低温对不同菠萝蜜种质资源光合生理特性的影响
2021-04-02宋奇琦朱鹏锦何江叶维雁欧景莉唐秀观李佳慧程琴谭秦亮庞新华欧克纬卢业飞
宋奇琦 朱鹏锦 何江 叶维雁 欧景莉 唐秀观 李佳慧 程琴 谭秦亮 庞新华 欧克纬 卢业飞

摘 要:菠萝蜜是一种典型热带果树,有巨大的潜在开发前景。低温寒害已成为制约菠萝蜜产业健康可持续发展的关键问题,也是菠萝蜜耐寒品种选育及生理生态研究重点关注的焦点。为了研究低温寒害对菠萝蜜叶片光合作用能量耗散过程以及光合色素影响,通过连续的气温数据监测、表征观测、叶绿素荧光参数及光合色素分析,结果表明4个菠萝蜜种质资源耐寒性强弱依次是博白菠萝蜜(A)>湛江菠萝蜜(B)>马来西亚菠萝蜜(C)>泰国菠萝蜜(D),低温对不耐寒种质的光系统损伤较大,叶绿素荧光参数、叶绿素含量均明显较耐寒型种植低。研究耐寒型菠萝蜜种质光合生理如何响应低温的机理,将是选育耐寒新品种和寒害防控的重点方向。
关键词:低温;菠萝蜜;叶绿素荧光成像;光合色素
中图分类号:S667.8 文献标志码:A
Effect of Low Temperature on Photosynthetic Physiological Characteristics of Different Artocarpus heterophyllus Lam. Germplasm Resources
SONG Qiqi,ZHU Pengjin*,HE Jiang,YE Weiyan,OU Jingli,
TANG Xiuguan,LI Jiahui,CHENG Qin,TAN Qinliang,
PANG Xinhua,OU Kewei,LU Yefei
(Guangxi Subtropical Crops Research Institute,Nanning,Guangxi 530001,China )
Abstract: Artocarpus heterophyllus Lam. (Jackfruit) is a typical tropical fruit tree with great potential development value. Cold damage has become the key problem restricting the healthy and sustainable development of Jackfruit industry,and is also the focus of attention in breeding of cold-resistant variety and in physiological and ecological research of jackfruit. Based on continuous temperature data monitoring,characterization,chlorophyll fluorescence parameters and photosynthetic pigments analysis,the effect of chilling damage on the energy dissipation process and photosynthetic pigments of jackfruit leaves was studied. Results showed that the order of cold hardiness of the four jackfruit germplasm resources was GXRZBLM00051(A)>GXRZBLM00039 (B)>GXRZBLM00006(C)>GXRZBLM00013(D) Low temperature had greater damage to the photosystem of cold-intolerant germplasm,and the chlorophyll fluorescence parameters and chlorophyll content of cold-intolerant germplasm were significantly lower than those of cold-tolerant germplasm. To study the mechanism of how photosynthetic physiology of cold-tolerant jackfruit germplasm responds to low temperature will be the key research direction for breeding new cold-tolerant varieties and preventing and controlling cold damage
Key words:Low temperature;Artocarpus heterophyllus Lam.;chlorophyll fluorescence imaging;photosynthetic pigment
菠蘿蜜(Artocarpus heterophyllus Lam.) 属于桑科菠萝蜜属植物,为典型热带果树,主要分布于印度、印度尼西亚、菲律宾、马来西亚、泰国等热带国家,而在我国的海南、广东、广西、云南等热带亚热带省(区)均有栽培,以海南省、广东湛江和广西玉林栽培较多[1, 2]。菠萝蜜不仅香味独特、果肉蜜甜且营养丰富,种子淀粉含量高可加工成各种食品,果皮经过加工处理可作为一种营养丰富的牛羊养殖饲料,而且根、树皮提取的活性成分可入药[3-5]。此外,木材还是很好的家具材料。鉴于此,其消费者群体迅速增长,市场需求日益增大,且在科研领域以及林木种植领域的研究与应用也日益受到人们的重视。菠萝蜜的生长发育对温度要求较高,喜炎热湿润的热带气候,不耐寒冷,要求种植地的年平均气温≥21℃,最冷月平均气温≥13℃,绝对最低温度≥0℃;温度低于5~7℃时易于落花落果。植物可以通过调控基因和蛋白质等的表达来适应胁迫,在长时间低温下仍能存活,植物器官和各代谢途径中的低温损伤程度可能不同,但几乎影响到所有的光合作用元件[6] 。广西极端低温时常发生菠萝蜜遭受寒害[7, 8],已成为制约菠萝蜜产业高质量发展的关键问题。目前,菠萝蜜的研究主要集中在种质资源的调查与收集[9, 10]、植物学性状评价[11]、营养品质[12]、高效栽培技术等方面[13],而针对菠萝蜜响应低温胁迫的光合生理特性研究尚未有报道。本研究以抗寒能力不同的菠萝蜜种质为研究对象,通过叶绿素荧光图像分析系统(CF Imager; Technologica, UK)探讨菠萝蜜叶片光合作用能量耗散过程对低温的响应,并测定其叶绿素含量,以期为评价和选育耐寒菠萝蜜优良新品种提供理论依据和实验基础。
1 材料与方法
1.1 材料与试验地点
参试种质材料分别是博白菠萝蜜(GXRZBLM00051,编号为A)、湛江菠萝蜜(GXRZBLM00039,编号为B)、马来西亚菠萝蜜(GXRZBLM00006,编号为C)、泰国菠萝蜜(GXRZBLM00013,编号为D)4个保存于广西壮族自治区亚热带作物研究所菠萝蜜种质资源圃的核心种质。该种质圃位于广西中部(N22.899°E108.343°,广西南宁),海拔95.2 m,年平均氣温 16.0~23.0℃,累年极端最高气温为33.7~42.5℃,累年最低气温为-8.4~2.9℃,冬季最冷的1月平均气温12.8℃,夏季最热的7、8月平均气温28.2℃,年降水量均在1304.2 mm以上,平均相对湿度为79%,属湿润的亚热带季风气候。参试菠萝蜜材料为3~6年生嫁接树,定植株行距5 m × 5 m,树形常见对生枝干的塔形。
1.2 试验方法
1.2.1 菠萝蜜树冠层温度监测
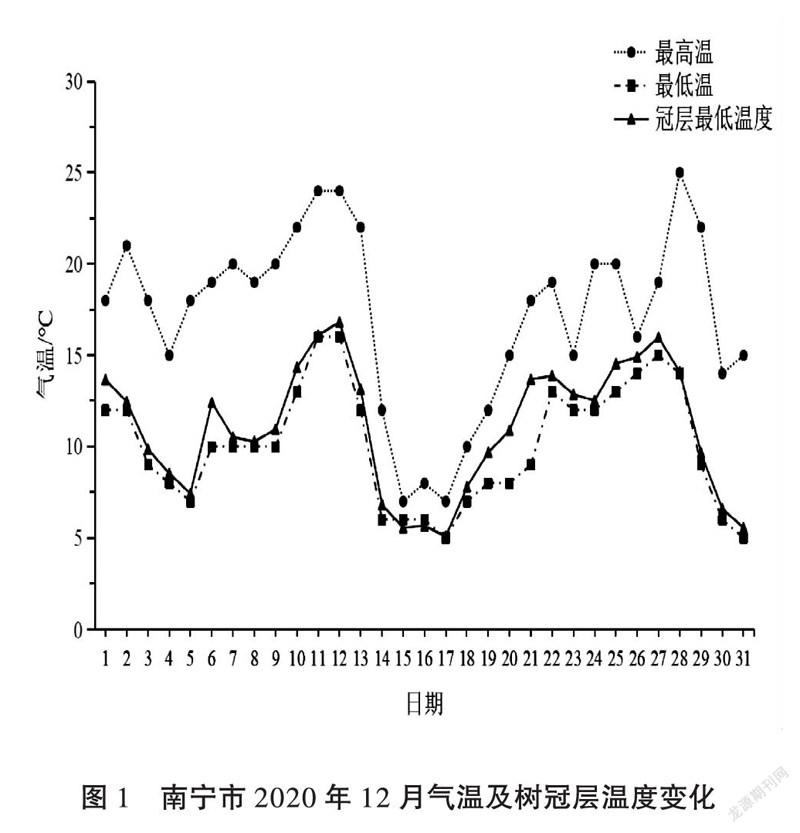
根据2020年12月天气预告广西将出现大幅度降温且可能出现极端低温,对菠萝蜜种质圃开展近地面温度的监测及菠萝蜜树寒害情况的调查,选择4个不同菠萝蜜种质资源进行树冠层温度的监测。温度记录仪(COS-04,建大仁科)测温探头统一固定于菠萝蜜树冠西北方向距地面2.0 m高,温度记录时间间隔为30 min,每隔7 d采集1次温度数据,监测时间为2020年12月1日至12月31日。同时,收集野外气象台站监测的气温数据。
1.2.2 叶绿素荧光测定
叶绿素荧光图像分析系统具有快速、灵敏、无损伤测定样品叶绿素荧光参数的优点,通过荧光成像来反映叶片光合状态。本试验在经过自然低温胁迫后,采摘不同菠萝蜜种质的+1叶(每个树枝上从上往下数第一片叶子),进行菠萝蜜叶片的叶绿素荧光参数分析。根据 CF Imager 使用说明,叶片经暗处理30 min后, 测定初始荧光(Fo)、最大荧光(Fm) ;充分光适应30 min后, 测定光适应下的最小荧光Fo′、最大荧光Fm′、稳态荧光F′,Fv'/Fm'、NPQ、Fq'/Fm'等荧光参数由软件控制并自动计算。
1.2.3 叶绿素含量测定
叶片测定叶绿素荧光参数后,每个种质资源选取3张叶片避开主脉用打孔器(ΦA=6 mm)取18个圆片,混合后随机分成3组,每组6片并称重。然后将叶圆片分别放入装有10 mL叶绿素提取液(丙酮∶乙醇∶水=45∶45∶10)的试瓶中,密封瓶盖并编号标记,密封低温保存至叶圆片无色约10 d。采用紫外分光光度计(UV1800, GREEN)分别测定663 nm和645 nm波长下的吸光度。叶绿素含量的测定参照Arnon[14]计算:
叶绿素 a(Chl a)=(12.78A663-2.69A645)×V/ 1000W
叶绿素 b(Chl b) =(22.9A645-4.68A663)×V/ 1000W
叶绿素总含量(Chl)= Chla+Chlb
式中,A为吸光值,V为提取液总体积(mL),W为叶片鲜重(g),叶绿素含量单位为 mg/g。
1.3 数据处理与统计学分析
原始数据的计算与处理采用 Excel 2007软件,用IBM SPSS Statistics 26进行统计分析,各种质资源间的差异采用单因素方差分析(One-way ANOVA),多重比较采用最小显著差异法(LSD),显著性水平设定为α=0.05,使用Excel 2007和Origin2019作图。
2 结果与分析
2.1 菠萝蜜树冠层温度与日平均温度低温差异
据广西气象局的气温数据,2020年12月气温变化(图1),其中日最高气温为29℃,日最低气温为5℃。菠萝蜜正常生长发育的最冷月平均气温不低于13℃,然而自13日开始天气由阴转小雨气温急剧下降,日最高气温为24℃,日最低气温为12℃;到了17日气温降至12月份最低,其日最高气温为7℃,日最低气温为5℃;直到21日天气多云转晴气温开始快速回升,其日最高气温为18℃,日最低气温为9℃。同期实测的菠萝蜜树冠层低温与气象日最低温的变化趋势基本一致。
2.2 低温胁迫后不同种质资源叶片表型特征
在低温过后,不同品种受到胁迫程度表征不同(图2)。博白菠萝蜜(A)的叶片仍呈深绿色,叶表面和嫩芽无明显受损特征;湛江菠萝蜜(B)的叶片在低温过后叶片呈绿色,但在叶片边缘及局部出现小型点状褐色斑点且嫩芽出现少量褐斑;马来西亚菠萝蜜(C)的叶片呈黄色,叶表面出现大面积浅褐色冻斑且叶缘部分卷曲,嫩芽呈现黄绿色并出现部分褐色冻斑;泰国菠萝蜜(D)的叶片大面积深褐色冻斑并出现明显卷曲,嫩芽直接枯死。不同种质对低温寒害的耐受能力不同,从叶片和嫩芽表征判断,耐寒性强弱依次是:A>B>C>D。
图2 低温寒害下不同菠萝蜜种质资源叶片和嫩芽的表型特征
2.3 低温胁迫后不同菠萝蜜种质资源的叶绿素荧光参数变化
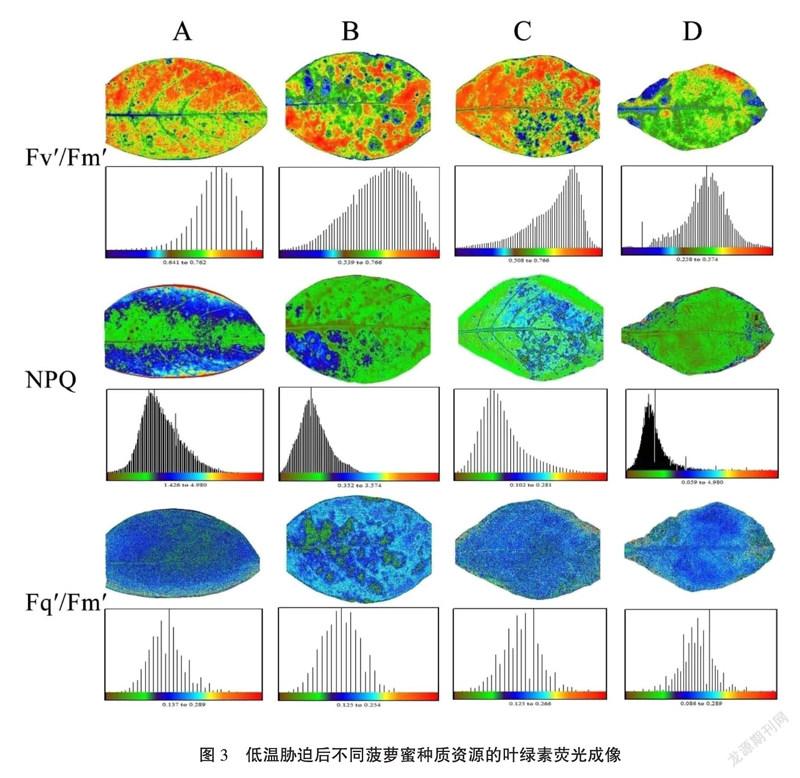
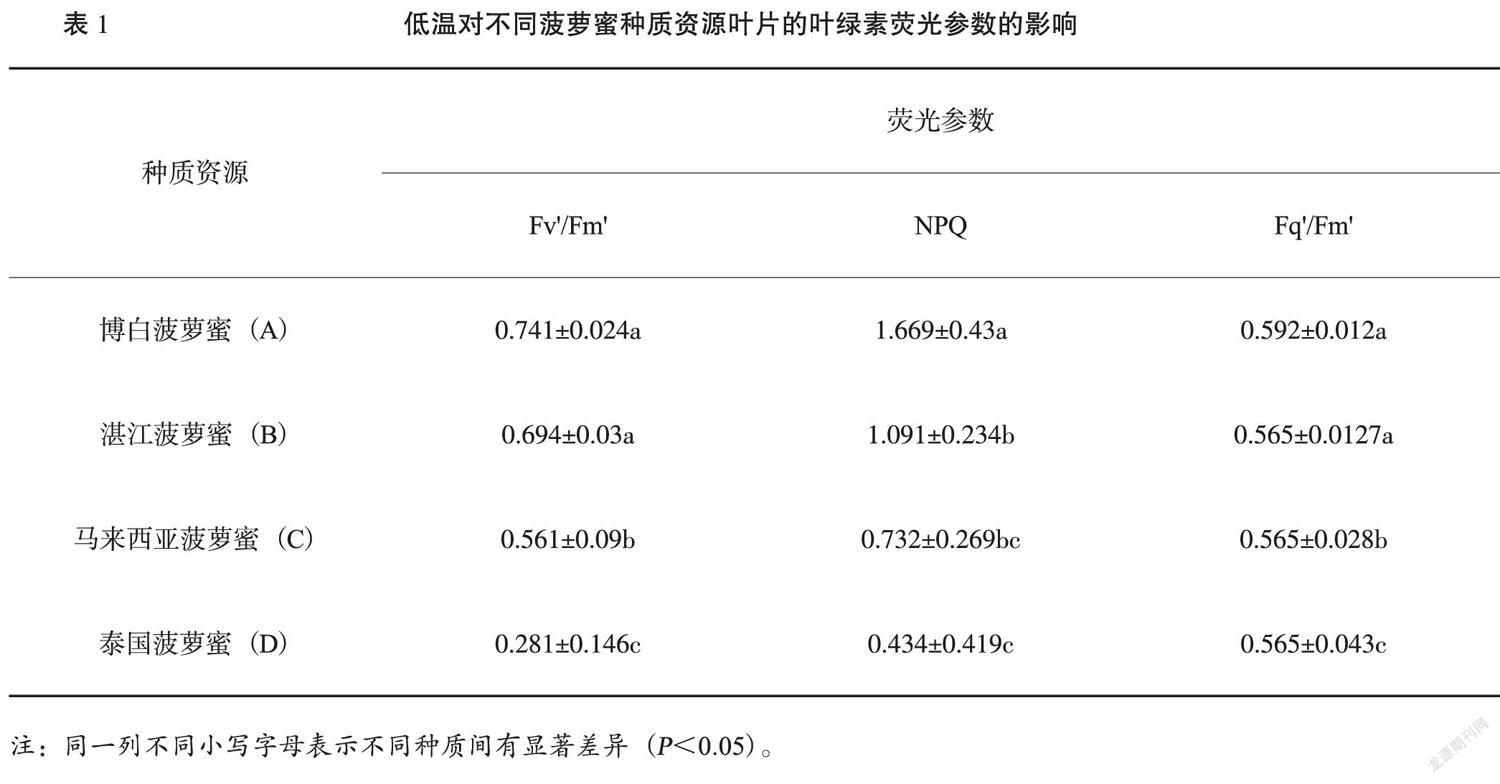
虽然不同种质资源的抗寒能力有差异,但在低温胁迫的早期表型判断不同种质的耐寒性难度很高。而叶绿素荧光图像分析系统不仅能快速且无损伤地探测光合机构 (PSII) 反应中心的光化学量子效率的响应,还具有全株荧光图像分析功能是研究胁迫生理的有力工具[15]。如图3所示,在低温胁迫后不同种质叶片的叶绿素荧光参数的成像具有明显差异,如博白菠萝蜜(A)其叶片Fv'/Fm'、NPQ、Fq'/Fm'的图像颜色最深,且不同耐寒性的种质资源其参数成像具有明显差异。其中,根据颜色与数值对应关系,叶片Fv'/Fm'、Fq'/Fm'的图像颜色具有相同变化趋势:种质资源A>B>C>D;而NPQ的变化趋势正好相反:种质资源D>C>B>A。
2.4 低温胁迫对不同菠萝蜜种质叶片光合色素的影响
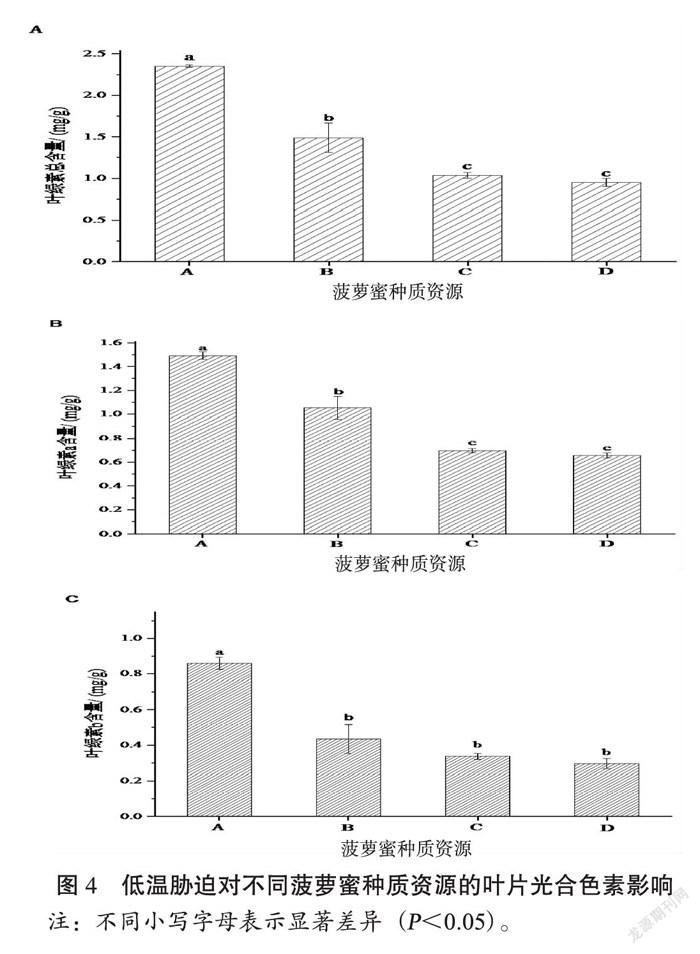
植物叶片光合色素参与光能的吸收、传递和转化过程,是反映环境胁迫状况的重要指标[16]。如图4所示,在低温胁迫后不同菠萝蜜种质叶片的叶绿素受到影响程度有明显差异。其中,叶片总叶绿素含量最高的是博白菠萝蜜(A),其次是湛江菠萝蜜(B),而马来西亚菠萝蜜(C)和泰国菠萝蜜(D)彼此间叶片总叶绿素含量没有明显差异,但其显著低于博白菠萝蜜(A)和湛江菠萝蜜(B)(图4A),结合4个种质资源的叶片表征(图2)说明低温胁迫对菠萝蜜的叶绿素具有明显的破坏;作为将光能转化为生物能的主要色素,叶绿素a的含量差异在低温胁迫下不同种质资源间存在明显的差异,田间寒害表征明显的马来西亚菠萝蜜(C)和泰国菠萝蜜(D)其叶绿素a含量分别是0.698 mg/g,0.655 mg/g与博白菠萝蜜(A)相比较,分别占46.8%,43.9%,其趋势与总叶绿素变化一致(图4B);叶绿素b主要结合于捕光色素蛋白复合体(LHCII),在低温寒害后博白菠萝蜜(A)的叶绿素b含量显著高于其它3个种质,且湛江菠萝蜜(B)、马来西亚菠萝蜜(C)和泰国菠萝蜜(D)彼此间没有显著差异(图4C)。
3 讨论与结论
菠萝蜜越来越受消费者青睐,市场需求不断增长,有巨大的潜在开发前景。目前,广西的菠萝蜜种植面积位居全国第三[17],但因其独特地理位置时常在冬季出现连续低温阴雨或者春季出现“倒春寒”[18],低温寒害导致菠萝蜜幼树出现黄化、枯死、生长迟缓,挂果树的嫩枝黄化及局部枯死、果实冻伤、品质下降等,已成为广西菠萝蜜种植中主要的自然灾害之一。
在低溫寒害后,叶绿素荧光参数显示Fv'/Fm'在博白菠萝蜜(A)和湛江菠萝蜜(B)两者间无显著差异,泰国菠萝蜜(D)的光适应下PSⅡ反应中心的激发能捕获效率最低,说明PSⅡ反应中心受到低温抑制,且不同品种之间受胁迫程度不同;荧光参数Fq'/Fm'表示光适应下PSⅡ实际光化学量子效率,博白菠萝蜜(A)和湛江菠萝蜜(B)彼此间的效率无显著,泰国菠萝蜜(D)的效率与其余3个相比差异显著,反映了低温抑制导致敏感型种质用于光化学途径激发能的比例降低;荧光参数NPQ在4个种质中,博白菠萝蜜(A)的系数最高,其耗散过剩激发能的能力最强,而泰国菠萝蜜(D)的系数最低,其耗散过剩激发能的能力最弱,表明低温导致其热耗散保护能力下降。虽然不同菠萝蜜种质间存在一定差异,但调查结果发现耐寒型种质博白菠萝蜜(A)和湛江菠萝蜜(B)的Fv'/Fm'和Fq'/Fm'差异不明显,但NPQ系数相对较大;而敏感型种质间马来西亚菠萝蜜(C)和 泰国菠萝蜜(D)不仅Fv'/Fm'和Fq'/Fm'参数受到抑制且NPQ系数也受到明显抑制,气温回升后造成严重的自由基失衡导致叶片黄化甚至枯死。这与王玉贤等[19]对低温胁迫下不同耐寒性叶用芥菜的叶绿素荧光参数分析基本一致。
光合色素不仅是作为判断植物光合生理能力,也是反映环境胁迫状况的重要指标[20]。许多研究表明[21-23] ,叶绿素含量往往与植物的抗寒能力具有一定的相关性,抗寒能力强的植物叶片叶绿素含量高于抗寒能力弱的植物,而较高的叶绿素含量有助于可溶性糖等渗透调节物质的产生,进而降低细胞冰点,增强植株的抗寒性[24] 。进一步分析低温寒害后不同菠萝蜜种质间的叶绿素含量,发现博白菠萝蜜(A)叶片叶绿素a、叶绿素b及总叶绿素的含量最高且耐寒性相对较强,而泰国菠萝蜜(D)的含量最低其叶片寒害表征最明显。这说明在低温胁迫下,低温敏感型菠萝蜜种质不仅因叶绿素合成前期的关键酶受到抑制导致含量降低,还因光合机构的光化学途径激发能利用减少而能量出现相对过剩引发光氧化,即光合机构受损,叶绿素降解(漂白),最后甚至导致叶片死亡。有研究证明遭受低温强光胁迫的植物,由于低温的抑制加大了光能过剩的程度,妨碍了保护和修复系统的正常运转,更易遭受光氧化的伤害[25, 26]。
通过连续的气温数据监测、表征观测、叶绿素荧光参数及光合色素分析,发现低温对不耐寒菠萝蜜种质的光系统损伤较大。因此,深入探究耐寒型菠萝蜜种质光合生理如何响应低温的机理是未来耐寒新品种选育和寒害防控的重点研究方向。本试验在菠萝蜜光合生理响应低温的分析中存在一定的局限性:从光合色素含量以及叶绿素荧光参数进行分析,不能够系统、全面地分析光合作用过程中色素的具体功能;从菠萝蜜表征判断或者筛选耐寒种质的方法,不能够快速选育、有效评价种质的耐寒性。后续研究重点是:自然条件下观测评价菠萝蜜相同(不同)种质的植物学性状和农艺性状,同时模拟低温胁迫,深入全面探讨其表征、叶片结构、生理生化及基因表达调控的响应机理。
参考文献
[1] 中国科学院中国植物志编辑委员会. 中国植物志第23卷第1册桑科[M]. 北京:科学出版社, 1998: 44.
[2] 苏兰茜, 白亭玉, 吴刚, 等. 菠萝蜜栽培研究现状及发展趋势[J]. 热带农业科学, 2019, 39(1): 10-15,41.
[3] 张彦军, 王少曼, 左慧玉, 等. 菠萝蜜主要功能活性成分及其研究进展[J]. 食品科学技术学报, 2019,37(6): 16-22.
[4] 贺书珍, 吴刚, 张彦军, 等. 不同基因型菠萝蜜种质资源挥发性香气成分分析[J]. 热带作物学报, 2019, 40(7): 1304-1311.
[5] 师江, 曹海燕, 郭芬. 菠萝蜜籽营养成分分析与安全性评价[J]. 食品研究与开发, 2018,
39(14): 193-196.
[6] Batista-Santos P, Lidon F C, Fortunato A, et al. The impact of cold on photosynthesis in genotypes of Coffea spp.—Photosystem sensitivity, photoprotective mechanisms and gene expression. Journal of Plant Physiology, 2011,168(8):792-806.
[7] 吕庆芳, 李映志, 余伟, 等. 持续低温引起菠萝蜜田间寒害症状调查及抗寒性分析[J]. 果树学报, 2012, 29(1): 81-85,157.
[8] 赵跃宾, 陈斌源, 王伟, 等. 菠萝蜜种植气象因子分析[J]. 福建热作科技, 2018, 43(1):
52-54.
[9] 范鸿雁, 王祥和, 华敏, 等. 海南菠萝蜜种质资源调研与收集评价[J]. 中国热带农业, 2012, 9(3): 34-39.
[10] 陆玉英, 苏伟强, 阮经宙, 等. 广西菠萝蜜种质资源调查与评价[J]. 中国热带农业, 2011, 8(3): 31-32.
[11] 叶春海, 吴钿, 丰锋, 等. 菠萝蜜种质资源调查及果实性状的相关分析[J]. 热带作物学报,
2006, 27(1): 28-32.
[12] 张涛, 潘永贵,菠萝蜜营养成分及药理作用研究进展[J]. 广东农业科学, 2013, 40(4):
88-90,103.
[13] 杜伟杰. 菠萝蜜大苗的移植栽培技术[J]. 安徽农学通报, 2008, 14(23): 233,136.
[14] Arnon, D I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris[J]. Plant Physiology, 1949, 24(1): 1-15.
[15] Barbagallo R P K, Oxborough K E, Pallett, et al. Rapid, noninvasive screening for perturbations of metabolism and plant growth using chlorophyll fluorescence imaging[J]. Plant Physiology, 2003, 132(2): 485-493.
[16] 孙小玲, 许岳飞, 马鲁沂, 等. 植株叶片的光合色素构成对遮阴的响应[J]. 植物生态学
报, 2010, 34(8): 989-999.
[17] 吴刚, 陈海平, 桑利伟, 等. 中国菠蘿蜜产业发展现状及对策[J]. 热带农业科学, 2013,
33(2): 91-97.
[18] 李宗锴, 刘学敏, 杨绍琼, 等. 云南省河口县菠萝蜜寒害情况调查及适应性分析[J]. 热带农业科学, 2019, 39(6): 46-51.
[19] 王玉贤, 姚培杰, 袁颖辉,等.低温胁迫下不同耐寒性叶用芥菜的生理差异分析[J].中国蔬菜, 2021(6): 74-79.
[20] 朱鹏锦, 庞新华, 谭秦亮, 等. 低温胁迫对不同甘蔗品种叶片光合色素和光合生理特性的影响[J]. 热带作物学报, 2019, 40(5):
875-881.
[21] Lin D,Zhang L,Mei J,et al. Mutation of the rice TCM12 gene encoding 2,3-bisphosphoglycerate-independent phosphoglycerate mutase affects chlorophyll synthesis,photosynthesis and chloroplast development at seedling stage at low temperatures. Plant Biology,2019,21(4):585-594.
[22] 周艳虹, 黄黎锋, 喻景权. 持续低温弱光对黄瓜叶片气体交换、叶绿素荧光猝灭和吸收光能分配的影响[J]. 植物生理与分子生物学学
报,2004, 30(2):153-160.
[23] 陈青君, 张福墁, 王永健, 等.黄瓜对低温弱光反应的生理特征研究[J].中国农业科学, 2003,36(1):77-81.
[24] 魏兰波, 董佳慧, 钱稷, 等. 冷驯化期间4种楸树叶片叶绿素荧光参数变化研究[J]. 林业与生态科学,2021,36(1):85-88.
[25] 林植芳, 彭长连, 林桂珠. 光氧化条件下碳代谢中间产物与光合电子传递对PSII光化学活性的调节作用[J]. 热带亚热带植物学报,
2005, 13(1): 1-7.
[26] 林植芳, 彭长连, 林桂珠. 光氧化作用引起几种亚热带木本植物膜损伤和PSⅡ失活[J]. 植物学报, 1999, 41(8): 871-876.
