野生大麦和栽培大麦籽粒性状及蛋白质含量的多样性分析
2021-06-08卢广周羽沈辉任喜峰
卢广,周羽,沈辉,任喜峰
1.华中农业大学植物科学技术学院,武汉 430070; 2.湖北省襄阳市原种繁殖场,襄阳 441004
大麦作为全球栽培的第四大禾谷类作物,在我国种植历史悠久,用途极其广泛,是集食用、饲用及啤酒酿造于一体的重要农作物[1]。大麦籽粒大小和千粒重是大麦产量和品质的重要组成部分[2-3]。籽粒大小和千粒重与大麦的生长环境、选育水平以及种质的遗传背景均有较大的相关性[4]。不同棱型大麦品种千粒重有很大差异,二棱普遍高于六棱(或多棱型),且同棱型的籽粒大小也因品种不同而异[5-6]。陈晓东等[7]将125份大麦材料按照棱型、皮裸以及选育水平进行分类,对粒重、粒长、粒宽与粒厚进行差异比较与相关性分析,发现大麦粒重与棱型相关,与皮和裸无关,二棱大麦的粒重、粒长和粒宽均高于六棱大麦。张新忠等[8]在不同试点对98份二棱大麦和89份六棱大麦品种的千粒重、粒长、粒宽进行测定和分析,结果表明,六棱大麦的千粒重、粒长和粒宽较二棱大麦普遍偏低;不同棱型大麦千粒重、粒长和粒宽在基因型间及环境间差异均达到显著或极显著水平。关于野生大麦不同分类及其籽粒性状的差异性研究鲜有报道。
据统计,全球大约95%的大麦都用于饲用或啤酒酿造[9],蛋白质含量(grain protein content,GPC)是谷物品质评价中的一个关键性指标[10]。GPC与大麦饲用质量和麦芽品质均密切相关,不同GPC的大麦品种具有不同的用途,高GPC的大麦品种通常作为食用或饲用,低GPC的大麦品种则适于麦芽生产和啤酒酿造[11-12]。因此,针对不同用途大麦的遗传改良,研究和选育不同GPC的大麦品种是十分必要的。产量与品质的提升均是作物育种的主要目标,然而,在很多作物中GPC与产量呈现显著的负相关,即在提高产量的同时提高GPC是相对困难的[13]。种质资源的遗传多样性是作物育种突破的关键[14],具有丰富遗传多样性的野生大麦则可以作为改良栽培大麦产量和GPC的重要种质资源。大麦籽粒性状的鉴定分析是大麦种质资源评价中较为直观、简单有效的方法。种质资源表型多样性研究中运用较为广泛的方法有表型的差异分析、相关性分析、聚类分析等。栽培大麦经过不断地人工选育使其遗传多样性大幅度降低,导致亲本遗传基础狭窄,增加了对病虫害的脆弱性。这些特征已成为当代大麦育种面临的主要困境[15]。本研究以214份来自国内外的不同类型大麦种质为材料,对其籽粒性状进行差异比较分析和相关性分析,以期为不同类型大麦品种的粒型选择与品质改良提供参考依据;同时,对筛选创造优异种质资源、有效利用大麦野生种质具有重要的意义。
1 材料与方法
1.1 试验材料
本试验选用的214份大麦种质资源包括来自西南亚、中亚和西藏起源地的97份野生大麦和来自世界不同区域的117份栽培大麦。根据多年表型检测和遗传进化分析[16-19],该供试材料遗传多样性非常丰富,是遗传改良的优异基因资源。214份材料分别于2018年和2019年连续2 a种植于华中农业大学小麦试验基地,按小区种植,每份材料3行,行长1.0 m,行距0.2 m,各材料按照顺序编号播种,3次重复,田间管理与大田一致。分别于次年5月收获种子,收获的种子晒干后置于4 ℃冷库保存。117份栽培大麦按棱型特征可分为58份二棱大麦和59份六棱大麦;按皮裸特征可分为96份皮大麦和21份裸大麦;按来源地可分为6个不同的群体,分别为59份东亚栽培大麦、5份澳大利亚栽培大麦、9份地中海沿岸栽培大麦、21份欧洲栽培大麦、16份北美栽培大麦和7份南美栽培大麦。97份野生大麦按棱型可分为87份二棱大麦和10份六棱大麦;按皮裸特征可分为93份皮大麦和4份裸大麦;按地理来源可分为3个群体,分别为20份西藏野生大麦、55份西南亚野大麦和22份中亚野生大麦。
1.2 籽粒表型性状的测定
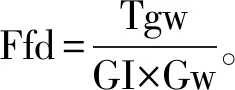
1.3 籽粒蛋白质含量的测定
每份材料分别随机取10粒饱满且无病虫害的种子装入袋内,置于70~80 ℃烘干至恒质量。烘干后的样品充分粉碎、过0.5 mm筛。为缩短检测时间和提高准确度,采用改进的微量凯氏定氮法对材料中氮含量进行测定[20],每个材料3次重复取其平均值。蛋白质转换系数为6.25[21]。
1.4 数据统计分析
籽粒性状录入Excle表中,对214份大麦材料按照不同分类方式对各个材料进行编号汇总;使用SPSS 24.0软件对2018年和2019年籽粒性状分别进行描述性统计分析;进一步将所有材料按照不同的分组进行各个籽粒性状的单因素方差分析或t检验,依据检验结果差异显著性(0.05水平)分别对各个性状进行两两比较并标注。用SPSS 24.0软件,采用Pearson相关系数法(双尾检验)计算各个性状之间的相关性以及相关性的显著性;对不同分类中GPC、千粒重、粒长、粒宽与籽粒直径5个籽粒性状的相关性同样选用Pearson相关系数法。利用R语言factoextra软件包和R自带的hclust函数基于10个籽粒性状对214份大麦材料进行层次聚类。用factoextra软件包中的fviz_dend函数进行聚类树可视化。
2 结果与分析
2.1 籽粒表型性状的描述性统计分析
对供试材料9个籽粒性状的表型数据进行统计分析(表1),结果显示:这些性状在2 a中均表现出丰富的变异(变异系数为8.70%~23.88%)。变异系数在2 a 均表现最高的性状是千粒重,其中,2018年千粒重的变异系数为23.88%,其值的变化范围为14.57~58.83 g;2019年千粒重的变异系数为23.68%,其值的变化范围为14.82~55.66 g。这表明大麦种质资源在千粒重性状上存在较大差异,具有丰富的变异类型和广泛的选择基础,品种改良潜力较大。变异系数在2 a中均表现最低的性状是籽粒直径,其中,2018年籽粒直径的变异系数为8.87%,其值的变化范围为4.01~6.41 mm;2019年籽粒直径的变异系数为8.70%,其值的变异范围为3.91~6.39 mm。2 a间各个籽粒性状的偏度和峰度的绝对值均小于1,说明所考察的各性状的表型数据基本符合正态分布。9个籽粒性状在2 a间进行独立样本t检验的结果均未表现出明显的差异,表明籽粒表型变异在供试2 a间受环境因素的影响较小。
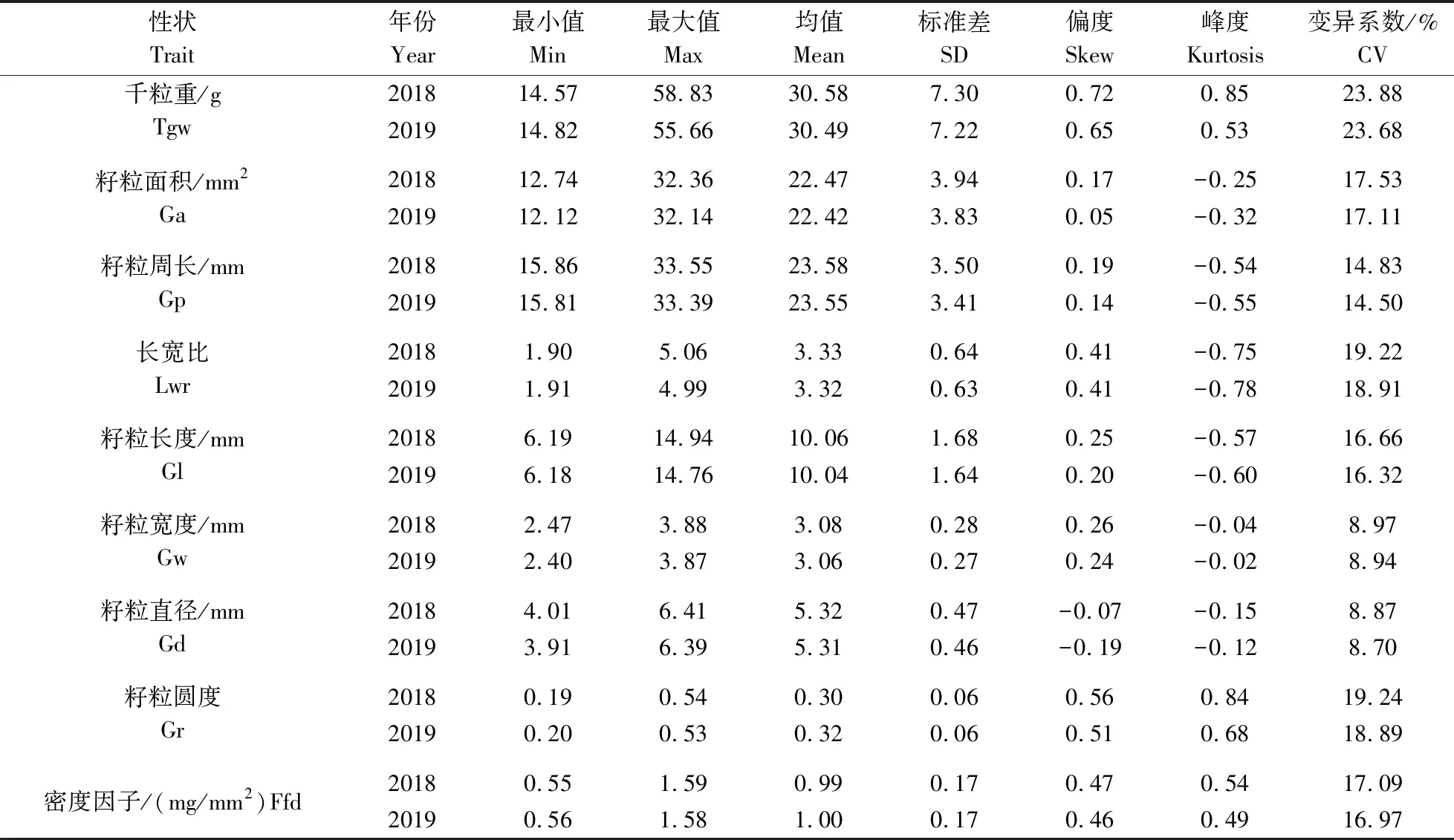
表1 大麦9个籽粒性状的描述统计Table 1 Descriptive statistics of 9 grain traits in barley
2.2 野生大麦与栽培大麦籽粒性状比较分析
由表2和表3可知,野生大麦的GPC(10.89%)显著高于栽培大麦(9.06%),且野生大麦GPC的变异系数(25.22%)和标准差(2.75)也要高于栽培大麦GPC的变异系数(20.16%)和标准差(1.85)。说明野生大麦GPC不仅表现出更高的水平,而且拥有更丰富的遗传多样性。栽培大麦和野生大麦的平均千粒重分别为31.17和30.06 g,差异不显著(表2)。但无论是栽培大麦还是野生大麦千粒重分布范围均较为广泛,其中栽培大麦千粒重的变化范围为14.57~ 58.83 g,野生大麦千粒重的变化范围为16.05~ 49.11 g,且二者均有较高的变异系数(表3)。在其他表型性状方面,野生大麦的籽粒面积(24.61 mm2)、周长(26.19 mm)及长宽比(3.84)均显著高于栽培大麦的籽粒面积(20.65 mm2)、周长(21.38 mm)与长宽比(2.89);野生大麦的粒长(11.36 mm)与籽粒直径(5.57 mm)均分别显著大于栽培大麦粒长(8.97 mm)与籽粒直径(5.10 mm);栽培大麦粒宽(3.12 mm)、籽粒圆度(0.34)与密度因子(1.06 mg/mm2)均分别显著大于野生大麦粒宽(2.99 mm)、籽粒圆度(0.26)及密度因子(0.92 mg/mm2)(表2)。结果表明,经过长期的人工驯化与改良,大麦GPC明显降低,籽粒面积、籽粒周长、长宽比、粒长与籽粒直径明显减小,粒宽、籽粒圆度及密度因子显著增加,但籽粒千粒重变化不显著。
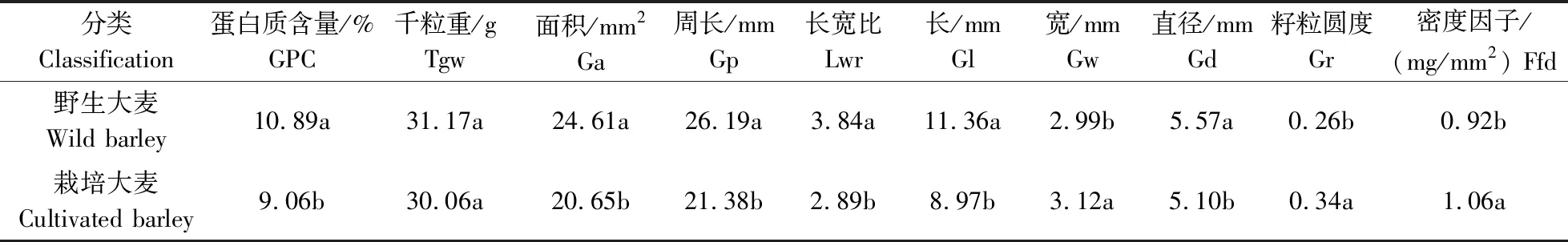
表2 栽培大麦和野生大麦籽粒性状的差异比较Table 2 Comparison of differences in grain traits between cultivated and wild barley accessions

表3 野生大麦和栽培大麦的蛋白质含量及千粒重的描述统计Table 3 Descriptive statistics of GPC and Tgw of wild barley and cultivated barley
为了更全面地了解野生大麦与栽培大麦籽粒性状的差异性,分别对野生大麦与栽培大麦在相同棱型/皮裸状态下进行了籽粒性状差异分析(表4)。结果表明:在相同棱型中,野生二棱大麦与栽培二棱大麦除了在千粒重方面没有显著差异外,在其他籽粒性状中均存在显著差异,其中野生二棱大麦GPC(11.26%)、粒长(11.51 mm)与籽粒直径(5.59 mm)均显著高于栽培二棱大麦GPC(8.51 mm)、粒长(8.92 mm)及籽粒直径(5.17 mm);而野生二棱大麦粒宽(2.95 mm)则显著低于栽培二棱大麦粒宽(3.22 mm)。而野生六棱大麦的GPC(7.30%)显著低于栽培六棱大麦(9.61%),但其他籽粒表型性状均要高于栽培六棱大麦,其中野生六棱大麦的千粒重、籽粒面积、籽粒周长、粒长、粒宽与籽粒直径均显著高于栽培六棱大麦;在同一皮裸类型中,野生皮大麦与栽培皮大麦在所有籽粒性状中的差异性与野生二棱大麦和栽培二棱大麦中的差异性相一致;在野生裸大麦与栽培裸大麦中,GPC、千粒重、籽粒周长、粒宽及籽粒密度在二者之间均存在显著差异,而其他籽粒性状差异均不显著。其中野生裸大麦GPC显著低于栽培裸大麦,而千粒重与粒宽均要显著高于栽培裸大麦。这些结果表明,同一类型(相同棱型/皮裸)野生和栽培大麦在籽粒性状中存在不同程度的差异,大麦籽粒性状在不同基因型间存在大量的变异。
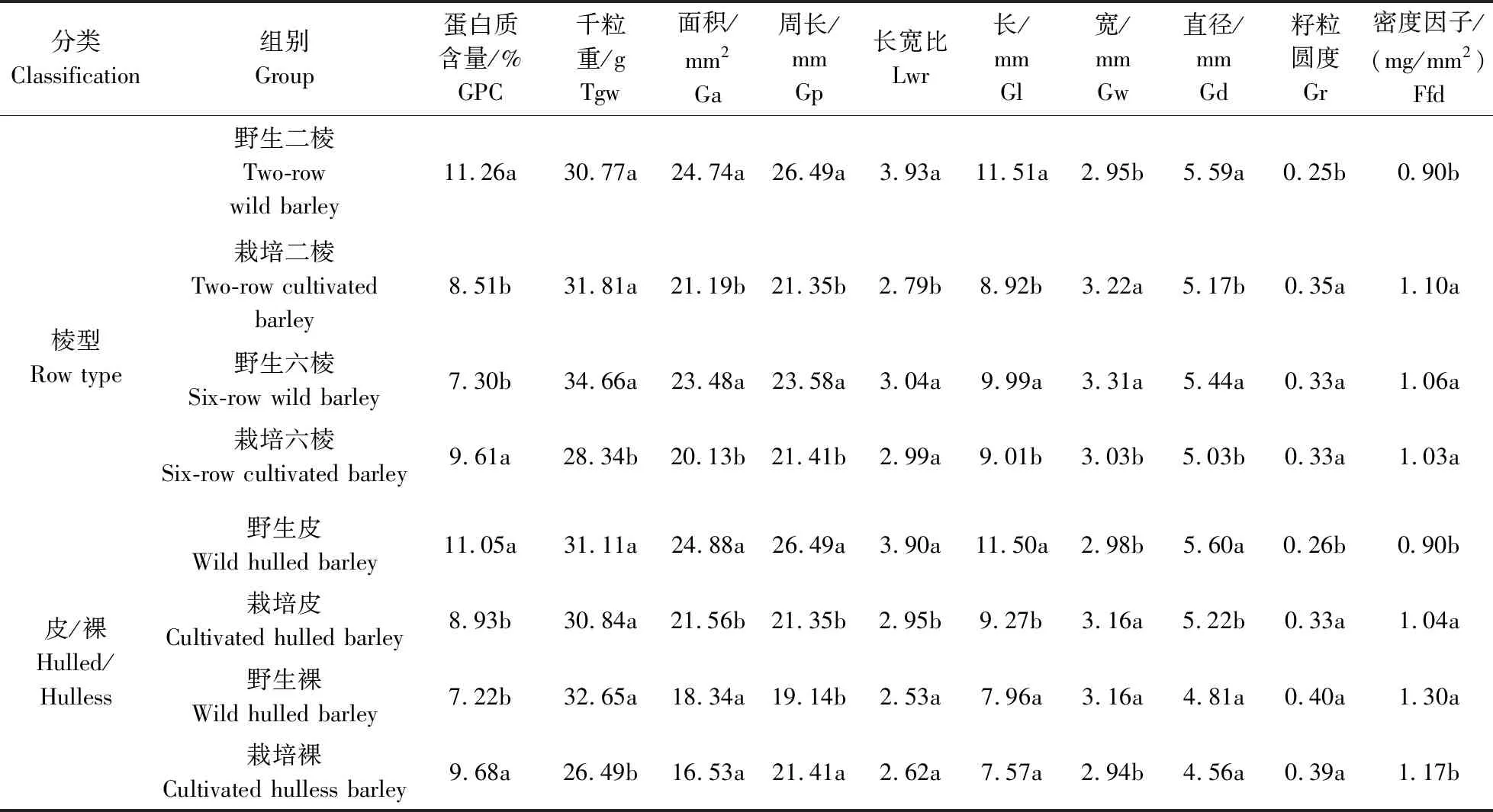
表4 不同分类栽培大麦和野生大麦籽粒性状差异比较Table 4 Comparison of differences in grain traits between cultivated and wild barley by different classification
2.3 不同分类野生大麦籽粒性状比较分析
由表5可见,在3个不同地理来源的材料中,西藏野生大麦GPC(6.85%)显著低于中亚野生大麦(11.34%)和西南亚野生大麦(12.18%)。其中西藏野生大麦GPC的最低值为5.12%,最高值仅为10.61%,还未达到中亚和西南亚野生GPC的平均水平,而西藏野生大麦GPC的变异系数却在这3个地区中是最大的,为24.03%(表6)。在千粒重方面,西藏野生大麦的千粒重(36.25 g)要显著高于中亚野生大麦(29.72 g)和西南亚野生大麦(30.18 g)(表5)。在8个籽粒表型性状中,中亚野生大麦和西南亚野生大麦均无显著差异,说明中亚野生大麦和西南亚野生大麦群体遗传背景更相近。在棱型分类中,野生二棱大麦的GPC(11.26%)要显著高于野生六棱大麦的GPC(7.30%),而野生二棱大麦千粒重(30.77 g)显著低于野生六棱大麦千粒重(34.66 g)。相对于野生六棱大麦,野生二棱大麦粒长与籽粒直径显著长于野生六棱大麦,而粒宽则显著短于野生六棱大麦。在皮裸分类中,除千粒重差异不显著外,其余所有籽粒性状在野生皮大麦与野生裸大麦中均存在显著差异。野生皮大麦GPC(11.05%)、粒长(11.50 mm)与籽粒直径(5.60 mm)均要显著高于野生裸大麦GPC(7.22%)、粒长(7.96 mm)及籽粒直径(4.81 mm),而野生裸大麦的粒宽(3.16 mm)与籽粒圆度(0.40)均显著大于野生皮大麦粒宽(2.98 mm)和籽粒圆度(0.26)。这些结果表明,在野生大麦中,GPC、粒长及粒宽与皮裸、棱型都存在一定相关性,千粒重与棱型存在一定相关性,籽粒直径与皮裸存在一定相关性。
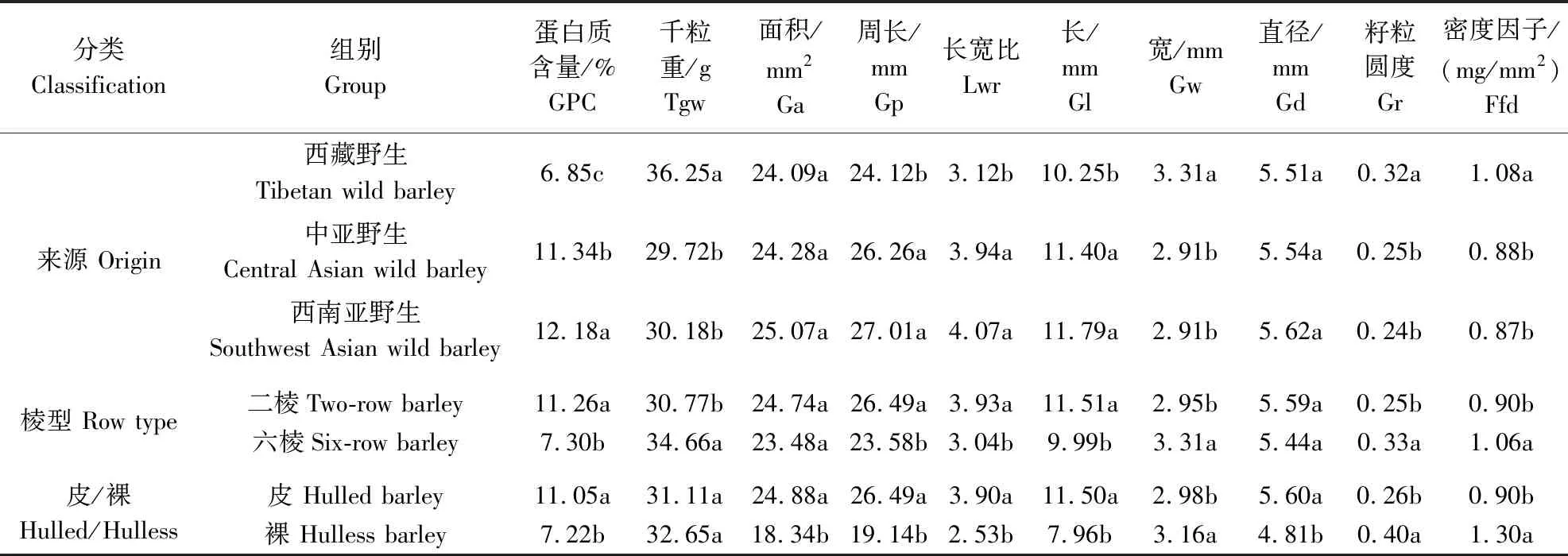
表5 野生大麦不同分类籽粒性状差异比较Table 5 Comparison of differences in grain traits of wild barley by different classification
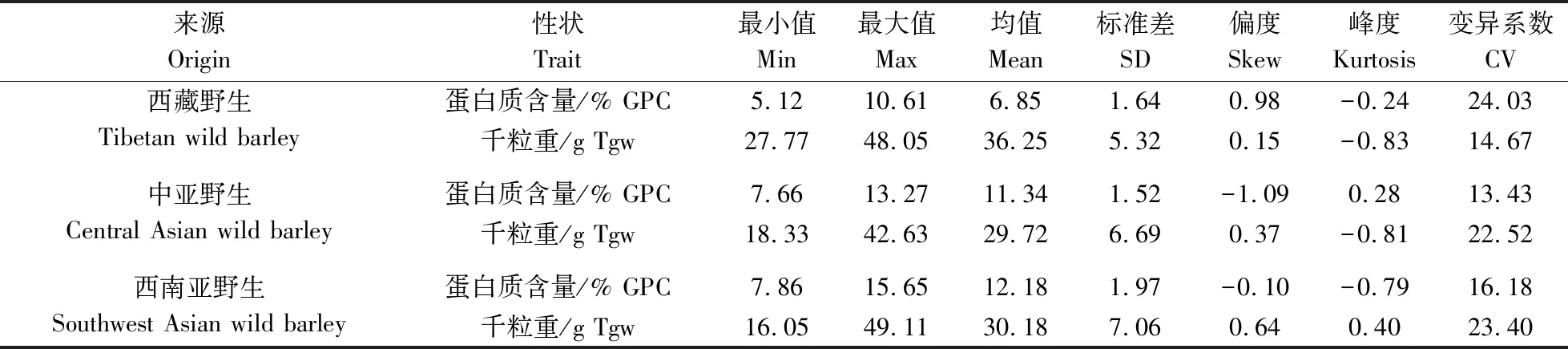
表6 不同来源野生大麦GPC和千粒重的描述性统计Table 6 Descriptive statistics of GPC and Tgw of wild barley from different origins
此外,由于西南亚和中亚野生大麦均为二棱皮大麦,而西藏野生大麦既有皮裸之分,又有不同的棱型,所以对西藏野生大麦不同棱型与皮裸大麦进行分类,结果发现西藏野生二棱大麦GPC显著低于西藏野生六棱大麦,而千粒重显著高于西藏野生六棱大麦;其他籽粒性状在西藏野生大麦不同棱型种质间均未表现出差异;GPC在西藏野生皮/裸大麦中并未表现出差异,而西藏皮大麦千粒重显著高于裸大麦(表7)。西藏野生大麦不同分类中籽粒性状差异研究表明,在西藏野生大麦中GPC在不同棱型中存在显著差异,在不同皮裸类型中差异不显著;在西藏野生大麦中千粒重在棱型与皮裸分类中都有显著差异;说明在西藏野生大麦中千粒重与棱型、皮裸都存在一定相关性,而GPC仅与棱型存在一定相关系。
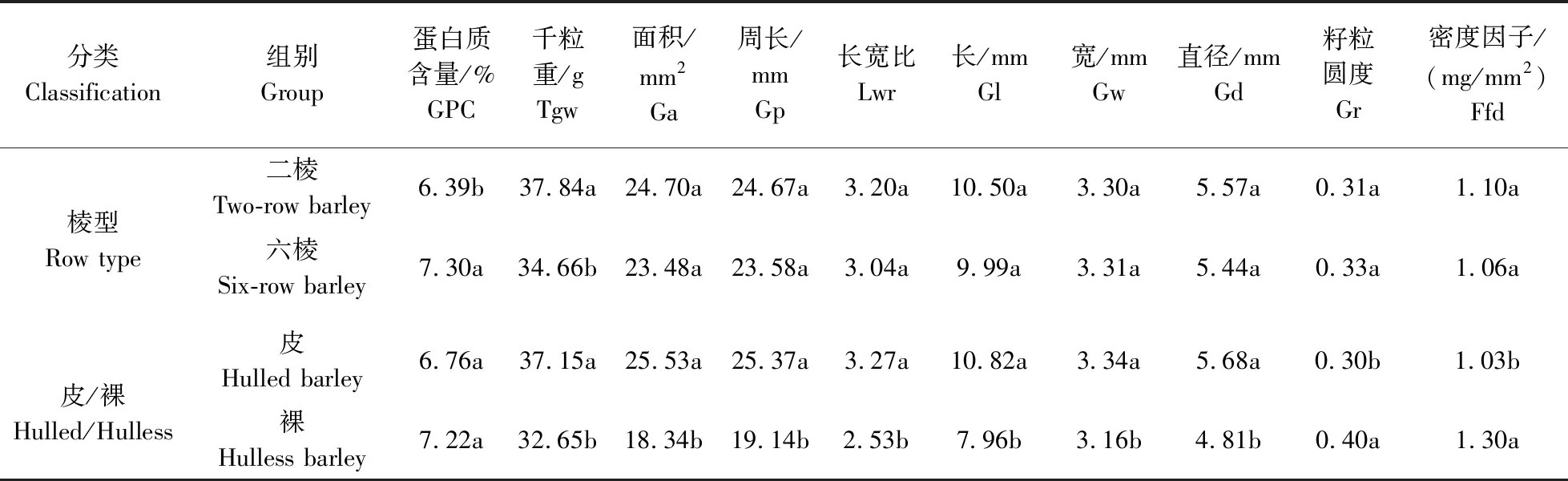
表7 西藏野生大麦不同分类的籽粒性状差异比较Table 7 Comparison of differences in grain traits of Tibetan wild barley by different classification
2.4 不同分类栽培大麦籽粒性状比较分析
由表8可知,在所有的6个地区中,东亚栽培大麦的GPC最高,为9.26%;而澳洲栽培大麦的GPC最低,为8.01%。6个不同地区栽培大麦GPC的多重比较显示两两之间在5%水平上差异均不显著,表明各个地区大麦经过长期的人工选育,GPC已经趋于一致。南美栽培大麦表现出最高的GPC变异系数,达23.73%;地中海栽培大麦的变异系数则最低,为6.99%(表9)。各个地区在千粒重方面有明显差异,最高的是北美栽培大麦,为32.47 g;最低的是澳洲栽培大麦,仅为23.69 g。千粒重变异系数东亚栽培大麦最高,为27.63%,地中海栽培大麦最低,为12.40%(表9)。在其他籽粒性状上,北美栽培大麦的粒长(9.68 mm)、粒宽(3.16 mm)、籽粒直径(5.31 mm)、籽粒面积(22.35 mm2)、周长(23.05 mm)都是6个地区中最高的(表8)。在不同棱型分类中,二棱大麦千粒重(31.81 g)显著高于六棱大麦(28.34 g),而GPC(8.51%)显著低于六棱大麦(9.61%),这与不同棱型野生大麦在该2个性状上表现的差异刚好相反。粒宽与籽粒直径在二棱和六棱栽培大麦中也表现出显著差异,而粒长差异不显著。在皮裸分类中,在栽培皮大麦与栽培裸大麦间的GPC差异不显著,而在8个籽粒表型性状上两类栽培大麦均存在显著差异。栽培皮大麦的籽粒面积、周长、粒长及籽粒直径均显著大于栽培裸大麦,这与野生大麦在皮裸两类型大麦之间的差异相一致。
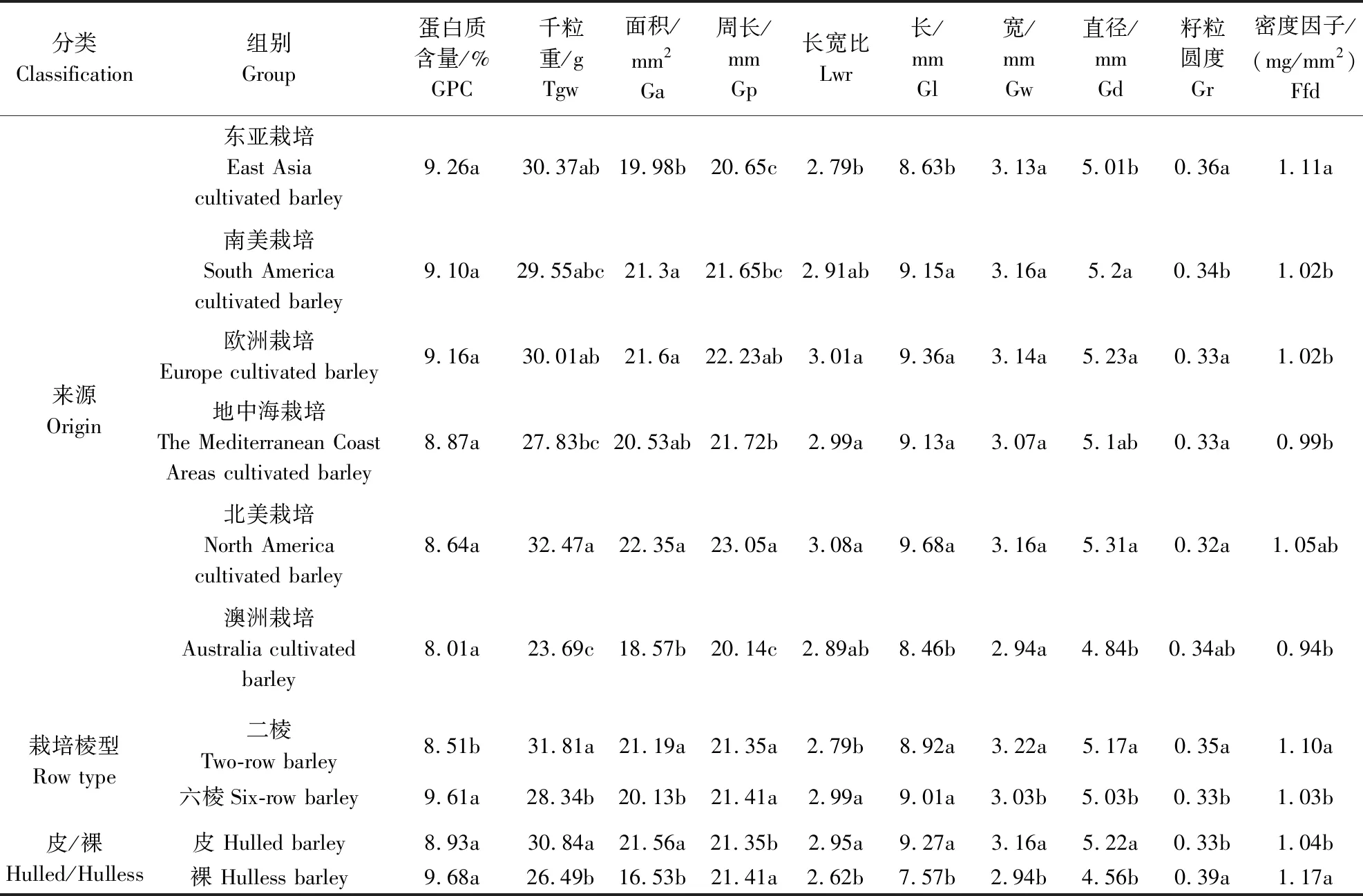
表8 栽培大麦不同分类的籽粒性状差异比较Table 8 Comparison of differences in grain traits of cultivated barley by different classification
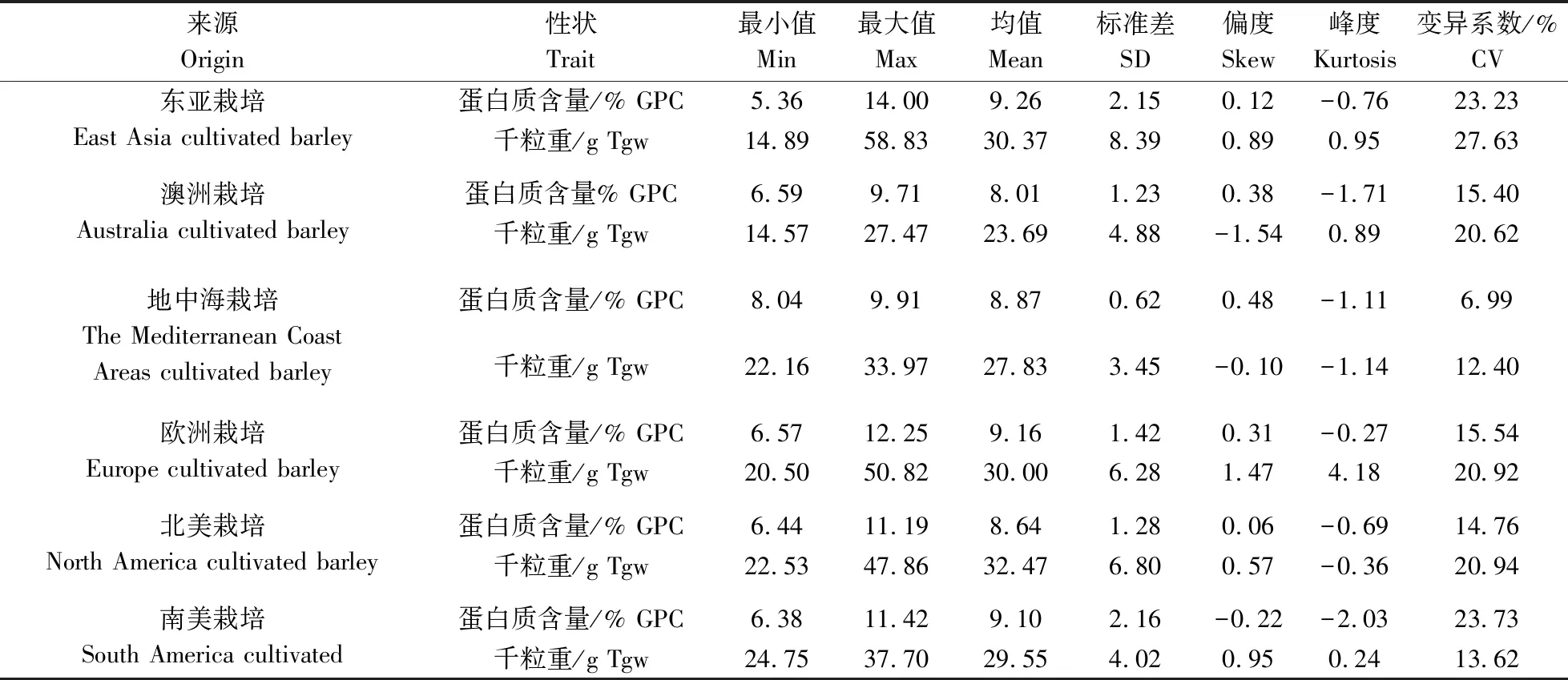
表9 不同来源栽培大麦GPC和千粒重的描述性统计Table 9 Descriptive statistics of GPC and Tgw of cultivated barley from different origins
2.5 大麦籽粒性状的相关分析
经相关性分析,发现GPC与所有表型性状均呈现显著或极显著相关(表10)。其中GPC与千粒重(-0.216**)、籽粒宽(-0.412**)、籽粒圆度(-0.440**)、籽粒密度(-0.395**)均呈极显著负相关;而与籽粒周长(0.331**)、籽粒长(0.360**) 均呈极显著正相关。从千粒重与其他表型性状相关性可知,各个籽粒形态指标与千粒重均呈显著或极显著的相关性。其中千粒重与籽粒宽(0.850**)、籽粒直径(0.681**)、籽粒面积(0.685**)、密度因子(0.610**)、籽粒周长(0.378**)、籽粒长(0.320**)均呈极显著正相关;而与长宽比(-0.121*)呈显著的负相关。在其他表型性状之间相关性中,粒长与粒宽之间没有明显的相关性,粒长与籽粒直径(0.879**),粒宽与籽粒直径(0.427**)均呈现极显著正相关。在所有呈极显著正相关的两两性状中,籽粒面积与籽粒直径、籽粒周长与粒长之间相关系数较高,分别为0.998**和0.996**,这表明籽粒直径和粒长分别对籽粒面积和周长的大小起决定性作用。籽粒长宽比与籽粒圆度极显著负相关系数最高,为-0.968**,说明籽粒长宽比是籽粒圆度的制约因子。
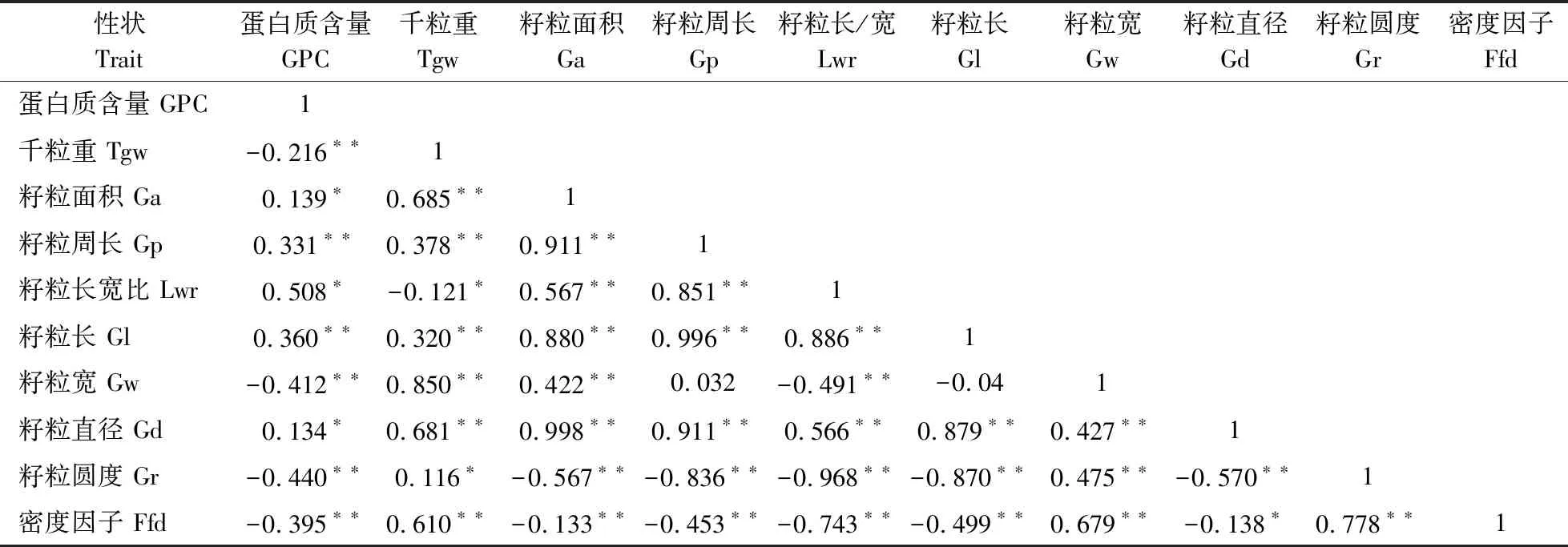
表10 大麦籽粒性状的相关系数Table 10 Correlation coefficient of grain traits in barley
2.6 不同类型大麦部分籽粒性状的相关分析
由于粒长、粒宽与籽粒直径是决定籽粒表型的关键性状,它们同时也是描述粒型最重要的性状,因此,选用这3个性状及GPC、千粒重共5个性状研究它们在不同分类方式中的相关性(表11)。结果表明,在GPC与千粒重的相关性上,除了野生六棱大麦呈极显著正相关(0.569**)外,其他组别GPC与千粒重均呈负相关;其中在野生皮大麦中,GPC与千粒重两者间呈极显著负相关性,相关系数最高,为-0.319。在不同分类方式中,GPC与粒长、粒宽、籽粒直径的相关性存在很大差异;而千粒重与粒长、粒宽及籽粒直径之间相关性基本一致,均呈现显著或极显著正相关;千粒重与粒宽相关系数都在0.85以上,最高的是野生裸大麦组别,为0.954,且千粒重与粒宽、籽粒直径的相关系数均大于千粒重与粒长的相关系数。大麦的粒长与粒宽在野生二棱、栽培、栽培二棱、栽培六棱、栽培皮大麦中均呈极显著正相关,在其他分类中相关性不显著。除野生六棱大麦外,粒长与籽粒直径、粒宽与籽粒直径在所有分类中均呈现极显著正相关。
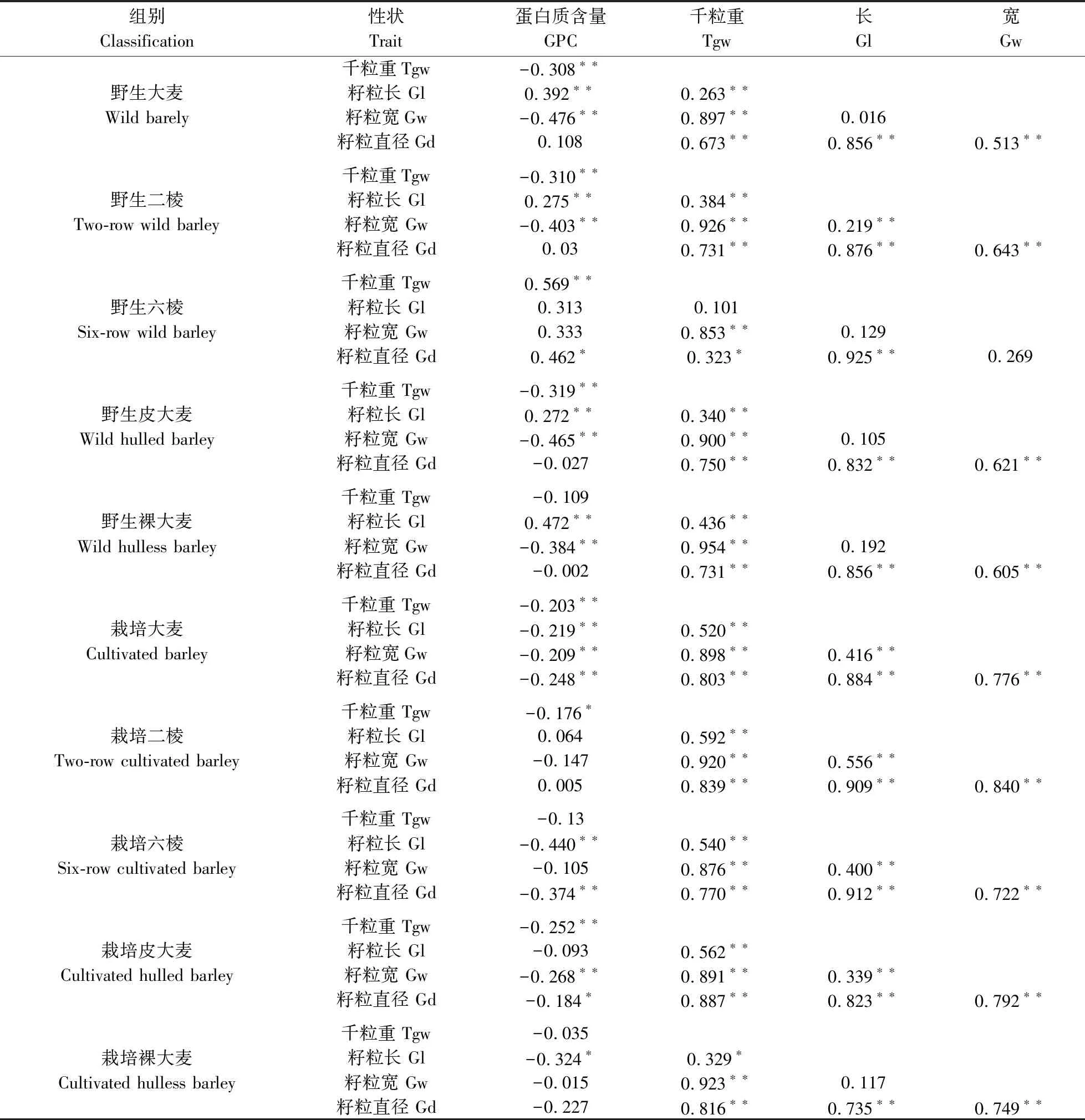
表11 不同分类大麦籽粒性状的相关系数Table 11 Correlation coefficient of barley grain traits by different classification
2.7 大麦籽粒性状的聚类分析
由于大麦的籽粒性状在2个年份均表现出较高的变异系数,且大部分性状在不同的分类层面上均表现出显著差异性,这说明214份种质在这些籽粒性状上表现出较高的多样性。为了进一步衡量大麦种质基于籽粒性状的分类情况及种质间的遗传背景关系,我们利用该研究中所测得10个籽粒性状对214份大麦材料进行层次聚类分析。结果表明,214份大麦材料首先被聚成两大类:一大类主要是野生大麦群体,另一大类主要是栽培大麦群体。该结果显示基于10个籽粒性状的野生大麦与栽培大麦2个群体间存在明显的遗传背景差异。其中第一大类又可分为Ⅰ和Ⅱ两个亚群,第二大类又可分为Ⅲ和Ⅳ两个亚群(图1)。第Ⅰ、Ⅱ、Ⅲ、Ⅳ亚群分别包括78份、23份、28份和85份大麦种质。其中第Ⅰ亚群多数以栽培皮大麦(红色圆形)组成,代表栽培皮大麦群体;第Ⅱ亚群主要以栽培裸大麦(绿色圆形)和野生裸大麦(绿色方形)组成,代表栽培裸大麦与野生裸大麦混合群体;第Ⅲ亚群代表野生皮大麦与栽培皮大麦混合群体;第Ⅳ亚群多以野生皮大麦(红色方形)组成,代表野生皮大麦群体。结果表明,以籽粒性状将214份大麦种质聚成4类时,野生裸大麦和栽培裸大麦亲缘关系更近,栽培大麦和野生裸大麦两群体与野生皮大麦群体之间发生了遗传分化现象。
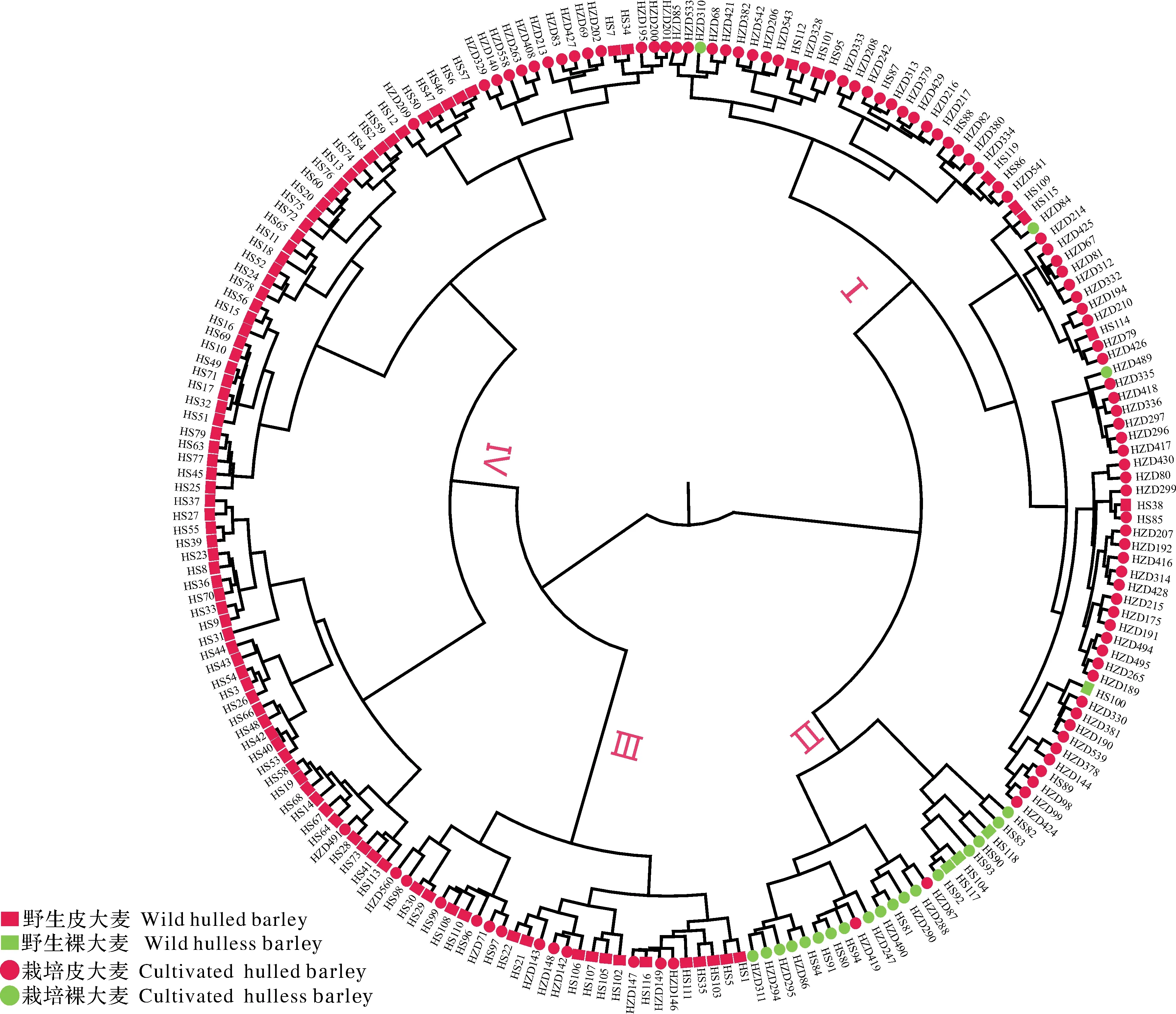
图1 214份大麦材料10种籽粒性状聚类分析Fig.1 Cluster analysis of 214 barley accessions with 10 grain traits
3 讨 论
为了解大麦群体籽粒性状遗传多样性,鉴定评价大麦种质资源,指导大麦育种的亲本选配,本研究对214份来自国内外大麦种质的籽粒性状进行了差异比较分析,研究结果表明野生大麦GPC要显著高于栽培大麦,这是由于长期以产量为育种目标而造成了栽培大麦GPC普遍降低的现象[22-23]。本研究的种质中野生大麦GPC拥有丰富的变异,变异系数高达25.22%,其中HS40(15.20%)、HS42(15.03%)、HS4(15.19%)、HS77(15.65%)4份材料具有较高水平的GPC,可以作为大麦品质改良的宝贵资源。栽培大麦相比于野生大麦,其粒长与籽粒直径显著变短,而粒宽、籽粒圆度与密度因子显著增加,这可能是与人们追求增加穗粒数的定向选择育种有关。粒大饱满均匀的籽粒不仅有利于提高产量和幼苗活力,而且可以提高麦芽产量和饲料品质,因此受到育种者的青睐[24]。不同棱形、皮裸特性的大麦,其作用有着很大的区别[25-26],我们分别对野生大麦和栽培大麦在同一棱形/皮裸特性下进行籽粒性状的差异比较分析,有针对性地对不同类型大麦新品种的选育提供参考依据。此外,棱型与皮裸性状是大麦驯化过程中的关键性状[24],通过同一类型中野生大麦与栽培大麦籽粒性状的比较,可以明确大麦进化过程中籽粒性状的变化历程。不同来源野生大麦的差异分析中,中亚野生大麦与西南亚野生大麦的各个表型性状均无显著差异,表明两地野生大麦遗传背景相近,闫松显[27]用分子标记的聚类结果也证明了西南亚野生大麦种质和中亚野生大麦种质具有较近的亲缘关系,而西藏野生大麦与上述2个地理来源的野生大麦之间在籽粒面积和籽粒直径以外的所有籽粒性状上均有显著差异,间接证明了西藏野生大麦具有独特的遗传背景,Dai等[28-29]研究结果也表明西藏野生大麦与西南亚野生大麦基因组间存在很大差异,西藏野生大麦对栽培大麦基因组有特殊的贡献,进一步支持了西藏是大麦一个起源中心的理论。在3个地区的野生大麦中西藏野生大麦拥有最高的千粒重和最低的GPC,因此,可以作为啤酒大麦新品种培育的重要资源。如西藏野生大麦种质中HS107 GPC最低,为5.12%,该结果低于马得泉等[30]研究的2 004份西藏野生大麦材料的最低值(8.42%)。其次,HS102(5.36%)、HS105(5.26%)及HS109(5.28%)等材料也均具有较低水平的GPC,这些低GPC大麦种质可以作为低蛋白大麦品种遗传改良的珍稀材料。在不同栽培大麦地区中,6个地区之间GPC差异不显著,说明长期的遗传改良不仅使得栽培大麦GPC逐渐降低,并且在世界各地区栽培大麦GPC已经趋于相似水平。因此,亟需加强对不同GPC大麦品种的研发,以满足畜牧业和啤酒加工业的需求。东亚栽培大麦千粒重和GPC均有较高的变异系数,这表明东亚栽培大麦在千粒重和GPC方面变异类型比较丰富,可作为千粒重和GPC改良的供体材料。北美栽培大麦在6个地区中各个籽粒性状均有较高表现,可引进该地区HZD140和HZD333等种质作为优化籽粒性状的种质资源。在不同棱型与皮裸分类中,就野生大麦GPC而言,二棱野生大麦高于六棱野生大麦,野生皮大麦高于野生裸大麦,这与马得泉等[30]的报道一致。目前对于野生大麦不同分类的籽粒表型性状的研究鲜有报道,本研究结果对大麦不同类型之间籽粒性状的差异分析进行了补充。陈晓东等[7]对125份栽培大麦材料籽粒性状研究发现栽培大麦中二棱大麦的粒重、粒宽和粒长都要显著高于六棱大麦,其中粒重与粒宽的差异与本研究的一致,而粒长差异不同。这可能是因为所选材料的来源差异引起的,本研究选用的是世界各地的栽培大麦材料,而陈晓东等[7]选用的栽培大麦材料仅来自于国内。对于栽培二棱大麦与六棱大麦之间的差异分析显示,二棱大麦比六棱大麦GPC更低而千粒重更高,因此更适合啤酒加工业的需要,而六棱大麦宜选作饲料大麦。无论是栽培大麦还是野生大麦,皮大麦的粒长与籽粒直径显著高于裸大麦,这应该是受到麸壳的影响[31]。皮裸大麦的用途不同,他们之间的遗传差异是长期驯化所致,层次聚类的结果也表明皮大麦与裸大麦在籽粒性状方面有明显差异。大麦籽粒的很多性状在不同类别间存在不同程度的差异,表明大麦籽粒性状在不同基因型间存在大量变异,人工选择对大麦籽粒性状改良有重要的作用。
谷类作物不同籽粒性状之间的相关性已有较多报道,本研究发现GPC与千粒重呈极显著负相关,与前人研究结果相一致[32]。然而,本研究发现在野生六棱大麦籽粒性状间的相关分析中,GPC与千粒重呈极显著正相关而且相关系数较高,后续可针对这一特殊现象在分子水平上或基因组水平上进行深度探析,该材料在指导大麦品质与产量同步优化方面具有重要意义。张亚东等[33]发现大粒型水稻粒重的增加主要取决于籽粒长度的增加,粒宽对粒重的影响较小,这与本研究的结果不同。张新忠等[8]发现千粒重与粒宽呈极显著正相关,且相关系数高于粒长与千粒重的相关系数,与本研究结果一致。另外本研究也对籽粒直径与千粒重相关性进行了研究,发现二者之间也存在极显著的正相关,并且相关系数高于粒长与千粒重的相关系数。在不同的分类方式下,千粒重与粒长、粒宽、籽粒直径之间的相关性均有一致的结果。因此,增加粒宽与籽粒直径可能是提高千粒重的有效途径。此外,本研究也发现籽粒的长宽比与千粒重呈显著的负相关,这与青海地区小麦粒型分析结果相一致[34]。许如根等[6]通过对籽粒性状的研究发现,籽粒的长宽比太大会影响到大麦的饲用和食用品质。人类在育种过程中的定向改良使得大麦品种的多样性受到选择和漂移等因素的侵蚀[35],从而使栽培大麦亲本遗传基础逐渐缩小,导致新育成品种在抗病、抗逆及品质性状等方面难以突破。而野生大麦的种质资源群体结构特殊,遗传多样性水平较高,含有丰富的抗(耐)性等优良基因,可作为大麦遗传改良的重要的基因资源。本研究所选用的大麦材料来自国内外各个地区,代表性较强,且选用的野生大麦在籽粒性状上存在丰富的变异,后续可开展野生大麦优异基因的挖掘等研究工作,以期为大麦亲本选配及新品种籽粒表型性状的选育和蛋白质品质遗传改良提供参考依据。
