulme P E,McGlone M S,et al. Are treelines advancing? A global meta-analysis of treeline response to climate warming[J]. Ecology Letters,2009,12(10):1040-1049.
[9]Shen C C,Shi Y,Ni Y Y,et al. Dramatic increases of soil microbial functional gene diversity at the treeline ecotone of Changbai Mountain[J]. Frontiers in Microbiology,2016,7:1184.
[10]Yuan C L,Mou C X,Wu W L,et al. Effect of different fertilization treatments on indole-3-acetic acid producing bacteria in soil[J]. Journal of Soils and Sediments,2011,11(2):322-329.
[11]Chantigny M H. Dissolved and water-extractable organic matter in soils:A review on the influence of land use and management practices[J]. Geoderma,2003,113:357-380.
[12]Cornfield A H. Ammonia released on treating soils with N sodium hydroxide as a possible means of predicting the nitrogen-supplying power of soils[J]. Nature,1960,187:260-261.
[13]Gyaneshwar P,Kumar G N,Parekh L J,et al. Role of soil microorganisms in improving P nutrition of plants[M]//Food security in nutrient-stressed environments:exploiting plants genetic capabilities. Netherlands:Springer,2002.
[14]Caporaso J G,Kuczynski J,Stombaugh J,et al. QIIME allows analysis of high-throughput community sequencing data[J]. Nature Methods,2010,7(5):335-336.
[15]Edgar R C,Haas B J,Clemente J C,et al. UCHIME improves sensitivity and speed of chimera detection[J]. Bioinformatics,2011,27(16):2194-2200.
[16]Pruesse E,Quast C,Knittel K,et al. SILVA:a comprehensive online resource for quality checked and aligned ribosomal RNA sequence data compatible with ARB[J]. Nucleic Acids Research,2007,35(21):7188-7196.
[17]R Development Core Team. R:a language and environment for statistical computing[M]. Vienna:the R Foundation for Statistical Computing,2012.
[18]李兰君,刘玳含,刘建斌,等. 连作对设施番茄土壤微生物及酶活性的影响[J]. 江苏农业科学,2018,46(18):130-134.
[19]王 涛,乔卫花,李玉奇,等. 轮作和微生物菌肥对黄瓜连作土壤理化性状及生物活性的影响[J]. 土壤通報,2011,42(3):578-583.
[20]Kolton M,Graber E,Tsehansky L,et al. Biochar-stimulated plant performance is strongly linked to microbial diversity and metabolic potential in the rhizosphere[J]. The New Phytologist,2017,213(3):1393-1404.
[21]Qiu M H,Zhang R F,Xue C,et al. Application of bio-organic fertilizer can control Fusarium wilt of cucumber plants by regulating microbial community of rhizosphere soil[J]. Biology and Fertility of Soils,2012,48(7):807-816.
[22]Zhen Z,Liu H T,Wang N,et al. Effects of manure compost application on soil microbial community diversity and soil microenvironments in a temperate cropland in China[J]. PLoS One,2014,9(10):e108555.
[23]Fierer N,Bradford M A,Jackson R B. Toward an ecological classification of soil bacteria[J]. Ecology,2007,88(6):1354-1364.
[24]Zhang Y C,Wang X,Liu B J,et al. Comparative study of individual and co-application of biochar and wood vinegar on blueberry fruit yield and nutritional quality[J]. Chemosphere,2020,246:125699.
[25]方 明,李 洁,赖 欣,等. 短期生物炭刺激对红壤和潮土微生物群落的影响[J]. 江苏农业科学,2020,48(11):250-258.
[26]任灵玲. 长期施肥棕壤中氮代谢功能基因的变化特征[D]. 沈阳:沈阳农业大学,2019.
[27]Welsh A,Chee-Sanford J C,Connor L M,et al. Refined NrfA phylogeny improves PCR-based nrfA gene detection[J]. Applied and Environmental Microbiology,2014,80(7):2110-2119.
[28]陈 晨,许 欣,毕智超,等. 生物炭和有机肥对菜地土壤N2O排放及硝化、反硝化微生物功能基因丰度的影响[J]. 环境科学学报,2017,37(5):1912-1920.
[29]Yin C,Fan F L,Song A L,et al. Different denitrification potential of aquic brown soil in Northeast China under inorganic and organic fertilization accompanied by distinct changes of nirS-and nirK-denitrifying bacterial community[J]. European Journal of Soil Biology,2014,65:47-56.
