茄子SmWRKY65 基因克隆及其对青枯病的抗性分析
2021-04-23余炳伟李丹丹曹必好
刘 开,余炳伟,李丹丹,陈 娜,曹必好
(1.华南农业大学园艺学院,广东 广州 510642;2.华南农业大学农学院,广东 广州 510642)
【研究意义】茄子(Solanum melongenaL.)是茄科茄属常见的蔬菜作物,起源于东南亚的热带地区,在我国南北各地广泛栽培,其根、茎、叶皆可入药,果实中富含蛋白质、脂肪、维生素、龙葵碱和多种微量元素,既可用作蔬食,又具有降血压、防治胃癌等功效[1],营养价值与药用价值极高。青枯病是茄子栽培中常见的细菌性土传病害之一,其致病因子是青枯雷尔氏菌(Ralstonia solanacearum),主要从根部侵入寄主植物的维管束部位并迅速扩散到地上部组织,对植株可造成毁灭性的破坏[2]。土温20 ℃左右时为茄子青枯病的发病高峰期,微酸性土壤、连作以及气温急剧上升等都会导致青枯病严重发生[3]。我国茄子青枯病发生较为普遍,发病时严重影响茄子的产量和品质,导致茄子大面积减产减收[3]。但茄子抗青枯病的分子机理十分复杂,涉及多种因子调控,迄今仍未研究清楚。因此,深入开展茄子抗青枯病的相关研究显得尤为重要。
【前人研究进展】植物WRKY 转录因子是一种具有多功能调控作用的转录因子,最早是从甘薯〔Dioscorea esculenta(Lour.)Burkill〕的SPF1基因中被鉴定出来的,因参与调控甘薯中的糖信号转导过程而备受关注[4]。之后,陆续从野燕麦(Avena fatuaL.)、欧芹〔Petroselinum crispum(Mill.)Hill〕和水稻(Oryza sativaL.)的基因中成功鉴定出相应的WRKY基因,并详细阐明了其在植物生长发育、信号转导及其抗逆过程中发挥的重要作用[5-7]。近年来,关于WRKY 转录因子响应生物胁迫及非生物胁迫相关过程的研究多有报道[8]。研究发现,小麦TaWRKY2和陆地棉GhWRKY33基因是响应干旱调节的关键基因,过表达TaWRKY2和GhWRKY33基因能够分别增强小麦和转基因拟南芥植株的抗旱性[9-10];决登伟等[11]发现,玉米ZmWRKY25-like基因在响应盐胁迫和低温胁迫中上调表达;Sun 等[12]认为,山葡萄VaWRKY33基因在低温胁迫下诱导表达以增强植株的耐寒性。进一步研究发现,WRKY 转录因子在调控植物抗病性方面起关键作用。周茜茜等[13]研究发现,苹果MdWRKY40基因通过介导水杨酸(Salicylic acid,SA)信号途径正向调控苹果对轮纹病菌的抗性;龙琴等[14]认为,过表达CsWRKY61能够激活与生物胁迫和信号转导相关的途径,增强柑橘对溃疡病的抗性;殷丽华等[15]研究发现,小豆VaWRKY33基因能够响应豇豆单胞锈菌的诱导上调表达,并明显提高小豆锈病的抗性;在拟南芥抗病性研究中发现,拟南芥AtWRKY61和AtWRKY70基因分别正向调控芜菁皱叶病毒和丁香假单胞菌的抗性[16-17];王丹等[18]研究发现,VqWRKY53通过促进SA 和芪类物质的合成,可增强中国野生毛葡萄植株的抗病性。
【本研究切入点】目前已有研究在茄属植物欧白英(Solanum dulcamara)的基因组中克隆得到WRKY基因,并对其相应的生物信息学功能进行了分析[19],但与茄子青枯病抗性相关的研究仍鲜见报道。【拟解决的关键问题】本研究通过克隆与茄子青枯病抗性相关的SmWRKY65基因,对其进行生物信息学、亚细胞定位及时空表达模式分析,并以清水和pTRV2空载为对照,构建pTRV2-SmWRKY65载体,探究SmWRKY65在茄子抗病植株(E-31)中沉默后对茄子青枯病的响应情况,以期为茄子青枯病抗病机理的相关研究提供理论基础,为茄子青枯病抗性品种的选育提供参考。
1 材料与方法
1.1 试验材料
茄子抗病材料(S.melongenaL.E-31,R)和感病材料(S.melongenaL.E-32,S)由曹必好教授提供。青枯菌致病菌株从感病材料中分离得到,VIGS 载体构建所需的烟草脆裂病毒载体pTRV-1和pTRV-2由华南地区园艺作物生物学与种质创制重点实验室保存,DH5α 大肠杆菌转化菌株和GV3101 转农杆菌菌株均购自生工生物科技有限公司。
RNA 提取试剂盒Trizol 购自北京华越洋生物科技有限公司,PCR 反应试剂盒2×HiFiTaq PCR StarMix、快速DNA 胶回收试剂盒StarPrep 和快速质粒小提试剂盒StarPrep 均为GenStar 产品,反转录试剂盒HiScript®ⅡQ Select RT SuperMix for qPCR 和实时荧光定量PCR 试剂盒a SYBR Premix Ex Taq kit 购自南京诺唯赞生物科技有限公司。
1.2 试验方法
1.2.1 茄子不同组织部位总RNA 的提取与cDNA 的合成 在茄子幼苗三叶期时分别采集茄子根、茎、叶部位的样品,用锡箔纸包裹并迅速放入液氮中,每个样品3 次重复并做好标记,之后采用Trizol 试剂盒的方法分别提取茄子根、茎、叶的RNA,并采用HiScript®ⅡQ Select RT SuperMix for qPCR 试剂盒进行逆转录反应合成cDNA,详细步骤参照说明书。cDNA 合成PCR 反应体系10 μL,反应程序为:25 ℃,10 min;50 ℃,30 min;85 ℃,5 min。
1.2.2SmWRKY65序列的获得与基因克隆 本课题组前期通过转录组测序得到WRKY65基因序列(登录号:Sme2.5_00423.1_g00013.1)。利用Primer 5.0 软件设计引物(SmWRKY65-F、SmWRKY65-R,表1),以茄子叶片cDNA 为模板通过PCR(10 μL 体系)扩增得到SmWRKY65基因序列。PCR 反应程序:94 ℃预变性2 min;94 ℃变性30 s、55 ℃退火30 s、72 ℃延伸50 s,32 个循环;72 ℃延伸5 min。采用1%琼脂糖凝胶电泳检测PCR 产物的特异性,将条带明亮、大小与预期一致的条带采用StarPrep 快速DNA 胶回收试剂盒进行纯化。
1.2.3SmWRKY65的生物信息分析 利用ExPASy ProtParam tool(http://www.expasy.org/tools/protparam.html)对SmWRKY65 蛋白理化性质进行分析;使用ProtScale(http://ca.expasy.org/tools/protscale.html)和SOPM 工具(https://npsa-prabi.ibcp.fr/cgibin/npsa_automat.pl?page=np-sa_sopma.html)分别预测SmWRKY65 蛋白亲/疏水性和二级结构类型;使用NCBI CD Search(https://www.ncbi.nlm.nih.gov/Struc-ture/cdd/wrpsb.cgi)和PredictProtein 工具(http://www.predictprotein.org/)对SmWRKY65 蛋白序列进行二级结构的结构域和结合位点预测;通过NCBI Blast 在线比对工具,检索SmWRKY65同源性基因,将相似性较高的同源性序列下载后,利用MEGA 7.0 软件UPGMA 法构建进化树。
1.2.4 PCR 产物的连接与SmWRKY65 质粒的提取 将上述1.2.2 中的PCR 产物连接pMD 19-T载体(pMD 19-T 载体:Solution I :回收产物=0.5 μL :4.5 μL :5 μL),之后转化大肠杆菌感受态DH5α,用pMD19-T 通用引物检测阳性菌落,委托广州擎科生物技术有限公司进行测序,确定重组子。挑取重组子进行扩摇,采用StarPrep 快速质粒小提试剂盒提取SmWRKY65 质粒,具体步骤参照说明书。
1.2.5SmWRKY65的时空表达模式分析 利用Primer 5.0 软件设计SmWRKY65荧光定量引物(q-SmWRKY65-F、q-SmWRKY65-R,表1),以茄子内参18S rRNA(18S rRNA-F、18S rRNA-R,表1)为对照,采用a SYBR Premix Ex Taq Kit 试剂盒对茄子根、茎、叶组织进行SmWRKY65实时荧光定量(qRT-PCR)表达量的测定,详细步骤参照说明书。再分别以茄子抗病和感病植株为材料,在茄子苗3 叶期接种青枯病菌以分析SmWRKY65在茄子抗病和感病植株中的表达情况,并在接种后0、3、6、9 h 4 个时间段分别采集抗病、感病植株叶片,以茄子内参18S rRNA(18S rRNA-F、18S rRNA-R,表1)为对照,采用SmWRKY65荧光定量引物(q-SmWRKY65-F、q-SmWRKY65-R,表1)进行qRT-PCR 表达量的测定,以分析SmWRKY65在抗病和感病材料中的表达量差异。qRT-PCR 程序为:95 ℃预变性2 min;95 ℃变性10 s、56 ℃退火20 s、72 ℃延伸35 s,35 个循环。每个样品3 次生物学重复,最后取平均值进行分析,基因的相对表达量应用2-ΔΔCt 方法进行分析[20]。

表1 引物序列Table 1 Primer sequences
1.2.6 构建亚细胞定位载体及转化洋葱 利用Primer 5.0 软件设计SmWRKY65亚细胞定位引物(SmWRKY65-GFP-EB-F、SmWRKY65-GFPEB-R,表1),通过PCR 反应克隆SmWRKY65亚细胞定位目的片段。使用EcoR I 和BamH I 酶对亚细胞定位载体pC018 和目的片段进行双酶切并连接成重组质粒SmWRKY65-GFP,测序后将重组质粒转入农杆菌GV3101,转化洋葱内表皮组织,3 d 后在显微镜下观察亚细胞定位结果。
1.2.7 VIGS 载体的构建 通过在线工具(vigs.solgenomics.net)寻找特异性的VIGS 载体区域,根据pTRV2载体多克隆位点所包含的酶切位点,设计1 对特异性引物(pTRV2-WRKY65-F、pTRV2-WRKY65-R,表1),以茄子叶片cDNA为模板进行PCR 反应获得长度为250 bp 左右的特异性PCR 产物,用双酶切(EcoR I 和BamH I)法将产物片段连接到pTRV2载体上,之后转入大肠杆菌DH5α,阳性单菌落进行PCR 菌落筛选,送广州擎科生物科技有限公司测序,确定重组子。将阳性单菌落进行扩摇,提取重组质粒(pTRV2-SmWRKY65)后转入农杆菌GV3101 用于侵染。
1.2.8 农杆菌侵染茄子幼苗 将抗病和感病茄子种子分别播种在含泥炭和珍珠岩的混合基质中,于25 ℃、16 h 光照和8 h 黑暗的培养室中进行培养。当茄子幼苗长至3~4 片真叶时,吸取含有重组质粒(pTRV2-SmWRKY65)的阳性菌落扩摇后配制成侵染液,并将分离到的青枯菌采用伤根断根法对茄子幼苗进行侵染,青枯菌的分离与接种参照曹必好等[21]的方法,每组15 株,3 次重复,之后在16 ℃、60 %的相对湿度下暗处理1 d,随后置于培养室中生长。3 d 后采用断根法接种青枯病菌,观察其表型变化并拍照,2 周后调查统计病情指数,方法参照Chen 等[22],之后通过qRT-PCR 法测定pTRV2-SmWRKY65沉默植株表达量。
试验数据采用SPSS Statistics 17.0 进行t检验,采用Excel 2010 制图。
2 结果与分析
2.1 SmWRKY65 基因的克隆
以茄子叶片cDNA 为模板,通过PCR 扩增得到948 bp 的SmWRKY65片段,经1%琼脂糖凝胶电泳,结果显示目的条带与预期结果一致(图1),将目的条带切胶回收、纯化、测序。对SmWRKY65测序序列与WRKY65原始序列(登录号:Sme2.5_00423.1_g00013.1)在DNAMAN 8.0工具上进行比对,结果显示同源性为88.06%,表明在茄子中成功克隆了SmWRKY65(图2)。

图1 SmWRKY65 电泳结果Fig.1 Electropherogram result of SmWRKY65
2.2 SmWRKY65 蛋白的生物信息学分析
SmWRKY65 蛋白理化性质分析结果显示,该基因开放阅读框为834 个核苷酸序列,编码277 个氨基酸残基,蛋白相对分子量大小为30155.21 ku,等电点为5.51,分子式为C1304H1998N30O430S13,原子总量为4115,预测该蛋白属于亲水性蛋白。对SmWRKY65 蛋白的二级结构进行预测,结果显示,其主要由不规则盘绕结构(72.92%)、α-螺旋(12.27%)、延伸链(11.91%)和β-转角(2.89%)等结构元件组成(图3)。预测该蛋白属于WRKY 家族,在71~131 区域具有1 个含60 个氨基酸的WRKY 保守域,可以特异结合DNA 序列(T)(T)TGAC(C/T),具有DNA 结合位点3 个、蛋白结合位点3 个、核苷酸多肽链结合位点2 个、大分子结合区域7 个,主要多集中在WRKY 家族的保守结构域内,预测该基因定位于细胞核中(图4)。通过NCBI 上Blast 在线比对工具,将检索到的12条与SmWRKY65同源性较高的基因进行多序列比对,使用MEGA 7.0.21 软件构建进化树,结果表明,茄子SmWRKY65与马铃薯、番茄的WRKY65同源性最高(图5)。
2.3 SmWRKY65 的时空表达模式和亚细胞定位分析
以茄子叶片cDNA 为模板,通过PCR 检测SmWRKY65荧光定量引物的特异性,经1%琼脂糖凝胶电泳,结果显示在250 bp 左右有1 条明亮且符合预期的条带(图6A),说明引物特异性较好,可用于后续试验。对茄子根、茎、叶不同组织部位表达量的分析结果显示,SmWRKY65在茄子根组织部位中的表达量最高,在叶中的表达量次之,茎中最低(图6B)。在茄子苗期接种青枯病菌后0、3、6、9 h 4 个时间段分别采集抗病、感病植株叶片进行实时荧光定量分析,结果表明,SmWRKY65在抗病和感病材料中的表达量均随接种时间延长逐渐升高,9 h 时达到最高,且在抗病材料中的表达水平明显高于在感病材料中的表达水平(图6C),说明SmWRKY65响应茄子青枯病抗性相关基因的表达。
使用EcoR I 和BamH I 酶对亚细胞定位载体pC018 和亚细胞定位引物成功克隆出的PCR 片段进行双酶切,并连接成大小为831 bp 的重组质粒SmWRKY65-GFP(图7A),将重组质粒转化农杆菌后转化洋葱表皮细胞,在共聚焦显微镜下观察到SmWRKY65定位在细胞核中表达(图7B),这与预测结果一致。

图6 茄子不同组织及抗感病植株中SmWRKY65 的表达分析Fig.6 SmWRKY65 expression of different tissues and disease-resistant and disease-susceptible plants of eggplant

图7 SmWRKY65 亚细胞定位分析Fig.7 Subcellular localization analysis of SmWRKY65
2.4 SmWRKY6 沉默在抗病茄子中响应青枯病的情况
2.4.1pTRV2-SmWRKY65载体构建结果 以茄子叶片为材料提取RNA 并通过逆转录反应合成cDNA,再以cDNA 为模板,利用构建VIGS 载体所设计的特异性引物进行PCR 扩增,1%琼脂糖凝胶电泳显示,获得长度为242 bp 的PCR 产物(图8A),将产物片段和pTRV2 载体同时用EcoR I和BamH I 进行双酶切后成功将SmWRKY65连接到pTRV2载体上获得重组质粒,经测序保证连接片段和读码方向正确后转入农杆菌GV3101。从阳性菌落检测结果来看,与pTRV2(图8B1)空载相比,pTRV2-SmWRKY65(图8B2)载体构建成功。
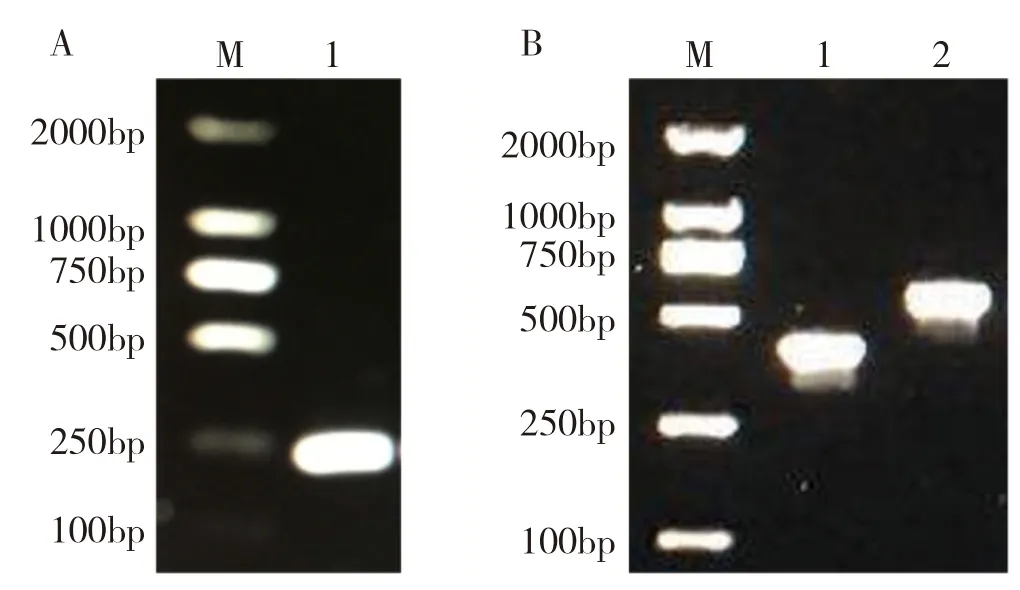
图8 pTRV2-SmWRKY65 载体构建Fig.8 pTRV2-SmWRKY65 vector construction
2.4.2 青枯病菌接种VIGS 沉默植株 为了验证pTRV2-WRKY65沉默植株对茄子青枯病的响应情况,在3~4 片真叶时,对茄子抗病植株接种青枯病病原菌。接种7 d 后观察到接种清水的叶片均没有表现出明显的萎蔫症状,接种pTRV2空载的植株仅有1 片叶片出现萎蔫,而接种pTRV2-WRKY65的15 株植株几乎全部表现出萎蔫症状;接种2 周后,对pTRV2-WRKY65植株进行病情指数调查,结果(表2)表明,SmWRKY65基因被沉默后影响了抗病材料对青枯病的抵御能力,即表现为感病症状。pTRV2-SmWRKY65实时荧光定量检测结果(图9)显示,与清水和pTRV2空载相比,pTRV2-SmWRKY65表达量明显下降。

表2 VIGS 沉默植株的病情指数统计Table 2 Statistics of disease index of VIGS silent plants

图9 VIGS 沉默植株的表型(A)及qRT-PCR 分析(B)Fig.9 Phenotype(A)and qRT-PCR analysis(B)of VIGS silent plants
3 讨论
茄子是我国南北各地广泛栽培的夏秋季重要蔬菜之一,在我国的栽培历史悠久,种植面积大,具有较高的营养价值和经济价值。但茄子易受青枯病侵袭,尤其在我国南方地区,其病原菌适应性强、分布广泛、繁殖速度较快,常规方法难以有效防治,而抗病育种可以有效防御病菌的危害,能较大幅度提高茄子的产量和品质[2,23]。近年来,研究人员在提高茄子青枯病抗性及抗病品种选育方面作了较多努力。邵欣欣[24]在茄子青枯病抗性研究中发现,ERF 转录因子参与调控茄子抗青枯病的信号途径;肖熙鸥等[25]研究认为,SA 信号途径中MAPK6、MKK2、NPR1、PAD4等基因能够正向调控茄子青枯病的抗性;李威等[26]研究发现,1.5~2.0 mmol/L 的硅浓度处理可有效提高快圆茄青枯病的抗性;Qiu 等[27]认为,过表达SmMYB44可诱导茄子中亚精胺相关基因(SmSPDS)的表达,促进亚精胺的合成与积累,从而增加对茄子青枯病的抗性。此外,广州市农业科学研究院经过多年研究选育出的翡翠绿2 号青茄新品种在大田抗病性鉴定中表现为中抗青枯病[28],与广东省农业科学院植物保护研究所联合选育出的紫荣6 号和玫瑰紫花茄均为抗青枯病的优质茄子新品种[29-30]。
研究表明,在生物胁迫下,植物体内会通过信号传导途径和防御反应作出应激防卫,而转录因子在这一系列反应中起关键作用[31]。近年来有关WRKY 转录因子在植株抗病性上的研究成为一个热门话题[32]。已有研究报道,WRKY 转录因子涉及调控与青枯病抗性相关的信号转导过程。王鹏飞等[33]研究发现,野生花生AdWRKY37基因在受到青枯病菌液胁迫诱导后表达量明显升高,并推测该基因可能通过介导水杨酸(Salicylic acid,SA)和茉莉酸(Jasmonic acid,JA)途径参与青枯病抗病相关过程的调控;Dang 等[34]认为,CaWRKY27 转录因子通过调节SA、JA 和内皮素(Endothelin,ET)介导的信号通路,可增强烟草对青枯菌的抗性;Liu 等[35]发现,在烟草中过表达NtWRKY50能够促进SA 合成,从而增强植株对青枯病的抗性;Dang 等[36]在辣椒抗病育种的研究中发现,CaWRKY41 转录因子能够提高辣椒对青枯菌的抗性。然而,有关WRKY 转录因子在茄子青枯病抗性中的相关研究还鲜见报道。本研究以茄子叶片cDNA 为模板成功克隆了SmWRKY65基因,与茄子数据库中原始WRKY65序列比对同源性为88.06%,序列差异性的产生可能与PCR 扩增过程中单核苷酸碱基变异及引物二聚体的形成有关。实时荧光定量分析结果表明,SmWRKY65在茄子抗病和感病植株中均响应青枯病的诱导,其表达量在接种青枯病菌后0、3、6、9 h 4 个时间点不断升高,且SmWRKY65在茄子抗病材料中的表达水平明显高于感病材料,说明SmWRKY65可能涉及茄子青枯病抗病的调节过程。在抗病茄子(E-31)植株中接种青枯菌的分析结果显示,与清水和pTRV2空载相比,SmWRKY65沉默植株表现出明显的萎蔫、易感症状,且qT-PCR 分析表明SmWRKY65在茄子中的表达量较清水和pTRV2空载明显降低,这表明SmWRKY65沉默后可能抑制了茄子青枯病抗病信号途径中相关基因的表达,从而降低了茄子植株对青枯病原菌的抗性。
4 结论
本研究成功克隆了SmWRKY65基因,与茄子数据库中原始WRKY65序列的同源性为88.06%,该基因开放阅读框为834 个核苷酸序列,编码277 个氨基酸残基,在71~131 区域包含1 个含60 个氨基酸的WRKY 保守域,定位于细胞核中,与马铃薯、番茄WRKY65的同源性最高。时空表达分析发现,SmWRKY65在茄子根组织中的表达量最高,叶中次之,茎中最低。实时荧光定量PCR 分析发现,SmWRKY65在茄子抗病和感病材料中均响应青枯病菌的诱导上调表达,且在抗病材料中的表达水平明显高于感病材料;VIGS 结果表明,接种pTRV2-SmWRKY65植株的叶片表现出明显的萎蔫症状,结合qRT-PCR 分析发现,pTRV2-SmWRKY65表达量较对照明显下降,表明SmWRKY65沉默后茄子抗青枯病的能力下降,说明SmWRKY65可能涉及茄子青枯病抗病相关过程的调控。
