茶树WOX基因家族的鉴定和表达模式分析
2021-09-10艾安涛张宝会彭徐乾吕立堂
艾安涛 张宝会 彭徐乾 吕立堂

摘要:WOX转录因子在植物的生长发育和非生物胁迫响应中起着重要的调控作用。文章基于全基因组数据,从茶树基因组中鉴定出29个WOX基因,并对其基因结构、进化关系、保守域、染色体定位进行分析,同时分析了它们在PEG诱导的干旱胁迫、盐胁迫处理中的转录组数据。结果表明,29个CsWOX(茶树WOX)基因在茶树染色体上分布不均;根据进化关系将茶树WOX基因分为4类;基因结构和保守基序分析发现相同亚家族的基因结构和保守结构域基本一致;基因表达分析显示,CsWOX基因在花和果实组织中具有较高的表达水平,且部分基因随着叶片成熟度的增加,表达水平升高;不同的CsWOX基因在PEG诱导的干旱胁迫、盐胁迫处理下存在差异表达,说明CsWOX基因广泛参与茶树生长发育并在响应非生物胁迫中发挥重要作用。该结果将为进一步研究WOX基因在调控茶树生长发育和非生物胁迫响应中的作用提供一些有价值的信息,为茶树WOX基因的功能研究与利用提供科学依据。
关键词:茶树;WOX基因家族;转录因子;基因表达分析
Identification and Expression Pattern Analysis of
WOX Gene Family in Camellia sinensis (L.)
AI Antao1, ZHANG Baohui2, PENG Xuqian1, L Litang1,2*
1. College of Tea Science, Guizhou University, Guiyang 550025, China; 2. Institute of Agro-Bioengineering, Guizhou
University College of Life Sciences, Key Laboratory of Plant Resources Conservation and Germplasm
Innovation in Mountainous Regions, Ministry of Education, Guiyang 550025, China
ü
Abstract: WOX transcription factors play an important regulatory role in plant growth, development and response toabiotic stresses. Based on the tea whole genome data, this study identified 29 WOX genes and analyzed their genestructures, evolutionary relationship, conserved domains, and chromosome locations. Meanwhile, their responses to drought stress and salt stress induced by PEG were studied. The results show that 29 CsWOX genes were unevenlydistributed on tea plant chromosomes. They were classified into category IV according to evolutionary relationship. Their gene structures and conserved domains of the same subfamily were basically the same. Gene expression analysisshows that the CsWOX genes had higher expression levels in flower and fruit tissues, and the expressions of somegenes increased with the increase of leaf maturity. CsWOX genes exhibited different expression patterns underPEG-induced drought stress and salt stress, indicating that CsWOX genes are widely involved in tea plant growth anddevelopment and play different roles in response to abiotic stress. This result will provide some valuable informationfor further research on the role of WOX gene in regulating the growth and development of tea plants and response toabiotic stresses. It also provided a scientific basis for the functional research and utilization of tea plant WOX genes.
Keywords: Camellia sinensis (L.), WOX gene family, genome identification, gene expression analysis
WOX蛋白是植物特有的一類转录因子,其名称源于一段由约60个氨基酸组成的高度保守的螺旋-环-螺旋-拐角-螺旋结构域,此外,在WUS组或者I组的WOX蛋白氨基酸C端还存在一个高度保守的基序,命名为WUS-box (TLXLTP),该基序在WOX蛋白调控植物生长发育进程中起重要的作用[1]。
近几年,研究者通过正向遗传和反向遗传方法对不同植物的研究表明,WOX转录因子对植物的生长发育、干细胞稳态维持、抗逆性、初生和次生物质代谢、植物激素信号转导等有重要的调控作用,如柳枝稷WOX转录因子STF基因通过直接抑制下游细胞分裂素氧化酶相关基因的表达,增加了转基因水稻叶片生物量和糖的含量[2];WOX11通过抑制RR2基因的表达,调控水稻冠根的生长发育,在水稻中提高WOX11的表达增加了冠根的数目和生长速度,相反,降低该基因的表达导致冠根数目和生长速度降低[3],并且WOX11基因通过调控水稻根的发育,增加了转基因水稻对干旱胁迫的抗性[4];拟南芥WOX5功能缺失突变体促进干细胞分化,反之,WOX5超表达降低了干细胞的分化,该结果表明,WOX5在维持拟南芥干细胞稳态中起关键的调控作用[5];拟南芥WOX13通过抑制JAGGED基因(该基因编码一个包含锌指结构域的蛋白,参与拟南芥外侧器官形态发生的调控)的表达,促进拟南芥果实胎座框(Replum)的形成[6]。以上研究结果表明,WOX转录因子主要通过改变下游基因的表达,影响植物生长发育进程。
基于WOX蛋白全长氨基酸序列的相似性和拓扑结构,拟南芥WOX基因家族蛋白被分成3个组,即组Ⅰ、Ⅱ和Ⅲ[1]。目前,随着全基因组学和转录组学测序技术的发展,许多物种WOX转录因子已经被鉴定和分析,如拟南芥、水稻、桑树、陆地棉等[7-8]。值得注意的是,关于WOX蛋白的克隆和功能研究主要集中在拟南芥、水稻等模式植物中,在茶树中未有报道。
茶树是我国重要的经济作物,良好的生态环境是茶树正常生长、茶叶品质形成的主要影响因素,然而,茶树生长过程中不可避免受到干旱、低温等多种逆境胁迫,极大地影响了茶园产量。目前,对茶树WOX家族基因的研究报道较少,关于茶树WOX家族基因逆境相关的研究更是很少有报道。
本试验采用生物信息学等方法,对茶树WOX家族基因进行了基因组鉴定,并用鉴定到的茶树、拟南芥以及水稻WOX基因构建了进化树,并进一步对其基因结构、保守结构域、染色体分布、启动子顺式作用元件以及逆境下的表达模式进行分析,能够获得对茶树WOX家族基因特性更多的了解,为茶树WOX家族基因机制和抗逆响应中的功能奠定基础。
一、材料与方法
1. 茶树WOX基因家族的筛选与分析
从茶树的全基因组数据库(http://tpia.teaplant.org)和拟南芥的全基因组数据库(https://www.arabidopsis.org)中下载两者的全基因组数据、启动子序列、染色体定位、DNA、CDS信息等。首先从 Pfam (http://pfam.xfam.org)中下载WOX基因特有的保守结构域模型(PF00046),利用该模型在茶树蛋白数据库中进行BLASTP比对,设定阈值为E<1e-5 ,获得假设的茶树WOX基因,接着利用第一次鉴定到的假设茶树WOX基因序列重新构建关于茶树的WOX基因隐马尔可夫模型,进行第二次搜索鉴定,又一次获得假设的茶树WOX基因。将第二次获得的茶树假设WOX基因蛋白序列提交至NCBI-CDD(https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi,http://smart.embl-heidelberg.de)以及Pfam(http://pfam.xfam.org)进行保守结构域的鉴定,剔除冗余序列,最终得到茶树WOX基因。利用HMM 3.0软件中的perl脚本对茶树WOX蛋白质的分子质量、等电点、氨基酸长度和染色体位置进行分析。
2. 系统进化树、基因结构和蛋白质保守结构域分析
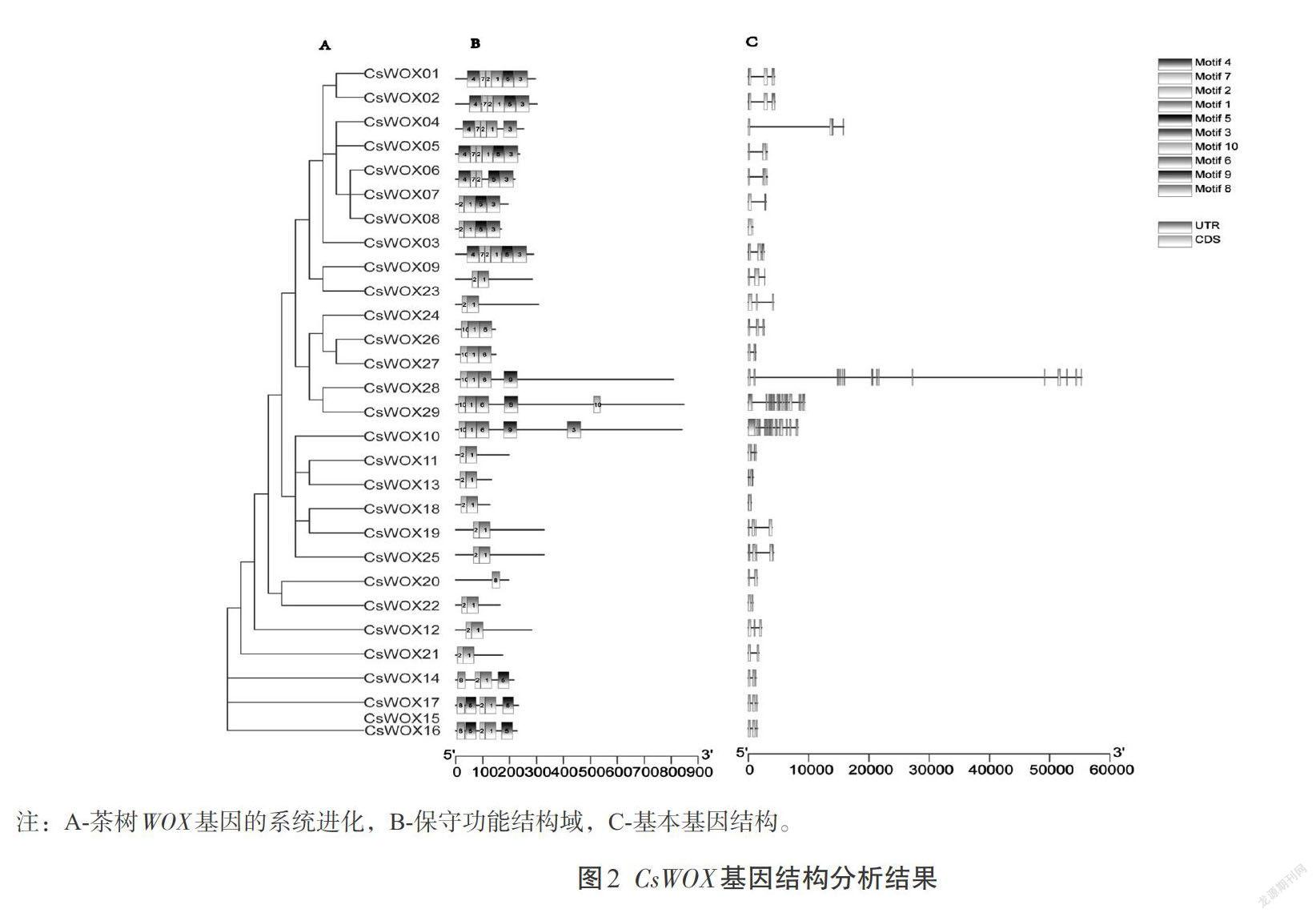
利用DNAMAN软件对茶树、拟南芥和水稻的WOX蛋白序列进行多序列比对,比对结果利用MEGA X软件进行临近法(Neighbor joining, NJ)分析,Boostrap参数设为1 000,构建系统进化树。将CsWOX基因组DNA序列和CDS序列提交至GSDS 2.0(http://gsds.cbi.pku.edu.cn)进行基因结构分析,利用MEME(http://meme-suite.org)进行保守结构域分析,motif数目设置为15个、宽度范围设置为1~50个氨基酸。
3. 染色体分布分析
利用HMM 3.0软件中的perl脚本从茶树的基因组注释信息中获取CsWOX基因在染色体上的起始位置信息。根据CsWOX基因两两之间的BLASTP比对结果获取基因同源复制事件,最后将染色体定位和比对结果利用Circos v0.67工具进行可视化,同源或复制基因用曲线连接,最后利用GSDS 2.0進行可视化。
4. 表达模式分析
为分析茶树CsWOX基因家族的表达情况,从茶树基因组TPIA数据库(http://tpia.teaplant.org)下载它们在茶树不同组织(花、茎、根、顶芽、嫩叶、成熟叶、老叶)、盐胁迫、PEG 诱导的干旱胁迫后的转录组数据(TPM值),同时鉴定差异表达基因[9],结果使用TBtools软件制作热图。
二、结果与分析
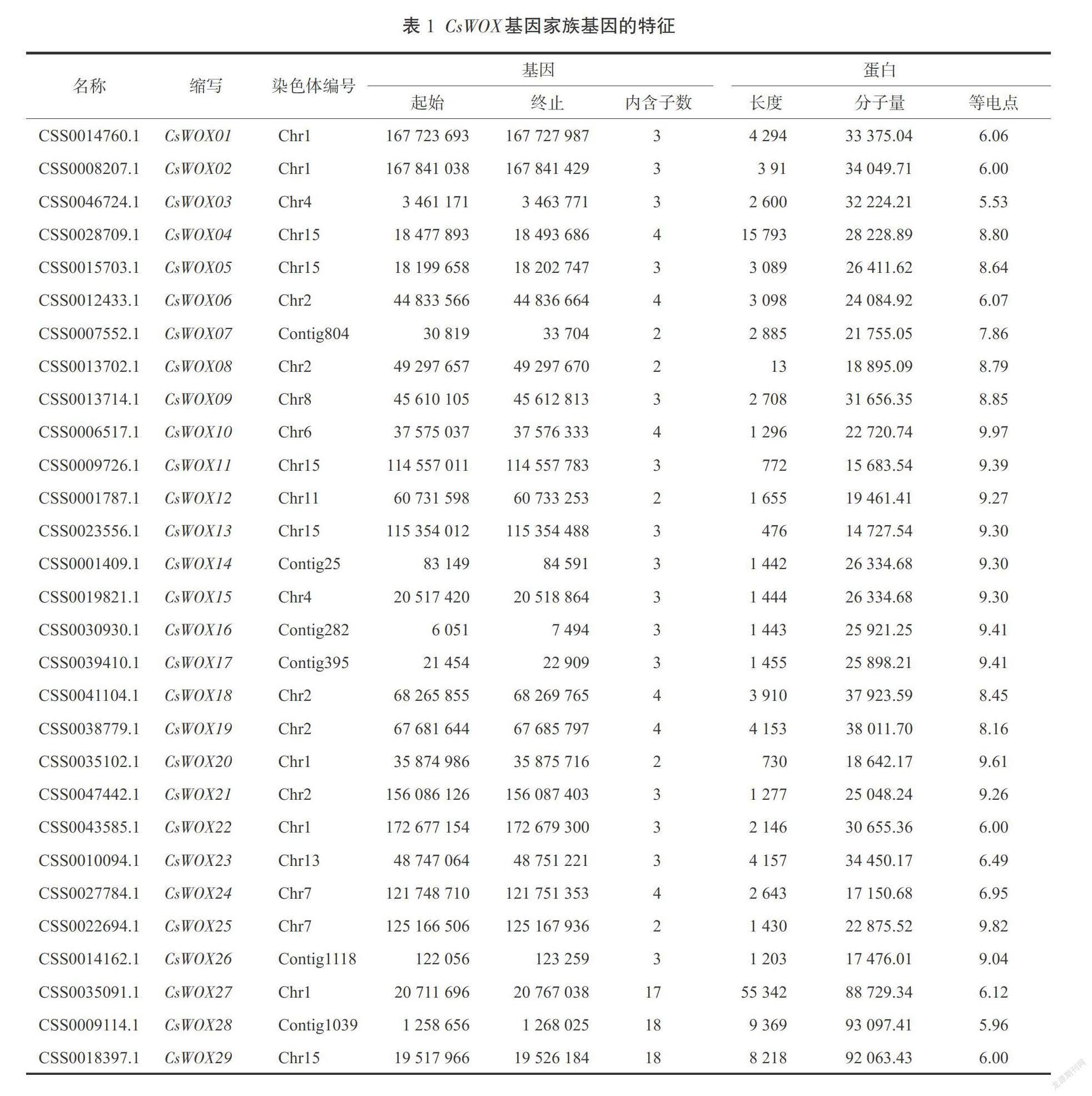
1. 茶树WOX基因家族成员鉴定及理化性质预测
通过2次比对搜索以及将比对的候选基因提交到SMART、Pfam、NCBI-CDD等在线网站进行保守域的结构验证,最终得到了29个茶树WOX基因,并根据基因在染色体的位置及其同源关系对其命名。进一步对茶树WOX基因编码的蛋白质进行理化性质分析(表1),结果显示,茶树最长的WOX蛋白CsWOX27含有55 342个氨基酸残基,最短的CsWOX13含有476个氨基酸残基;相对分子量在14.7(CsWOX13)~93.1 kD(CsWOX28);理论等电点(Pi)介于5.53(CsWOX03)~9.97(CsWOX10)之间,同时发现Pi >7的CsWOX 蛋白达到了65.52%(19个),表明大部分CsWOX蛋白富含碱性氨基酸。
2. 茶树WOX系统进化、基因结构和保守结构域分析
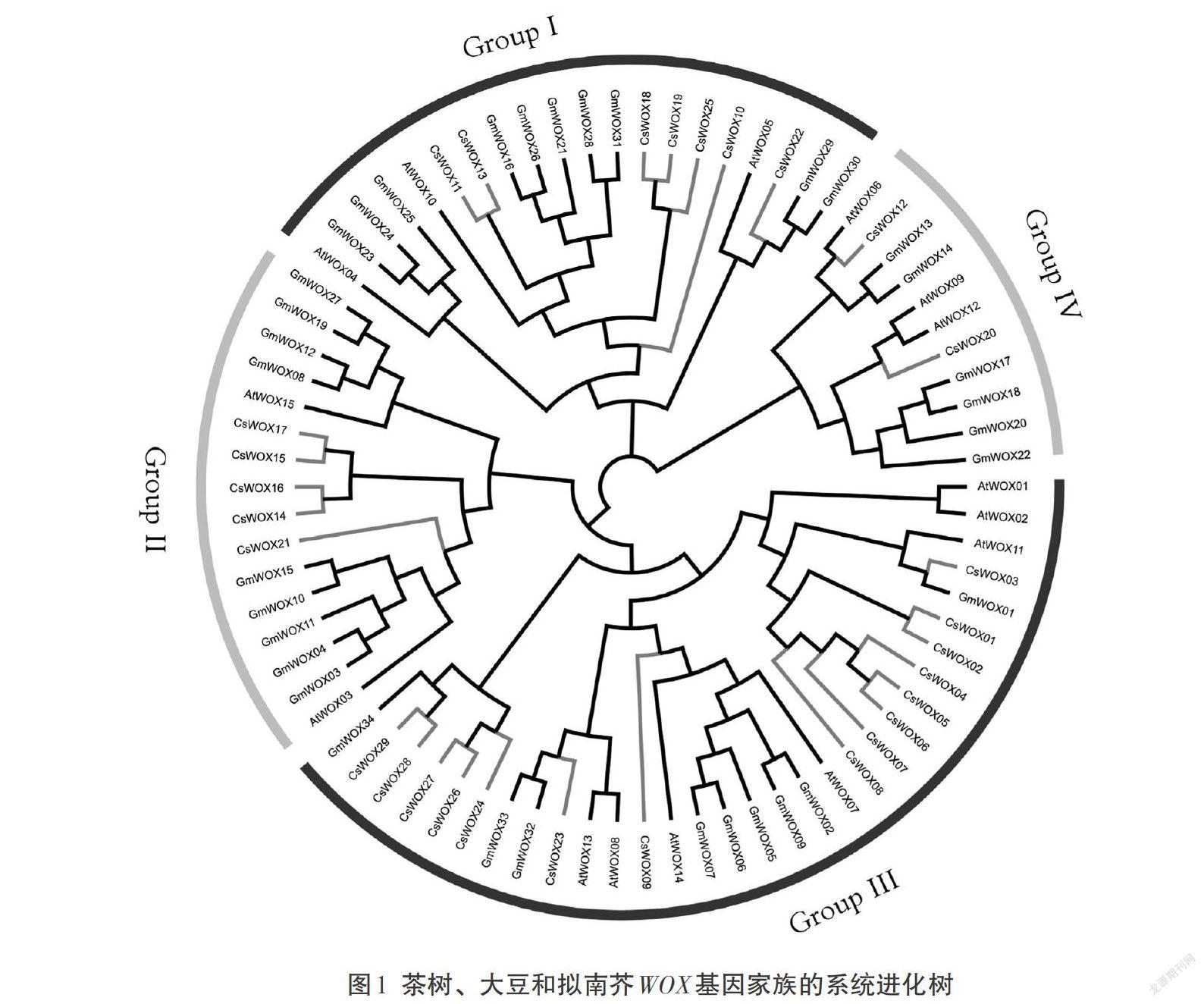
为了明确茶树WOX家族成员的分类,将15个拟南芥WOX基因和34个大豆WOX基因的蛋白序列和29个茶树WOX蛋白进行多序列比对并构建进化树。结果发现茶树CsWOX基因和拟南芥、水稻同源性很高,表明WOX基因物种分化非常保守,蛋白序列的相似性表明这3个物种的WOX基因可能具有相似的生物学功能。根据拟南芥和水稻的同源基因分类,将茶树WOX家族分为4个亚家族(图1)。根据系统进化树分析,WOX基因不仅在茶树、拟南芥及水稻中都有发现,且同源性较高,表明了WOX基因在物种分化过程中具有较高的保守性,进化时间也要早于茶树、拟南芥和大豆的分化时间。
为深入了解茶树WOX家族基因的功能,利用茶树WOX基因组DNA序列和CDS序列对其基因结构进行分析。利用MEME 软件对茶树WOX家族基因的保守基序组成和数目进行分析,一共鉴定到10个保守基序,依次命名为基序1到基序10(图2)。各亚组所包含的保守基序基本一致,表明同一亚组由相似的保守结构域构成,可能具有相似的生物学功能。
3. 染色体定位和CsWOX的同源基因分析
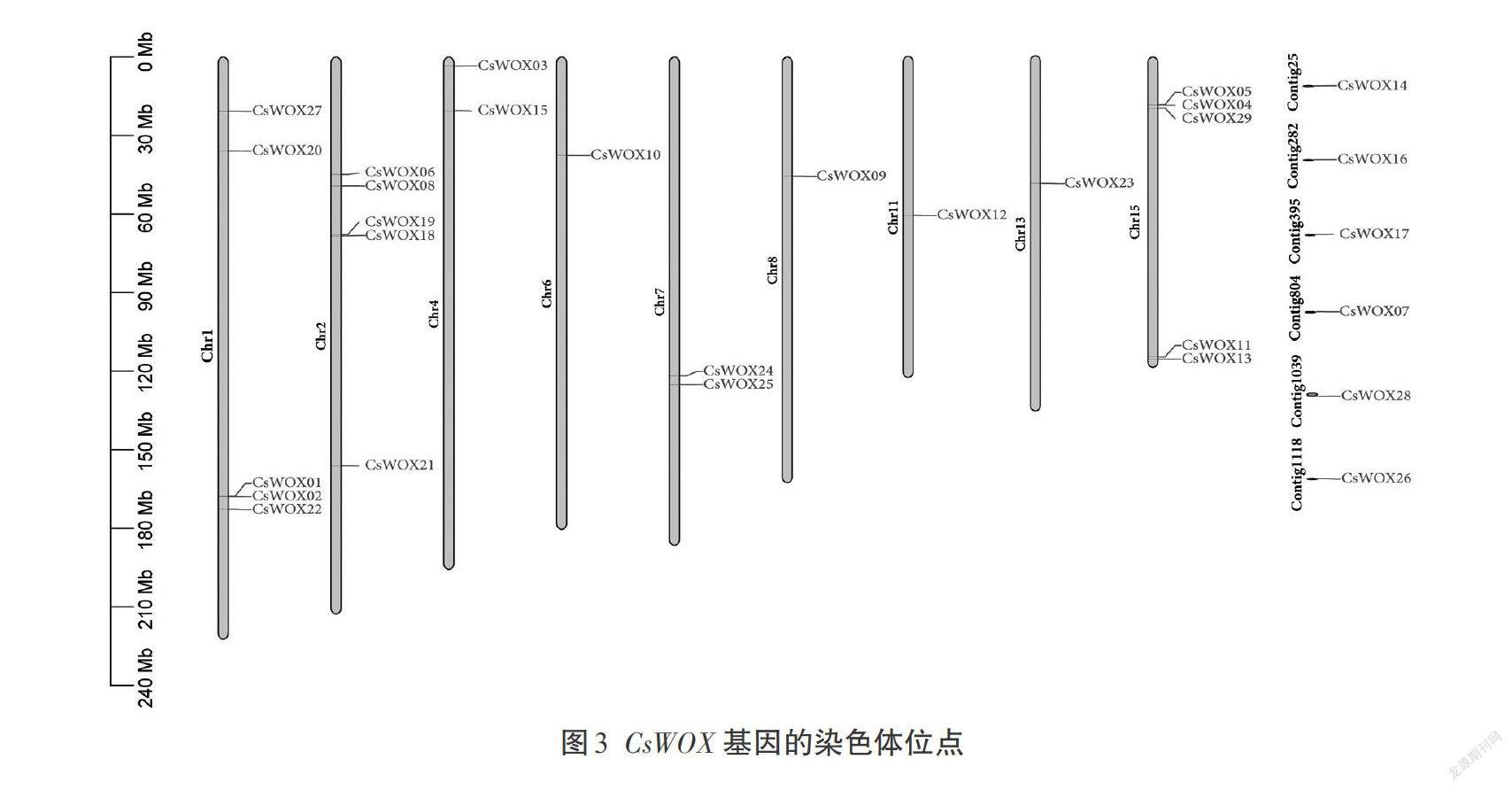
根据茶树基因组染色体注释信息,将鉴定到的CsWOX基因进行染色体定位(图3)。结果显示,6个CsWOX(CsWOX07、CsWOX14、CsWOX16、CsCCWOX17、CsWOX26、CsWOX28)基因不能定位到染色体上,其他23个CsWOX基因不均等地定位到不同染色体上,其中1、2、15号染色体上包含的基因个数最多,均有5个CsWOX基因;而3、5、9、10、12、14号染色体不含CsWOX基因。
此外,茶树不同染色体组中同样分布不均等,其中4、7号染色体有2个,6、8、11、13号染色体有1个,暗示了CsWOX基因在茶树进化过程中可能发生了基因的复制或丢失。
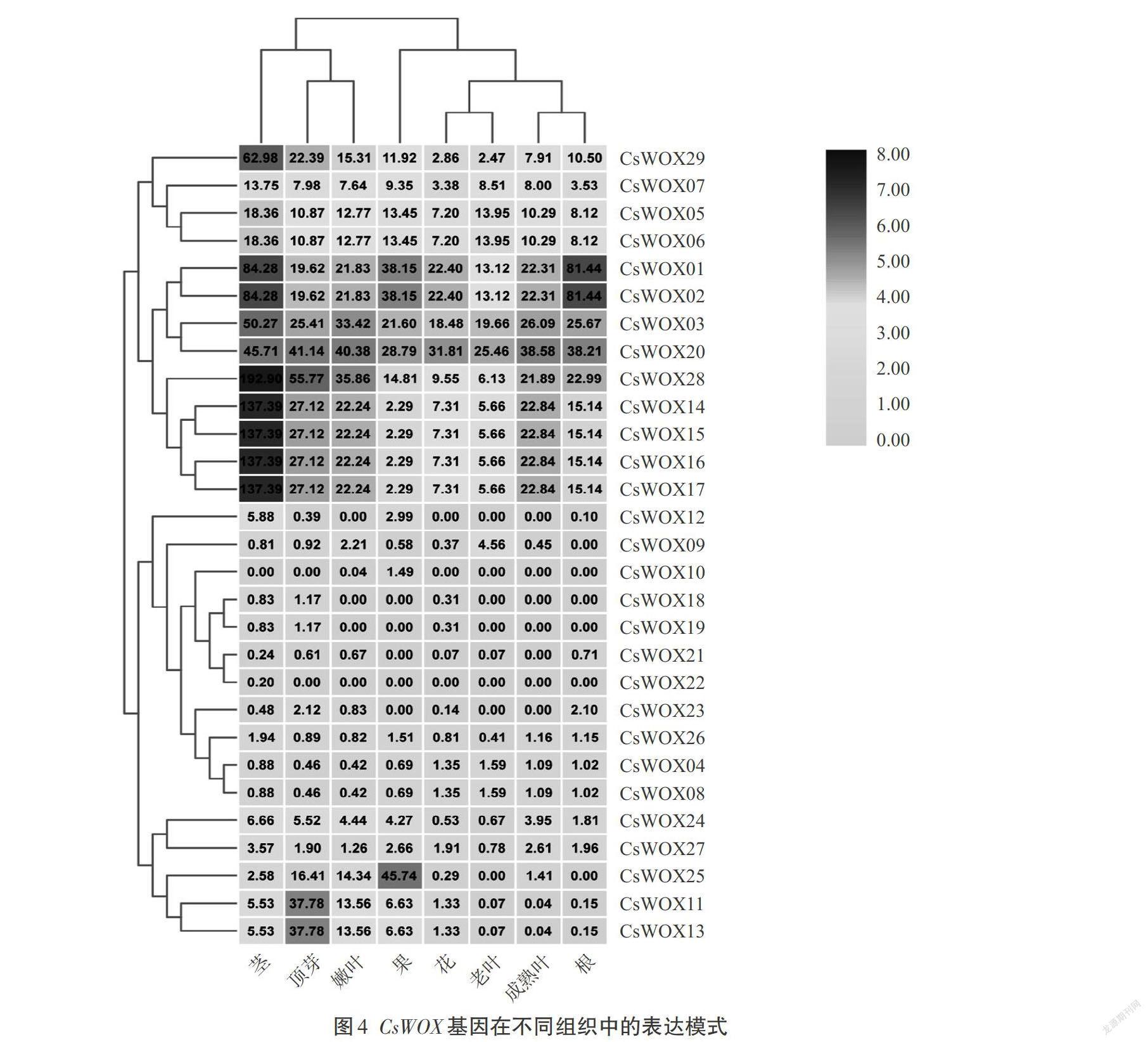
4. CsWOX基因在茶树中的不同组织表达分析
为了进一步明确CsWOX在茶树中的组织表达特异性,从TPIA中下载它们在茶树茎、顶芽、嫩叶、果、花、老叶、成熟叶、根等8个组织的转录组TPM表达数据,并对它们进行了分析。结果(图4)显示,CsWOX在多种组织中均有表达如:CsWOX01、CsWOX02、CsWOX03、CsWOX05、CsWOX06、CsWOX07、CsWOX14、CsWOX15、CsWOX16、CsWOX17、CsWOX20和CsWOX28、CsWOX29在所有组织中均具有较高的表达水平,而CsWOX04、CsWOX08、CsWOX09、CsWOX10、CsWOX12、CsWOX18、CsWOX19、CsWOX21、CsWOX23在各組织中表达较低,有部分CsWOX基因几乎不表达,如CsWOX22。相对其他组织而言,CsWOX29在茎中的表达水平总体比其他组织的要高。同时研究发现CsWOX05和CsWOX06,CsWOX01和CsWOX02,CsWOX04和CsWOX08,CsWOX11和CsWOX13,CsWOX14、CsWOX15、CsWOX16和CsWOX17在各组织中表达量一样。基因除了在根部、成熟叶、老叶表达量低,逐渐降低甚至不表达。在其他组织表达都较高的有CsWOX11、CsWOX13、CsWOX25等,其表达量在顶芽、嫩叶、果中较高。以上结果表明,不同CsWOX基因可能参与茶树的不同生长发育过程。
5. CsWOX基因在干旱胁迫、盐胁迫处理下的表达模式
对茶树CsWOX基因在PEG诱导的干旱胁迫(0、24、48、72 h)、盐胁迫处理(0、24、48、72 h)平均温度达到10 ℃以上,去驯化后的转录组数据进行分析。结果显示,在干旱胁迫下,与其他基因的表达水平相比,CsWOX01、CsWOX02、CsWOX03、CsWOX20、CsWOX28和CsWOX29的表达量较高;CsWOX04、CsWOX08、CsWOX10、CsWOX12、CsWOX18、CsWOX19、CsWOX21的表达量在干旱下被抑制,其中CsWOX03基因的表达在24 h时被上调,72 h时又下降;CsWOX21的表达水平在24 h呈上调趋势,之后被抑制;其余表达量不变。CsWOX10、CsWOX12、CsWOX18、CsWOX19、CsWOX22等基因不受干旱胁迫处理的调控(图5)。
在盐胁迫处理下,CsWOX10、CsWOX12、CsWOX18、CsWOX19、CsWOX22等基因的表达量在盐胁迫下被抑制;CsWOX01、CsWOX02、 CsWOX03、CsWOX20、CsWOX28的表达量较高,而且不受盐胁迫的调控,CsWOX21的表达水平在盐胁迫下在24 h、72 h呈上升趋势(图5)。
三、讨论
WOX基因家族在多个物种中已经被鉴定。其中高粱11个、玉米21个、水稻72个[3]、纸桑10个[7]、大豆33个[10]、小桐子12个[11],本研究在全基因组水平上一共鉴定到29个茶树CsWOX基因。CsWOX基因等电点范围为5.53(CsWOX03)~9.97(CsWOX10),有65.52%(19个)的茶树WOX基因等电点大于7,表明大部分CsWOX蛋白富含碱性氨基酸,可能会在碱性亚细胞环境发挥作用,与大豆GmWOX成员的蛋白性质基本一致。系统进化分析将15个拟南芥WOX基因、34个大豆WOX基因和29个茶树WOX基因分成了4个亚家族。其中GroupⅢ是最大的一个亚家族,包括15个茶树WOX基因,GroupⅣ是最小的一个亚家族,包括2个茶树WOX基因,符合植物中CsWOX基因在不同亚家族的分布特征。基因结构和保守基序分析发现,各亚族WOX基因之间差异很大,但相同亚组之间结构较为保守。此外,茶树WOX基因不仅在不同亚基因组中数量不同,而且在相同亚基鉴定的WOX基因数目也有所不同,说明了WOX基因在茶树进化过程种发生了基因的保留和丢失。基因复制结果说明了茶树WOX基因在茶树进化过程中发生了不同染色体间的复制,这些同源基因对在进化关系上具有良好的近缘关系。
茶树是中国乃至全球最重要的经济作物之一,干旱、盐等非生物胁迫严重影响了茶树生长发育,造成了茶叶产量和品质的大幅下滑。研究结果表明,植物WOX基因在器官形成和多条激素响应具有重要调节作用,然而WOX基因家族中还有很多调控机制尚不清楚,尤其是在木本植物中关于WOX基因家族的报道更是鲜有。本研究从基因組层面鉴定了茶树中CsWOX基因家族,分析了它们的生物信息学特征,表达模式分析显示它们可能在茶树生长发育及逆境胁迫等响应中发挥重要功能。研究结果为进一步深入了解CsWOX基因参与茶树生长发育、茶叶品质成分变化及茶树的器官诱导作用机制奠定基础,但茶树WOX基因对激素和其他逆境胁迫响应的具体机制尚不明确,需要进一步研究。
参考文献
[1] ZHANG X, ZONG J, LIU J, et al. Genome-wide analysis of WOXgene family in rice, sorghum, maize, arabidopsis and poplar[J]. Journal of Integrative Plant Biology, 2010(11): 1016-1026.
[2] HUI W, NIU L F, FU C X, et al. Overexpression of the WOX gene stenofolia improves biomass yield and sugar release in transgenic grasses and display altered cytokinin homeostasis[J]. Plos Genetics, 2017, 13(3): e1006649.
[3] ZHAO Y, HU Y F, DAI M Q, et al. The wuschel-related homeobox gene WOX11 is required to activate shoot-borne crown root development in rice[J]. The Plant Cell, 2009, 21(3): 736-748.
[4] JIANG W, ZHOU S L, ZHANG Q, et al. Transcriptional regulatory network of WOX11 is involved in the control of crown root development, cytokinin signals, and redox in rice[J]. Journal of Experimental Botany, 2017, 68(11): 2787-2798.
[5] SARKAR A K, MARIJN L, SHUNSUKE M, et al. Conserved factors regulate signalling in Arabidopsis thaliana shoot and root stem cell organizers[J]. Nature, 2007, 446(7137): 811-814.
[6] MAIDA R B, JUAN J R, YANOFSKY M F, et al. The WOX13 homeobox gene promotes replum formation in the Arabidopsis thaliana fruit[J]. The Plant Journal, 2013, 73(1): 352-356.
[7] FENG T, CHEN N Z, ZHAO M L, et al. Identification and functional divergence analysis of WOX gene family in paper mulberry[J]. International Journal of Molecular Sciences, 2017, 18(8): 1782-1786.
[8] YANG Z, GONG Q, QIN W Q, et al. Genome-wide analysis of WOX genes in upland cotton and their expression pattern under different stresses[J]. Bmc Plant Biology, 2017, 17(1): 113-119.
[9] 邢光伟, 王梦醒, 马小飞, 等. 小麦LBD基因家族的全基因组鉴定,表达特性及调控网络分析[J]. 麦类作物学报, 2017, 37(7): 855-863.
[10] HAO Q, ZHANG L, YANG Y, et al. Genome-wide analysis of the WOX gene family and function exploration of GmWOX18 in soybean[J]. Plants (Basel, Switzerland), 2019, 8(7): 1134-1139.
[11] 唐跃辉, 包欣欣, 王健, 等. 小桐子WOX基因家族全基因组鉴定与表达分析[J]. 分子植物育种, 2019, 17(4): 1154-1162.
