锰胁迫下鸡眼草的富集特征及生理响应
2021-07-19李欣航肖泽华匡雪韶王悟敏罗亮宇刘文胜
李欣航,肖泽华,匡雪韶,王悟敏,罗亮宇,刘文胜
(中南林业科技大学生命科学与技术学院,湖南长沙410004)
锰(Mn)是植物生长发育必需的微量营养元素,作为代谢酶的组分参与许多生物化学反应[1]。过量的锰会抑制植物生长,甚至可通过食物链危害人体健康[2]。然而,由于长期不合理的矿山开采、冶金以及锰肥等富锰物质的滥用,我国许多地区土壤锰污染日益严重。因此,锰污染区的生态修复与治理已成为刻不容缓的任务。
近年来,人们已采用多种方式开展矿区治理。植物修复因具有环境友好和经济实惠等特点被广泛关注,成为矿区污染治理的重要方式[3]。开展植物修复的关键是筛选一批耐锰性强、经济美观的植物或生态型,并揭示它们的锰耐受性机制。生长在重金属污染土壤上的植物,由于长期经受高重金属浓度的选择压力,逐渐适应该环境;其耐受重金属胁迫的能力较生长在非污染土壤上的植物强,进而形成了不同的生态型[4]。因而,重金属污染区植物具有与非污染区植物不同的重金属耐受性。开展两种来源植物重金属耐受性特征的比较,是筛选植物修复材料并揭示其耐受机制的重要途径。
植物生长、生理生化特征和重金属富集能力均为衡量植物对重金属耐受性的重要指标,可反映植物对重金属胁迫的适应能力。例如,刘玉通等[5]对比研究不同生态型蜈蚣草(Pteris vittata)在铅(Pd)胁迫下的表型特征,结果发现矿区生态型蜈蚣草的生物量受胁迫影响较小,比非矿区生态型蜈蚣草有更强的铅耐受性;胡国涛等[6]通过研究两个品种速生树种竹柳(Salixsp.)对重金属的生理反应,发现在重金属胁迫作用下竹柳五号可维持较高的可溶性糖、可溶性蛋白含量和过氧化物酶、超氧化歧化酶活性,得出竹柳五号比竹柳三号对重金属胁迫有更强的耐受性;王一峰等[7]比较研究了污染区与非污染区的乌苏里风毛菊(Saussurea ussuriensis)的叶绿素含量、抗氧化酶系的酶活性、丙二醛含量、可溶性蛋白含量等生理生化特征,发现污染区乌苏里风毛菊这些指标受胁迫变化较非污染区的小,表明其已经产生了适应性抗性进化。Zambrosia 等[8]通过对不同锰浓度下生长的6 个甘蔗(Saccharumspp.)品种的研究表明,耐性品种植物组织的锰含量与敏感品种相当或更高,植物组织锰含量是锰耐受性的重要指标。Ding 等[3]对两个红麻(Hibiscus cannabinus)品种在铬(Cr)胁迫下的表型、生理生化与富集特征进行了比较研究,结果显示随着铬浓度升高,植物体内重金属含量增加,这些指标均可反映其耐受性。刘攀道等[9]通过研究对比两种基因型圭亚那柱花草(Stylosanthes guianensis)在不同Mn2+浓度下的表型、生理生化及Mn积累特性,发现维持根系生长、降低植物组织锰含量和提高抗氧化酶活性是苗期柱花草TF226 耐锰的生理机制。这些研究说明植物生长、生理生化特征和富集特性可较好地揭示植物耐受性机制。
鸡眼草(Kummerowia striata)为豆科一年生草本植物,具有生态适应性广、繁殖能力强和能进行生物固氮等优点;还是一种优良的园林地被植物和重要中草药[10],值得进一步开发利用。鸡眼草在锰矿废弃地的高锰含量地段生长繁茂,这表明该植物对锰具有较强耐受性,是一种兼性金属型植物,也是一种理想的生物修复备选植物。近年来已有许多学者开展了鸡眼草的种子萌发[10]、生物学特征[11]和水分胁迫响应[12]等方面的研究,这些研究表明了鸡眼草具有较强的耐胁迫能力。本研究以污染区和非污染区鸡眼草为材料,研究不同浓度Mn 胁迫下,两种来源鸡眼草的生长、重金属累积特征及生理生化反应的变化规律,以期揭示鸡眼草对锰胁迫的耐受机制,研究结果也可用于矿山恢复实践。
1 材料与方法
1.1 种子采集及盆栽试验
供试污染区鸡眼草种子于2017 年11 月中旬采自湖南省湘潭市锰矿区(北纬28°03′,东经112°55′),该地段土壤锰含量达 17990.21 mg·kg−1(湖南省土壤锰含量平均值为 459 mg·kg−1[13])。非污染区鸡眼草种子于 2017 年 11月下旬采自位于湖南省长沙市的中南林业科技大学校园内,其土壤锰含量达138.25 mg·kg−1。现场调查显示这两个地段鸡眼草的分布面积均超过1 hm2,长势较好,且均为自然生长。在两个地段各选择100 株长势一致、果实已成熟的鸡眼草植株,采集其地上部分并用信封装好后带回实验室分离萌发所需种子。
分别选取两种来源籽粒饱满、大小均匀的鸡眼草种子进行种子萌发。用0.5% NaClO 溶液将种子浸泡消毒30 min,然后用去离子水冲洗3~4 次后,最后将种子分别放置于干净沙子中。根据预实验结果及参照吴哲等[10]的结果,种子萌发在25 ℃、12 h 光照/12 h 黑暗、相对湿度80%的人工气候箱中进行。待到鸡眼草生长出5~6 片真叶时移栽至温室大棚的花盆中继续培养。
通过预备试验以及参考潘高等[14]的研究,鸡眼草的盆栽试验在温室大棚中开展,利用MnCl2·4H2O 设置0(对照)、1000、5000、10000、15000、20000 μmol·L−16 个锰浓度梯度。两种来源的鸡眼草每个浓度各设置 10 个花盆,每个花盆7 株幼苗,共设置120 个花盆、840 株幼苗。每个塑料花盆(直径18 cm)中装入2 kg 按1∶1 混合的珍珠岩和细沙。日常浇水,防病虫害,每周浇一次Hoagland’s 营养液,保证植物正常生长。待幼苗在花盆中适应生长15 d 后,加入相应浓度的MnCl2溶液。在胁迫第30 天时,收获试验材料,测量鸡眼草生长、生理生化和富集等相关指标。生长相关指标包括芽长、根长、鲜重和干重;生理生化指标包括可溶性糖、可溶性蛋白、脯氨酸和叶绿素含量及超氧化物歧化酶(superoxide dismutase,SOD)、过氧化物酶(peroxidase,POD)活性和丙二醛(malondialdehyde,MDA)含量;富集相关指标包括根、茎和叶各部分的Mn 含量。
1.2 测定项目及方法
1.2.1 生长指标的测定 植株芽长和根长的测定:污染区和非污染区鸡眼草的每个处理随机选取15 株植株,采用精度为1 mm 的直尺分别测量幼苗芽长、根长等指标。
植株鲜重和干重的测定:将鸡眼草植株分为地上(芽)与地下(根)两个部分,分别称量鲜重,然后再于95 ℃下杀青10 min,65 ℃下烘干至恒重后称重。

1.2.2 生理生化指标的测定 随机选取污染区和非污染区鸡眼草幼苗叶片,测定生理生化指标。采用蒽酮比色法测量可溶性糖(soluble sugar,SS)含量;采用考马斯亮蓝G-250 染色法测量可溶性蛋白(soluble protein,SP)含量;采用酸性茚三酮比色法测量脯氨酸含量;采用氮蓝四唑法测量超氧化物歧化酶活性;采用愈创木酚法测量过氧化物酶活性;采用硫代巴比妥酸比色法测量丙二醛含量[16]。以上各指标均分别称取污染区和非污染鸡眼草新鲜叶片各0.1 g,重复3 次。
1.3 富集指标的测定
收获的样品先用去离子水冲洗干净,再将根浸入20 mmol·L−1EDTA-2Na 溶液中交换15 min,以去除表面吸附的金属离子,最后再用去离子水冲洗干净,吸干植株表面的水分。将样品分为根、茎、叶,测定各部分鲜重。将新鲜样品放在95 ℃的烘箱内保持30 min,然后再在65 ℃下烘48 h,测定植物各部分干重,最后用不锈钢粉碎机磨细,过2 mm 尼龙网筛,供分析测定。
采用湿法消解法[V(HNO3)∶V(HClO4)=3∶1][17]将植物材料进行消解至澄清,消解液用去离子水定容至 50 mL,用火焰原子吸收分光光度计(北京普析,TAS-990 型)测定Mn 含量。转运系数反映植物吸收重金属后从根部转移到地上部分的能力,转运系数(translocation factor,TF)为鸡眼草地上部分与根部Mn 含量的比值。
1.4 数据处理
采用SPSS 21.0 软件对两种来源鸡眼草的生长指标、生理指标、富集指标与胁迫浓度进行双因素方差分析,并对每个指标不同处理之间的差异显著性使用LSD 法进行多重比较。比较的数据结果均以“平均值±标准差”表示。用SigmaPlot 12.5 作图。
2 结果与分析
2.1 锰胁迫对植物生长的影响
随着锰浓度的升高,两种来源鸡眼草的根长、芽干重、根干重、根冠比均逐渐降低,芽长则呈先升后降的趋势(表1)。当锰浓度达5000~20000 μmol·L−1时,与对照相比,污染区鸡眼草的芽干重下降幅度为4.34%~27.71%,根干重下降幅度为19.00%~66.06%;非污染区鸡眼草芽干重下降幅度为16.33%~49.77%,根干重下降幅度为27.90%~77.54%。两种来源鸡眼草在对照及1000 μmol·L−1时芽干重、根干重和根冠比无显著差异;当锰浓度达5000 μmol·L−1及以上时,污染区鸡眼草的根干重则均显著高于非污染区。这说明鸡眼草的生长受高锰胁迫抑制,且在高锰浓度时,污染区鸡眼草受到抑制程度较低。
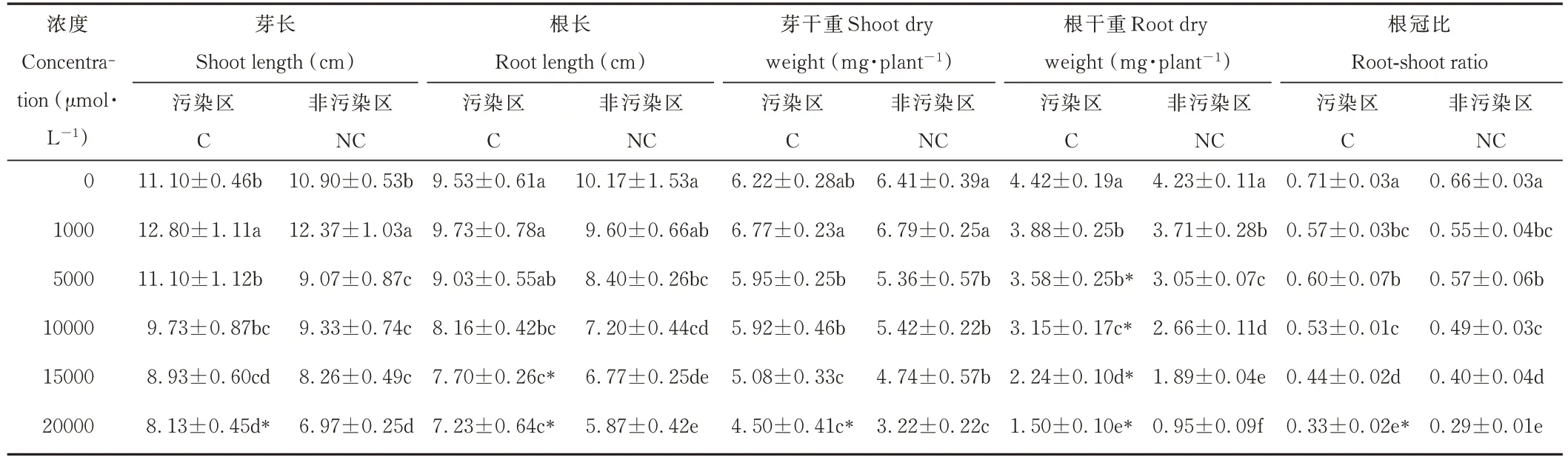
表1 锰胁迫对两种来源鸡眼草生长的影响Table 1 Effect of Mn stress on K.striata growth from two sources
2.2 锰胁迫对植物生理生化特征的影响
2.2.1 叶绿素和渗透调节物质含量 双因素方差分析结果显示,植物来源与锰浓度对叶绿素、可溶性糖、可溶性蛋白和脯氨酸含量均有极显著影响,且两者交互作用极显著(图1)。随着锰浓度的升高,两种来源鸡眼草的叶绿素、可溶性糖和脯氨酸含量均呈先升后降的趋势,可溶性蛋白的含量则呈逐渐下降的趋势。锰浓度为1000 μmol·L−1时,两种来源鸡眼草的叶绿素含量达最大值;相同锰浓度下污染区鸡眼草的叶绿素含量均显著高于非污染区鸡眼草。锰浓度为5000 μmol·L−1时,两种来源鸡眼草的可溶性糖和脯氨酸含量均达到最大值;当锰浓度高于5000 μmol·L−1时,污染区鸡眼草可溶性糖和脯氨酸含量则均显著高于非污染区。锰浓度低于1000 μmol·L−1时,污染区与非污染区鸡眼草的可溶性蛋白含量无显著差异;当锰浓度高于5000 μmol·L−1时,污染区鸡眼草可溶性蛋白含量均显著高于非污染区。这说明锰浓度高于5000 μmol·L−1时,污染区鸡眼草叶绿素和渗透调节物质含量受到的影响小于非污染区。
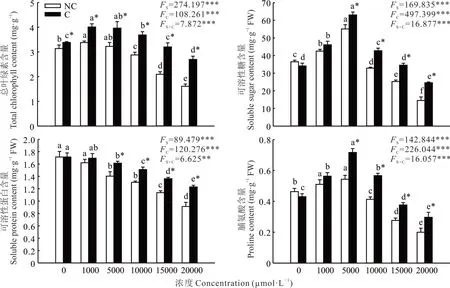
图1 锰胁迫对两种来源鸡眼草生长过程中总叶绿素、可溶性糖、可溶性蛋白和脯氨酸含量的影响Fig.1 Effect of Mn stress on the total chlorophyll,soluble sugar,soluble protein and proline contents of leaves from two sources of K.striata
2.2.2 过氧化物酶、超氧化物歧化酶活性和MDA 含量 双因素方差分析结果表明,锰浓度和植物来源对SOD、POD 酶活性和MDA 含量均有极显著影响,且二者有极显著的交互作用(图2)。两种来源鸡眼草叶片的POD、SOD 活性随锰浓度的升高呈先升后降的变化趋势,且锰浓度高于5000 μmol·L−1时,污染区鸡眼草的POD、SOD 活性均显著高于非污染区;与对照相比,污染区鸡眼草的SOD、POD 活性的下降程度低于非污染区。这说明锰浓度高于5000 μmol·L−1时,污染区鸡眼草的SOD、POD 活性受到的抑制程度低于非污染区。
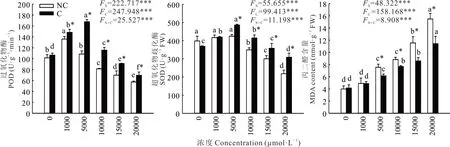
图2 锰胁迫对两种来源鸡眼草过氧化物酶、超氧化物歧化酶活性和丙二醛含量的影响Fig. 2 Effects of Mn stress on activities of peroxidase,superoxide dismutase and malondialdehyde content in two sources of K.striata
随着锰浓度的升高,两种来源鸡眼草叶片的MDA 含量均逐渐升高。当锰浓度小于1000 μmol·L−1时,两种来源鸡眼草的MDA 含量差异不显著。锰浓度高于5000 μmol·L−1时,污染区鸡眼草的MDA 含量则显著低于非污染区(图2)。
2.3 锰胁迫对植物组织锰含量的影响
由表2 可知,鸡眼草各器官锰含量高低顺序为:叶>根>茎。随着锰浓度的升高,污染区和非污染区鸡眼草的根、茎、叶中的锰含量均显著增加;当锰浓度达20000 μmol·L−1时,污染区和非污染区鸡眼草叶的锰含量分别达到了12553.03 和8558.05 mg·kg−1,为各自对照的16.53 和13.41 倍。两种来源鸡眼草相比,低浓度时污染区与非污染区鸡眼草的根、茎、叶锰含量无显著差异;高锰浓度时污染区鸡眼草的根、茎、叶锰含量则均高于非污染区。污染区和非污染区鸡眼草的转运系数随着锰浓度的升高而增加,污染区和非污染区鸡眼草的转运系数则无显著差异。
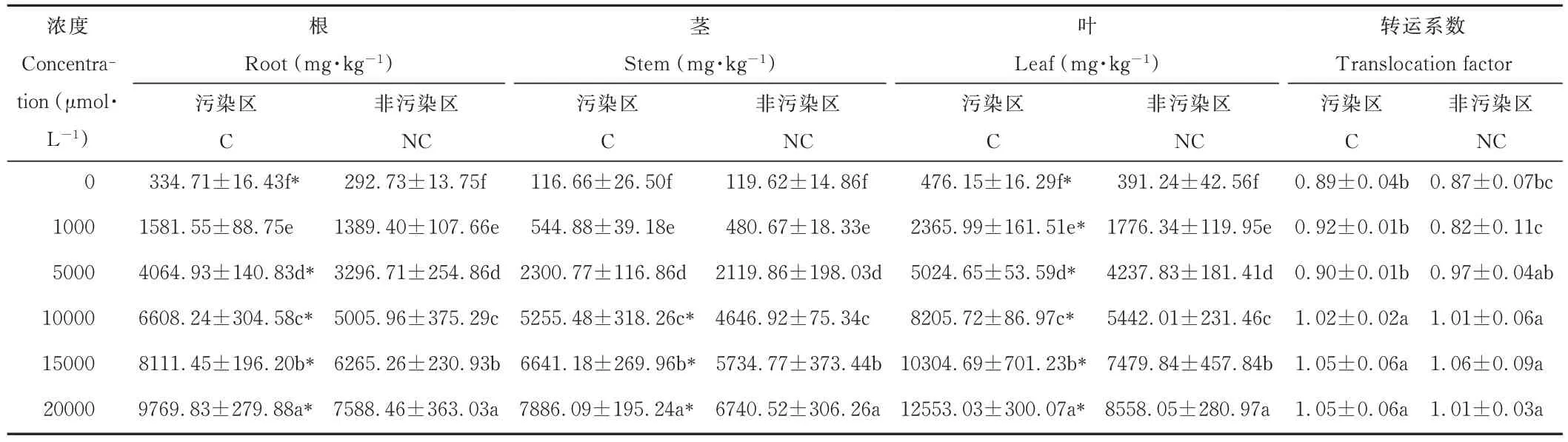
表2 锰胁迫下两种来源鸡眼草根、茎、叶的Mn 含量Table 2 Mn content of roots,stems and leaves from two sources of K.striata under Mn stress
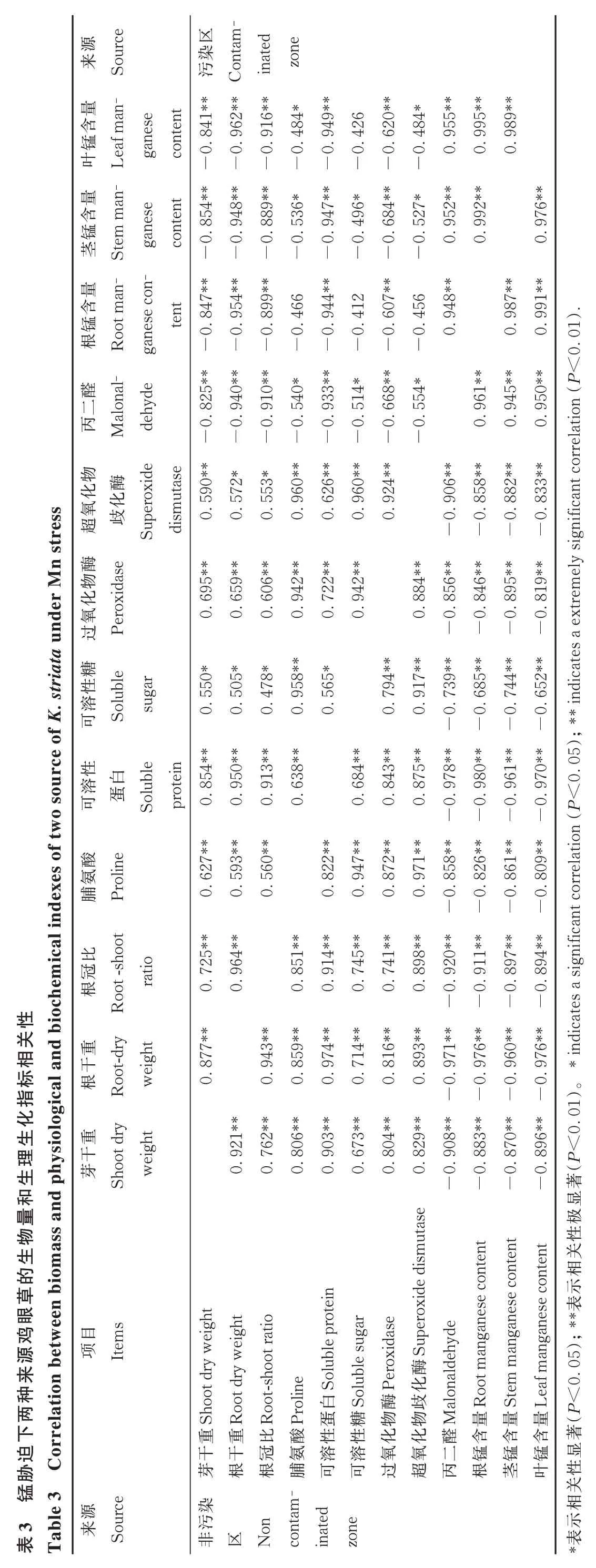
2.4 各指标的相关性
由表3 可知,两种来源鸡眼草的丙二醛含量、根锰含量、茎锰含量、叶锰含量与根干重、芽干重、根冠比、脯氨酸含量、可溶性蛋白含量、可溶性糖含量、SOD 活性、POD 活性之间均呈显著负相关关系。两种来源鸡眼草的丙二醛含量、根锰含量、茎锰含量、叶锰含量两两之间均呈显著正相关关系。两种来源鸡眼草的根干重、芽干重、根冠比、脯氨酸含量、可溶性蛋白含量、可溶性糖含量、SOD 活性、POD 活性两两之间均呈显著正相关关系。
3 讨论
3.1 生物量的变化
植物在逆境下的生长情况是植物对该环境适应性的最直接反映[18]。本研究结果显示,随着锰浓度的升高,两种来源鸡眼草的芽干重、根干重均逐渐降低,污染区鸡眼草的芽干重、根干重的下降幅度低于非污染区鸡眼草。Ding 等[3]、刘玉通等[5]、Zambrosia 等[8]的研究也发现随着重金属浓度的升高,植物生物量降低,耐性较强的基因型(或生态型)生物量受重金属胁迫影响较低,本研究结果与此基本一致。这说明高浓度锰胁迫抑制鸡眼草生长,污染区较非污染区鸡眼草具有更强的锰耐受性。其原因可能是污染区鸡眼草在长期处于高锰胁迫环境条件下,逐渐适应立地环境,已产生较强的锰耐受性。
3.2 生理生化特征的变化
3.2.1 叶绿素和渗透调节物质含量 叶绿素是植物光合作用的基础物质,其含量反映了植物光合能力的高低。本研究结果显示,随着锰浓度的升高,两种来源鸡眼草叶绿素含量均呈先上升后下降的趋势,且同浓度下污染区鸡眼草的叶绿素含量高于非污染区,这与刘玉通等[5]、刘攀道等[9]、王一峰等[7]对重金属胁迫下不同生态型对比的结果一致。这说明两种来源鸡眼草在高浓度锰胁迫时,其叶绿素均受到抑制;污染区鸡眼草具有更强的耐受性。其原因是当锰浓度过高时,高浓度的锰会破坏叶绿体的结构,导致叶绿素合成受阻,从而降低了叶绿素的含量及光合作用能力;而污染区鸡眼草可能已产生特殊机制来提高锰的耐受性。
生物体在代谢过程中,可通过有机渗透调节物质的积累和分解来调节细胞渗透平衡,以缓解重金属胁迫导致的伤害[19]。本研究结果表明,随着锰浓度的升高,两种来源鸡眼草幼苗的可溶性糖和脯氨酸含量呈先升高后下降的趋势,可溶性蛋白含量呈逐渐下降的趋势;当锰浓度高于5000 μmol·L−1时,污染区鸡眼草的可溶性糖、可溶性蛋白和脯氨酸含量的下降程度均低于非污染区。这与刘攀道等[9]、王一峰等[7]、潘高等[14]的研究结果一致。其原因是锰作为植物的必需元素,低浓度的锰可以促进植物体内所需物质的合成,这可能使植物体内的糖和脯氨酸的含量增加;过量的锰则会使植物受到锰毒害,导致植物体内可溶性糖和脯氨酸的合成受阻,植物体内的可溶性蛋白、可溶性糖和脯氨酸含量降低。在锰胁迫条件下,污染区鸡眼草的耐受性高于非污染区,可能是因为长期在污染区生长的鸡眼草已经产生了对锰的适应性抗性进化,对锰产生了一定的抗性。
3.2.2 SOD、POD 活性和 MDA 含量 植物长期生长在逆境环境中,为了适应环境而进化出抗氧化保护酶系统。其中,SOD 是植物清除活性氧的重要物质,它可以将O−快速歧化为H2O2和O2;而 POD 则可将 H2O2进一步分解成 H2O 和 O2,从而达到植物的自我保护作用[20]。本研究中,随着锰浓度的升高,两种来源鸡眼草叶片的SOD、POD 活性呈先升后降的变化趋势,且污染区鸡眼草的POD、SOD 活性变化幅度低于非污染区,这与刘玉通等[5]、王一峰等[7]的研究结果一致。说明在锰胁迫环境下,抗氧化酶的去氧化是植物重要的防御反应,使植物对逆境胁迫具有一定的调节能力;较强的去氧化能力,加强了活性氧的清除能力,使植物具有较强的重金属耐受能力;而在高浓度重金属胁迫下,抗氧化保护酶活性受到抑制,过高浓度的重金属使植物抗氧化体系遭到破坏,从而削弱了其保护能力,造成对植物体的损伤[21]。另一方面,本研究也说明污染区鸡眼草具有较强的去氧化能力,其对逆境的自我调节能力高于非污染区。其原因可能是污染区生长的鸡眼草已经产生了对锰的适应性抗性进化,具有较强的抗氧化保护酶系统,是污染区鸡眼草具有较强锰耐受性的重要生理机制。
?
植物在正常生长情况下,体内的活性氧物质会处于平衡状态。当植物受到外界胁迫时,这种平衡被打破,导致植物体产生过量的活性氧物质。当植物体吸收一定量的重金属后,会使植物组织中的重金属含量增加,引起植物膜脂过氧化,MDA 含量升高,从而打破植物体内活性氧物质的平衡。本研究中,两种来源鸡眼草的MDA 含量均随锰浓度的升高逐渐上升,同浓度下污染区鸡眼草的MDA 含量显著低于非污染区。说明污染区鸡眼草在锰胁迫下表现出更强的抵抗能力,受到的伤害更小。这与赵建桥等[22]对铅胁迫下2 种生态型五节芒(Miscanthus floridulus)的研究及王一峰等[7]对铅胁迫下污染区与非污染区乌苏里风毛菊研究中MDA 含量变化的结果相似。其原因可能是因为长期在污染区生长的鸡眼草已经产生了对锰的适应性抗性进化,形成了对锰的抗性居群,因此在低锰浓度下产生的MDA 少于非污染区鸡眼草。
3.3 锰胁迫下鸡眼草的锰累积特征
本研究结果显示,污染区和非污染区鸡眼草根、茎、叶中的锰含量均随着锰浓度的升高而增加。Ding 等[3]对铬胁迫下红麻体内铬含量的研究及陈小米等[23]对镉和锌复合污染条件下竹柳重金属含量的研究结果均显示,植物体内重金属含量随着环境重金属浓度的增加而增加。本研究结果与此一致,说明鸡眼草具有较强的锰富集能力;植物体内重金属含量随环境重金属浓度的增加而增加是一个普遍规律。本研究还显示,相同浓度锰胁迫下,污染区鸡眼草根、茎、叶的锰含量均高于非污染区。这与Gan 等[24]对铜(Cu)胁迫下长萼鸡眼草(Kummerowia stipulacea)的研究、Pan 等[25]对锰胁迫下污染区与非污染区苍耳(Xanthium strumarium)的研究结果一致。这说明污染区鸡眼草对锰的吸收与富集能力高于非污染区。其原因可能是鸡眼草长期生长在矿区高锰浓度的环境条件下,其种群产生了适应性遗传分化,已进化形成了相应的能在体内累积较高浓度的锰而不受伤害的耐受性机制,例如螯合机制等。需要有更多的研究来验证并揭示相关机制。
两种来源鸡眼草在锰浓度20000 μmol·L−1的胁迫条件下仍能生长,且污染区和非污染区鸡眼草的叶片中的锰含量分别达到了12553.03 和8558.05 mg·kg−1,这说明鸡眼草对锰具有较强的富集能力。鸡眼草具有耐不良生长环境、繁殖能力强、生长迅速等特点,是一种非常好的园林地被植物。同种锰浓度下,污染区鸡眼草植株内锰的含量显著高于非污染区,所以利用污染区鸡眼草是矿区土壤Mn 污染生态修复的一种较好的选择。
4 结论
随着锰浓度的升高,两种来源鸡眼草的芽干重、根干重均逐渐降低,污染区鸡眼草降低幅度低于非污染区鸡眼草。随着锰浓度的升高,两种来源鸡眼草的叶绿素、可溶性糖和脯氨酸含量、SOD、POD 活性均呈先升后降的趋势,可溶性蛋白含量逐渐下降,MDA 含量逐渐升高。随着锰浓度的升高,污染区和非污染区鸡眼草根、茎、叶中的锰含量均增加,污染区鸡眼草较非污染区锰富集能力强。污染区鸡眼草锰耐受能力及富集能力均高于非污染区,提高抗氧化酶活性是其重要的生理机制。
