吊罗山薹草开花生物学特征及繁育系统研究
2021-05-19林鹏杨虎彪
林鹏 杨虎彪

摘 要:开花生物学特征和繁育系统在生殖生长过程中有着重要的作用。对吊罗山薹草(Carex diaoluoshanica)的开花生物学特征和繁育特性开展定点观测研究,发现吊罗山薹草的小穗呈两性,但顶生小穗的雌花完全退化,而基部侧生小穗的雄花虽高度退化但仍有1~2朵匿藏的雄花发挥自交作用;开花物候和开花动态观测中发现,吊罗山薹草开花同步指数为0.16,顶生雄小穗的花期约5 d,侧生两性小穗的花期约6 d,而群体花期为26 d;在散粉后1 h时花粉活力达到最大,为85%,随后几小时内快速下降;柱头在散粉前1 d已经达到成熟状态,在散粉时依然保持着极高的可授性,到散粉第5天时,柱头不再具有可授性;根据异交指数和花粉/胚珠比检测,吊罗山薹草的繁育系统为兼性自交,进一步的套袋试验验证发现,其繁育系统是以自交为主,兼有异交的混合型交配系统。
关键词:吊罗山薹草;开花物候;花粉活力;柱头可授性;繁育系统
中图分类号:S54 文献标识码:A
Flowering Biological Characteristics and Breeding System of Carex diaoluoshanica
LIN Peng1,2, YANG Hubiao1*
1. Tropical Crops Genetic Resources Institute, Chinese Academg of Tropical Agricultural Sciences, Haikou, Hainan 571101, China; 2. College of Agro-grassland Sciences, Nanjing Agriculture university, Nanjing, Jiangsu 210095, China
Abstract: The characteristics of flowering biology and breeding system play an important role in plant growth and reproduction. In this paper, the biological characteristics and breeding characteristics of Carex diaoluoshanica were studied based on the population of Diaoluoshan Natural Reserve. It was found that the spikelets were bisexual, but the female flowers of the terminal spikelets were completely degenerated, while the male flowers of the basal lateral spikelets were highly degenerated, but 1~2 male flowers were still remained. According to the observation of flowering phenology and flowering dynamics, the flowering synchronization index of C. diaoluoshanica was 0.16, and the flowering period of male spikelet was about 5 d, while that of lateral pistillate spikes was about 6d, and the flowering period of population was 26 d. The pollen viability reached a maximum of 85% at 1 h after powder dispersal, and then decreased rapidly within a few hours. The stigma had reached the mature state one day before powder dispersal, and remained highly activity in pollinating period, but no longer with activity after 5 days of pollination. According to the OCI and P/O index, the breeding system type of C. diaoluoshanica was facultative autogamy, further bagging experiments showed that the breeding system was mixes with self-pollination and out-crossing, and tended to selfing.
Keywords: Carex diaoluoshanica; flowering phenology; breeding system; pollen viability; stigma receptivity
DOI: 10.3969/j.issn.1000-2561.2021.03.010
生殖過程在植物的生活史中扮演着重要角色,任何环节受到阻碍都有可能导致种群的衰退。繁育系统是植物为适应环境条件,形成影响后代遗传组成的有性生殖特征总和,其组成内容包括花部形态特征总和、花各部位活力和寿命、传粉授粉行为和交配习性,其本质上是内部遗传机制和外部环境影响相互作用的综合表现,并以此影响种群的动态,在植物进化路线和表征变异上起到决定性的作用[1-3],因此,对植物的花部特征与繁育系统特性的深入研究对濒危植物的保护具有重要的指导作用,且越来越受到研究人员的重视。近年来,丰都车前(Plantago fengdouensi)、红榄李(Lumnitzera littorea)、金花猕猴桃(Actinidia chrysantha)、长梗杜鹃(Rhododendron longipedicellatum)、泰山花楸(Sorbus taishanensis)等众多珍稀濒危物种相继被开展繁育生物学研究,以探索其濒危机制[4-8]。吊罗山薹草(Carex diaoluoshanica H. B. Yang, G. D. Liu & Q. L.Wang)是莎草科(Cyperaceae)薹草属(Carex)植物,该物种是本研究团队在海南吊罗山发现并公开发表的新物种[9],模式标本(Holotype)保存于中国科学院华南植物园标本馆IBSC,同形标本(Isotype)保存于中国热带农业科学院热带作物品种资源研究所标本馆。吊罗山薹草喜生于高郁闭度的热带雨林中,是热带雨林低层原生草本植物的重要代表,本研究团队对吊罗山薹草的种群分布展开了深入调查,目前只在吊罗山发现2个居群,其资源现状总体表现为种群数量极低、单个种群占有面积极小、种群呈严重破碎化分布。根据《中国生物多样性红色名录——高等植物卷》的评估方法[10],吊罗山薹草至少满足濒危(Endangered)的评定标准。根据分布区域十分狭窄且严重破碎化的现状,推测其资源现状将进一步恶化。因此,本文通过样地定点观测对吊罗山薹草的花部特征、开花物候、繁育系统等进行研究,为该物种的繁育保护提供重要支撑。
1 材料与方法
1.1 观测样地概况
吊罗山薹草(Carex diaoluoshanica)只在吊罗山自然保护区发现2个小居群,居群间的距离为4 km,居群海拔分别为948 m和974 m。观测样地所在区域的年降雨量为2300 mm,相对湿度月平均为85%,雨季达到90%以上,年均温度为24 ℃,土壤以山地黄壤为主。
1.2 方法
1.2.1 开花物候和花部特征测量 每居群内随机选20株吊罗山薹草进行标记,每株标记1个花序。花朵开放后,每天上午8:00观察一次,记录该花序上新开小穗的数量,直至开花结束。花部特征的观测注意记录花朵开放、花粉散出、柱头伸长、小花形状及颜色等信息等,并测量雌、雄小穗、花药、柱头等长度的变化。开花动态的观察,标记生长状态相近20个花序,从抽穗开始每天观察2次,观察开花的时间、花的状态和开放过程,雌雄蕊的变化等,并描述拍照。种群开花进程的记录按照Dafni所述的标准开展[11]。
1.2.2 花粉活力与柱头可授性测定 使用TTC染色法测定吊罗山薹草的花粉活力。标记花期一致的5个花序,观测雄花开放动态,由花药完全抽出开始标记检测,散粉后每1 h检测一次,直至散粉4 h之后结束。具体操作是将花粉置于载玻片上,滴加1~2滴0.5%TTC溶液,放入培养皿中并在25 ℃黑暗条件下培养2 h左右。在显微镜下观察,花粉变红则表明有活力,无变化或变黑色则表明无活性。统计全部花粉中红色花粉所占比例。以染色率表示花粉的活力百分率,即有活力花粉的比率=红色花粉数/观察花粉总数× 100%[12]。
用联苯胺—过氧化氢法测定柱头可授性[12]。标记花期一致的5个花序,从侧生小穗雌花开放前2天开始每天检测一次,直至雌花完全开放后第4天柱头颜色由白色透明转棕色时结束。具体操作是将柱头从小花上剪下置于载玻片上,滴加少量反应液(1%联苯胺∶3%过氧化氢∶水=4∶11∶22),黑暗条件放置15 min。若柱头具有可授性,柱头附近的液体会呈蓝色并产生气泡,气泡的多少反映出柱头可授性的强弱。
1.2.3 异交指数的计算 按照Dafni的方法测量相关指标参数来判断繁育系统的类型。Dafni标准中,异交指数值是花朵直径赋值(<1 mm记为0;1~2 mm记为1;2~6 mm记为2)、雌雄小花开放时间间隔赋值(同时或雌蕊先熟记为0;雄蕊先熟记为1)、柱头与花药空间位置赋值(同一高度记为0;位置分离记为1)的三者之和。当异交指数为0时,交配方式为闭花受精;当异交指数为1时,为专性自交;当异交指数为2时,为兼性自交;当异交指数为3时,为自交亲和有时需要传粉者;当异交指数为4时,为部分自交亲和,异交需要它媒[11]。
1.2.4 花粉/胚珠比检测 选取10个侧生两性小穗,取小穗的全部花药,待其散粉后,加入纤维素酶溶液2 mL,于常温放置24 h将游离花粉粒制成悬浮液,吸取定量悬浮液5次重复镜检统计,根据公式计算花粉量。吊罗山薹草的花粉/胚珠比(P/O)的测算如下,P/O=小穗平均单花花粉量/小穗胚珠数。依据Cruden的标准(P/O值区间在2.7~5.4时为闭花授精;P/O值区间在18.1~39.0时为专性自交;P/O值区间在31.9~396.0间为兼性自交;P/O值区间在244.7~2588.0时为兼性异交;P/O值区间在2108.0~195525.0时为专性异交)判断交配方式[13]。
1.2.5 套袋试验 套袋试验的处理如下:①不去雄,直接套袋检测自交结实率;②雌、雄花开放前去雄,并作套袋处理检测无融合生殖现象;③雌、雄花开放前去雄,并作套袋处理,待雌花开放后辅助授异株花粉检测异花授粉结实率;④对照处理为标记自然授粉植株统计其结实率。每个处理5个重复。
1.3 数据处理
用SPSS 19.0和Excel 2010軟件进行数据处理,结果以平均值±标准差表示。
2 结果与分析
2.1 开花物候观察
从个体和群体2个水平随机对20株吊罗山薹草进行开花物候的观察,结果如表1。从表1可以看出,吊罗山薹草的个体于5月9日进入初花期,因其小穗上小花致密,开花时小花同日开放,其个体水平的盛花期与始花期重叠,因此,其盛花期也为5月9日,于5月15日进入个体的终花期,花期结束,个体水平的花期约为6 d。吊罗山薹草在5月20日开放个体数超过标记植株的25%,进入始花期,在5月30日标记植株中半数已开放,进入盛花期,6月15日开花个体数不足5%,吊罗山薹草的群体水平进入终花期,其群体花期持续时间约为26 d。吊罗山薹草的开花同步指数为0.16,开花同步指数代表各个体开花时间的交叉程度,数值越大表示各个体开花交叉时间越长。
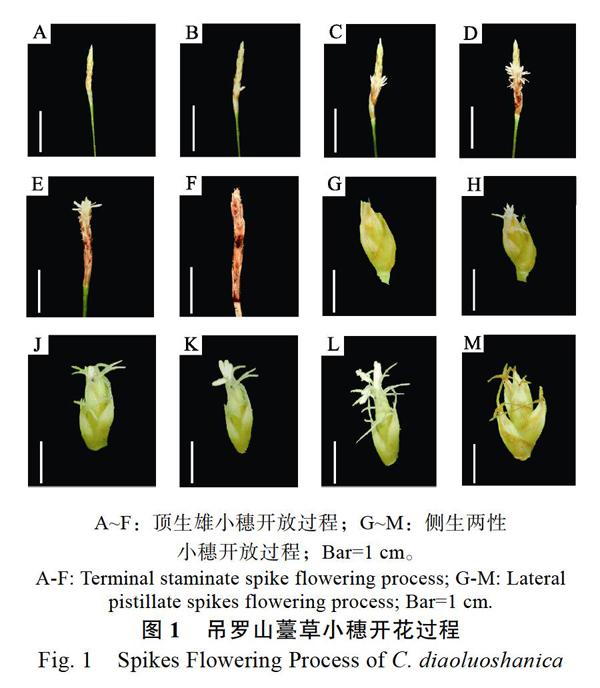
开放动态上,吊罗山薹草顶生小穗的花期持续时间约为5 d;基部侧生两性小穗花期约为6 d。开花前雄小穗上各个小花之间紧密排列成穗状,即将开花时各个小花之间出现明显分界,开花时底部小花先开放,花药伸出。随后,小花自下而上呈轮状开放,直到顶部小花开放,顶部小花开放后,整个小穗逐渐枯萎,5 d之后开花结束。花药多在上午伸出,于中午左右散粉,开裂方式为纵裂,散粉过程持续2~3 h(顶生雄小穗的开花动态如图1,A~F)。两性小穗雌花开放时,柱头伸出伸长,2~3 d后花药从雄小花苞片中伸出,散粉,散粉多于中午开始,持续2~3 h。散粉后1~2 d雌花柱头的颜色逐渐转为棕色并枯萎,子房膨大,开花结束(侧生两性小穗的开花动态如图1,G~M)。此外,观测过程中发现,雄花的散粉与天气状况有关,散粉集中于睛天气温回升的中午前后,当遇持续降雨时不散粉,超过2 d以上的持续降雨时成熟的花药发生腐败。
2.2 花部综合特征观测
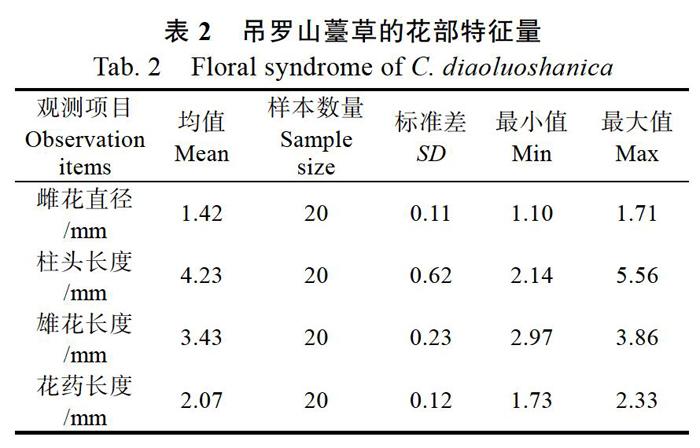
前期观测认为,吊罗山薹草具有1枚独立的顶生雄小穗,而基部侧生小穗是雌性,因而被认为吊罗山薹草的其小穗是单性。然而,本研究套袋实验中发现,去雄(顶生雄小穗)之后基部侧生雌小穗仍然正常发育,这一现象引起了对它花部特征的再度关注,深入观察后发现其基部侧生小穗总体上是雌花,但位于小穗顶部位置仍匿藏有1~2朵雄花,而此雄花与雌花同形,在正常条件下被雌花所包围而不易发现,只在雌花完全开放之后此枚雄花才开放,具有一定的隐匿性。因此,吊罗山薹草的小穗应该是两性,但顶生小穗的雌花极度退化只剩雄花,而基部侧生小穗的雄花高度退化,但仍有1~2朵雄花正常发育。其完整的花部特征如下,花序顶部有1枚独立雄小穗(雌花完全退化),多朵雄花呈纵向致密排列;基部侧生者是两性小穗,其顶部匿藏有1~2朵雄花并被雌花包围,具3枚花药,花药圆柱形、白色、长约(2.07±0.12)mm;雌花4~6朵,少数达10朵,每朵3枚柱头,柱头丝状、白色、表面分泌粘液,长约(4.23±0.62)mm。吊罗山薹草的花部特征量如表2。
2.3 花粉活力和柱头可授性分析
吊罗山薹草的花粉在花药开裂散粉前没有活力,在散粉后1 h时活力达到最大为85%,随后几小时内快速下降,4 h后花粉活力仅为27%(图2)。
通过联苯胺?过氧化氢法对吊罗山薹草的柱头可授性进行测定,结果如表3,从表3可以看出,吊罗山薹草的柱头在散粉前1 d已经达到了成熟状态,具有最强的可授性,在散粉时依然保持着极高的可授性,当到散粉第2天时,其柱头颜色加深,柱头可授性呈现下降趋势,第3天其柱头颜色进一步加深为棕色,可授性持续下降,而到散粉第4天时,只有部分柱头维持着微弱的可授性,到散粉第5天时,柱头已经完全枯萎,变为深褐色,不再具有可授性。
2.4 异交指数测定
根据Dafni[11]的方法对吊罗山薹草的异交指数进行观测,随机测量20株植株的小穗。经计算,吊罗山薹草的平均小穗直径为(2.78±0.28)mm,赋值为2;雌花先熟,赋值为0;两性小穗雌花和雄花的空间位置相近,赋值为0。因此,其异交指数(OCI)为2,参考Dafni[11]的交配方式赋值总和,吊罗山薹草的繁育系统类型為兼性自交。
2.5 花粉/胚珠比检测
依据Cruden[13]的标准,随机对10个侧生两性小穗的未开裂的雄小花进行花粉量测定。其侧生两性小穗全部雄花通常只有1~2朵(平均单花花粉量为1003.33±77.33),对应的雌花有4~6朵。根据测算方法,吊罗山薹草的P/O值介于154~214间。因此,理论上吊罗山薹草的交配方式是兼性自交。
2.6 套袋试验
吊罗山薹草在去雄并套硫酸纸袋处理后,不结实,表明其繁育方式不包括无融合生殖;在直接套袋处理即自花授粉的状态下,结实率为91.00%±5.03%,这与对照处理的结实率间无显著差异;而在雌、雄花开放前去雄并套袋,待雌花开放后辅助授异株花粉处理下,结实率仅为43.70%±4.83%,显著低于自然状态下的结实率(P<0.05)(图3)。因此,根据异交指数、花粉/胚珠比检测和套袋试验的验证发现,吊罗山薹草的繁育系统是以自交为主,兼有异交的混合型交配系统。
3 讨论
开花物候是植物在不同的外界环境条件下,长期生长而形成的响应规律,可以用各水平花期、开花同步指数等参数来表示种群对环境的适应能力[1-2, 14-16]。吊罗山薹草的单花花期约为6 d,群体花期26 d,开花同步指数仅有0.16,开花极不同步,似乎没有机会进行异花授粉,类似的开花不同步性在珍稀濒危植物距瓣尾囊草(Urophysa rochkii)、矮沙冬青(Ammopiptanthus nanus)、灰叶胡杨(Populus pruinosa)等中也有表现[17-19],然而吊罗山薹草的开花物候依然区别于上述濒危植物,虽花期都极不同步,但前者的群体花期都长,而吊罗山薹草的群体花期只有1个月左右。关于前者,研究认为花期不同步、群体花期长的优势在于降低了开花期间外界因素对生殖过程的影响,是对干旱、严寒等极端气候条件的适应;大量的研究认为,开花物候与其生境间确实存有密切的联系[15, 20-21],吊罗山薹草的花期正值雨季,观测发现连续降雨时吊罗山薹草不散粉,而散粉过程中遭遇强降雨会导致花粉被冲刷脱落,同时发现超过2 d以上的持续降雨导致成熟的花药发生水溶性腐败,从而降低授粉成功的机率,而本观测居群所在区域的雨季长达4个月,因此形成群体花期缩短、花药晴天中午散粉和散粉后快速失活等特性是吊罗山薹草应对热带雨林特殊气候条件的适应性调整。
植物的交配方式中很少是单纯的自交或单纯的异交[22]。薹草属植物小穗有单性和两性的区别,两性小穗中,又可分为顶雄基雌和顶雌基雄[23-24],吊罗山薹草的小穗严格意义上也是雄雌顺序的两性,但又区别于以上常见模式,其顶生两性小穗的雌花完成退化而只剩雄花;而侧生两性小穗的雄花高度退化但依然于雌花间包藏有1~2朵雄花,侧生小穗开花时雌花先熟,雄花花药伸出后
几乎与柱头位于同一高度,这种小穗结构导致雄小花的花药散粉后,花粉几乎全部洒落在雌花柱头上,这种特征极大地增加了其自交的几率;而其顶生小穗(雄性,其雌花退化)的开放时间都早于基部的侧生两性小穗,形成了本株雌、雄花期不遇的假象(顶生雄小穗小花开放早于侧生两性小穗,形成其雄花与雌花不相遇的状态。但是,侧生两性小穗的雄、雌小花花期相遇),这种特殊的结构和开花行为决定了吊罗山薹草自交优先,但顶部雄小穗先于侧生两性小穗开放的习性也为异花授粉提供了机会。植物的繁育受到花粉、生境等多种因素的制约,相关研究表明,花粉限制是影响植物繁育的主要因素[25]。花粉活性和持续时间因物种和生境的不同而有所差异,如海枣花粉在?20 ℃条件下保存一年后仍然可以保持活力[26];禾本科的花粉活力比较低,玉米花粉在0~5 ℃条件下活力仅可持续10 h,超过24 h便失去活力[27];吊罗山薹草的花粉活力持续时间很短,散粉4 h后活力仅有27%。除了花粉活力之外,花朵成熟的另一个标志是柱头可授性,柱头可授性对植物的传粉方式有着重要的影响。柱头可授期随着物种的不同也存在些许差异,濒危植物肉苁蓉在花朵开放后柱头才有可授性,活性可持续2 d[28];珍稀濒危植物掌叶木(Handeliodendron bodinieri)的柱头在展开前已具备可授性,活性可持续3 d[29];吊罗山薹草的柱头在散粉前一天和散粉当天可授性最强,随着逐渐减弱,到第5天完全失去可授性。综上所述,花粉活力时间短、柱头可授性时间短和群体花期短等特点是吊罗山薹草主动应对花期外界条件的重要策略;而以环境适应性形成稳定的花部形态构造和开花习性差异推测其交配系统更倾向于自交,进一步的套袋试验也验证了吊罗山薹草异交亲和,但异交结实显著低于自交,形成了自然状态下居群长期自交的现状,长期自交导致后代适合度降低也是制约种群发展的重要因素,下一步的研究将聚集于上述推测进一步开展。
参考文献
[1] Schoen D J, Johnson M T J, Wright S I. The ecology, evolution, and genetics of plant reproductive systems[J]. The New Phytologist, 2019, 224(3): 999-1004.
[2] Lawrence D Harder, Spencer C H Barrett. Ecology and evolution of flowers[M]. Oxford University Press, 2016.
[3] 陈进燎, 周育真, 吴沙沙, 等. 台湾独蒜兰传粉机制和繁育系统研究[J]. 森林与环境学报, 2019, 39(5): 460-466.
[4] 万海霞, 邓洪平, 何 平, 等. 濒危植物丰都车前的繁育系统与传粉生物学研究[J]. 生态学报, 2018, 38(11): 4018-4026.
[5] 李太强. 极小种群野生植物长梗杜鹃的保护生物学研究[D]. 北京: 中国林业科学研究院, 2018.
[6] 宋雪艳. 珍稀濒危植物泰山花楸的濒危机制和保护研究[D]. 泰安: 山东农业大学, 2017.
[7] 张 颖, 李燕华, 张晓楠, 等. 濒危红树植物红榄李开花生物学特征及繁育系统[J]. 应用与环境生物学报, 2017, 23(1): 77-81.
[8] 莫权辉, 李洁维, 龚弘娟, 等. 濒危植物金花猕猴桃繁殖生物学初步研究[J]. 广西植物, 2016, 36(6): 640-645.
[9] Hubiao Y, Qinglong W, Changjun B, et al. Carex diaoluos-hanica (Carex sect. Lageniformes, Cyperaceae), a new species from Hainan, China[J]. PLoS One, 2014, 9(6): e97658.
[10] 环境保护部和中国科学院. 中国生物多样性红色名录高等植物卷评估报告[J]. 2013: 8-18.
[11] Dafni A. Pollination ecology: a practical approach[M]. Oxford: Oxford University Press, 1992: 1-57.
[12] 胡适宜. 植物胚胎学实验方法(一)花粉生活力的测定[J]. 植物学通报, 1993, 10(2): 60-62.
[13] Cruden R W. Pollen-ovule ratios: a conservative indicator of breeding systems in flowering plants[J]. Evolution, 1977, 31(1): 32-46.
[14] Mccabe G J, Wolock D M. Trends and Temperature Sensitivity of Moisture Conditions in the Conterminous United States[J]. Climate Research, 2002, 20(1): 19-29.
[15] 李曉婷, 陈 骥, 郭 伟. 不同气候类型下植物物候的影响因素综述[J]. 地球环境学报,2018,9(1): 16-27.
[16] 李荣平, 张晓月, 李 晶. 气候变化背景下紫丁香花期物候特征及其模拟研究[J]. 北方园艺, 2018, 40(1): 109-113.
[17] 焦培培, 李志军. 濒危植物矮沙冬青开花物候研究[J]. 西北植物学报, 2007, 27(8): 1683-1689.
[18] 周正立, 李志军, 龚卫江, 等. 胡杨、灰叶胡杨开花生物学特性研究[J]. 武汉植物学研究, 2005, 23(2): 163-168.
[19] 别鹏飞, 唐 婷, 胡进耀. 珍稀濒危植物距瓣尾囊草(Urophysa rochkii)的开花物候和繁育系统特性[J]. 生态学报, 2018, 38(11): 3899-3908.
[20] Silva P H M D, Sebbenn A M, Grattapaglia D. Pollen-mediated gene flow across fragmented clonal stands of hybrid eucalypts in an exotic environment[J]. Forest Ecology and Management, 2015(356): 293-298.
[21] Lessard-Therrien M, Davies T J, Bolmgren K. A phylogenetic comparative study of flowering phenology along an elevational gradient in the Canadian subarctic[J]. International Journal of Biometeorology, 2014, 58(4): 455-462.
[22] 王崇云, 党承林. 植物的交配系统及其进化机制与种群适应[J]. 武汉植物学研究, 1999, 17(2): 163-172.
[23] 中国科学院中国植物志编辑委员会. 中国植物志第12卷[M]. 北京: 科学出版社, 1990: 1-599.
[24] Standley L A. Anatomical aspects of the taxonomy of sedges (carex, cyperaceae)[J]. Canadian Journal of Botany, 2011, 68(7): 1449-1456.
[25] Pearse I S, Koenig W D, Funk K A, et al. Pollen limitation and flower abortion in a wind-pollinated, masting tree[J]. Ecology, 2015, 96(2): 587-593.
[26] 周 洲. 花粉贮藏温度对海枣花粉活力、着果率和果实品质的影响[J]. 中国果业信息, 2018, 35(5): 53-54.
[27] 任转滩, 洪德峰, 马 毅. 低温环境下玉米花粉生活力保持时间的研究[J]. 玉米科学, 2008, 16(4): 199-200.
[28] 宋玉霞, 郭生虎, 牛东玲, 等. 濒危植物肉苁蓉(Cistanche deserticola)繁育系统研究[J]. 植物研究, 2008, 28(3): 278-282, 287.
[29] 李在留, 李雪萍, 郭 松, 等. 珍稀濒危植物掌叶木的开花生物学特性与繁育系统[J]. 园艺学报, 2015, 42(2): 311-320.
责任编辑:白 净
收稿日期 2020-02-20;修回日期 2020-05-18
基金项目 国家自然科学基金项目“海南岛热带雨林特有新物种吊罗山薹草的繁育生物学研究”(No. 31702166)。
作者简介 林 鹏(1994—),男,硕士研究生,研究方向:种质资源保护利用。*通信作者(Corresponding author):杨虎彪(YANG Hubiao),E-mail:yanghubiao@163.com。
