基于功能类群分析呼兰河口湿地浮游植物群落结构特征
2021-03-31范亚文陆欣鑫
贾 鹏,范亚文,2,陆欣鑫,2,*
1 哈尔滨师范大学生命科学与技术学院, 哈尔滨 150025 2 黑龙江省水生生物多样性研究重点实验室, 哈尔滨 150025
浮游植物是湿地生态系统中的重要初级生产者,也是水生生物食物链中能量流动、物质循环的基础环节[1- 2],其群落结构、物种组成的变化是反映水环境状况的重要指标[3- 4]。浮游植物的时空分布格局不仅可以影响湿地的生态功能和稳定性,而且可以反映湿地生态环境的变化,也是探索湿地生态系统功能的重要手段之一[5- 6]。目前,在湿地生态系统中有关浮游植物的生态研究,主要集中在不同环境梯度和不同生活类型下浮游植物群落结构的差异性分析[7- 9]。这类研究多使用传统的浮游植物集群方法,不能有效地反映浮游植物在湿地生态系统中的功能。Reynolds[10]从个体环境适应性、生态位竞争机制角度出发,将具有相同环境适应策略的浮游植物种类划分为同一个类群,形成了具有相同生态适应特征的分组,即“功能类群”。随着功能类群研究的不断深入,Salmaso[11]等通过植物功能型和FG功能类群的优点提出了MFG划分法;Kruk[12]等根据浮游植物的形态特征,提出了MBFG划分法,这种划分方法大大简化了功能类群的应用。目前,浮游植物功能类群方法已广泛应用在湖泊[13]、河流[14]、水库[15]及湿地[16]等水域生态系统中。
湿地生态系统是水陆两种生态系统的过渡区域,与湖泊、河流等水体相比,水体流动性和营养物质更稳定,具有防控洪水、调节径流等功能[17]。近年来,由于人为活动干扰,导致湿地生态环境不断恶化、生物多样性降低和生态功能退化[18]。呼兰河口湿地保护区位于哈尔滨市呼兰区南部,松花江北岸,呼兰河河口,东西长63.5km,南北宽21.3km,总面积为192.6 km2[19]。该湿地具有调节周边小气候、蓄洪调水和保护物种多样性等生态功能[20]。随着对湿地生态系统保护研究的不断深入,目前已对呼兰河口湿地高等动物[19]、高等植物[20]、水生生物[16]和浮游植物群落结构[21- 22]开展了相关生态研究,以往的研究结果表明该湿地的浮游植物群落结构表现出明显的空间差异,FG功能类群分布特征对该湿地水环境空间异质性响应明显[22]。
以呼兰河口湿地保护区水域为主体,通过FG、MFG和MBFG三种功能类群对呼兰河口湿地浮游植物群落进行研究。定性、定量的评估浮游植物功能类群的演替模式;基于多元统计分析对驱动不同功能类群演替的环境因子进行筛选与识别;初步分析不同功能类群对于呼兰河口湿地水环境变化的响应效果。为温带湿地生态特征研究提供新的分析手段,为我国温带河口湿地水生生物多样性保护与受损湿地恢复提供理论依据。
1 材料与方法
1.1 采样点设置
呼兰河口湿地自然保护区位于黑龙江省哈尔滨市,北部与双井镇、方台镇相连,南与黄土山乡、巨源镇隔江相望。属于北温带大陆性气候,封冻期较长,气温较低,冬天漫长而寒冷;夏天暑期短,炎热、多雨,年平均温度3.6℃[21]。呼兰河口湿地自然保护区内水源地主要为两处:一是呼兰河,发源于小兴安岭西侧、自东向西流动的中型河流;二是松花江,湿地核心区域则是由呼兰河进入松花江后形成[23]。
本研究于2018年春季(4、5月)、夏季(6、7、8月)和秋季(9、10月)三个季节对呼兰河口湿地进行样品的采集,根据呼兰河口湿地的生境特点共设置11个采样点(图1),各采样点的海拔在110—130m之间。根据其生态特征,将11个采样点划分为四个生态功能区域,其中S1、S3、S11为入水口区域,受人为活动影响较大;S5、S7、S10为出水口区域,属于松花江流域;S4、S6、S8、S9为湿地保护区域,保护区正在治理与恢复;S2为农田区域,受农田耕种影响较大。利用GPS全球卫星定位系统收集采样点经纬度信息,采样点地图的绘制通过ArcMap 10.2完成。

图1 呼兰河口湿地采样点布设示意图Fig.1 Map and sampling sites in Hulanhe Wetland样点S1、S3、S11为入水口区域;样点S2为农田区域;样点S5、S7、S10为出水口区域;S4、S6、S8、S9为湿地保护区域
1.2 样品采集、处理及鉴定
分别于2018年4—10月末的上午对11个采样点进行定量样品与定性样品的采集。定性样品采用25号浮游生物网在水面表层下0.5m处作“∞”字形缓慢捞取;定量样品用1L的采水器在水表层以下0.5m处进行水样的采集,现场加入1.5%的鲁格氏固定液15mL进行固定,带回实验室沉淀24h后,浓缩至50mL,用光学显微镜进行镜检计数[24]。浮游植物的鉴定、功能类群的划分、生态分布信息主要参考胡鸿钧、魏印心、胡韧等文献[25- 28]。将至少在一个采样点相对丰度超过15%的功能类群定义为代表性功能类群。
1.3 理化指标的测定
采集水质样品与采集浮游植物样品同步进行,采用Hydrolab多参数水质分析仪现场测定水温(WT)、pH、电导率(SpCond.)、溶解氧(DO)、氧化还原电位(ORP)、浊度(Tur.)以及叶绿素a(Chl-a)。现场采集的水样用棕色玻璃瓶避光保存,带回实验室24h内依据《水和废水监测方法》(国家环境保护总局,2002)进行总氮(TN)、总磷(TP)、高锰酸盐指数(CODMn)、五日生化需氧量(BOD5)[29]的测量。
1.4 数据分析
使用PRIMER 5.0软件进行ANOSIM和SIMPER分析。独立样本T检验通过SPSS 20.0软件完成。去趋势分析(DCA)和冗余分析(RDA)采用软件CANOCO 4.5进行,在进行排序分析前,对解释变量和响应变量数据进行log(x+1)转换,使样本总体趋于正态分布。通过蒙特卡洛置换检验,判断解释变量对响应变量生态分布影响的显著性,建立二维排序图。使用Origin 2018与Excel 2018软件对图表进行绘制。
2 结果与分析
2.1 水体理化因子变化
独立样本t检验表明在不同季节之间水体理化因子变化明显,除TP和SpCond.外其他理化指标在季节之间均存在一定的变化规律(P<0.05)(表1)。DO、pH和Tur.都呈现随春、夏、秋季节变化逐渐升高的规律(P<0.05),三季平均值分别为2.66、8.73、41.48mg/L,变化范围分别在0.83—5.34mg/L、7.28—10.04和21.93—97.60mg/L之间;夏季的WT高于春、秋两季(P<0.05),三季平均值为19.45℃,变化范围在9.89—31.23℃之间;CODMn呈现出随着春、夏、秋季节变化逐渐降低的规律(P<0.01),三季平均值为5.49mg/L,变化范围在0.82—10.50mg/L之间;SpCond.和ORP呈现出春季最高,夏季最低的规律,三季平均值分别为402.57mS/cm和414.91mV,变化范围在161.13—1180.00mS/cm和340.00—511.00mV之间。
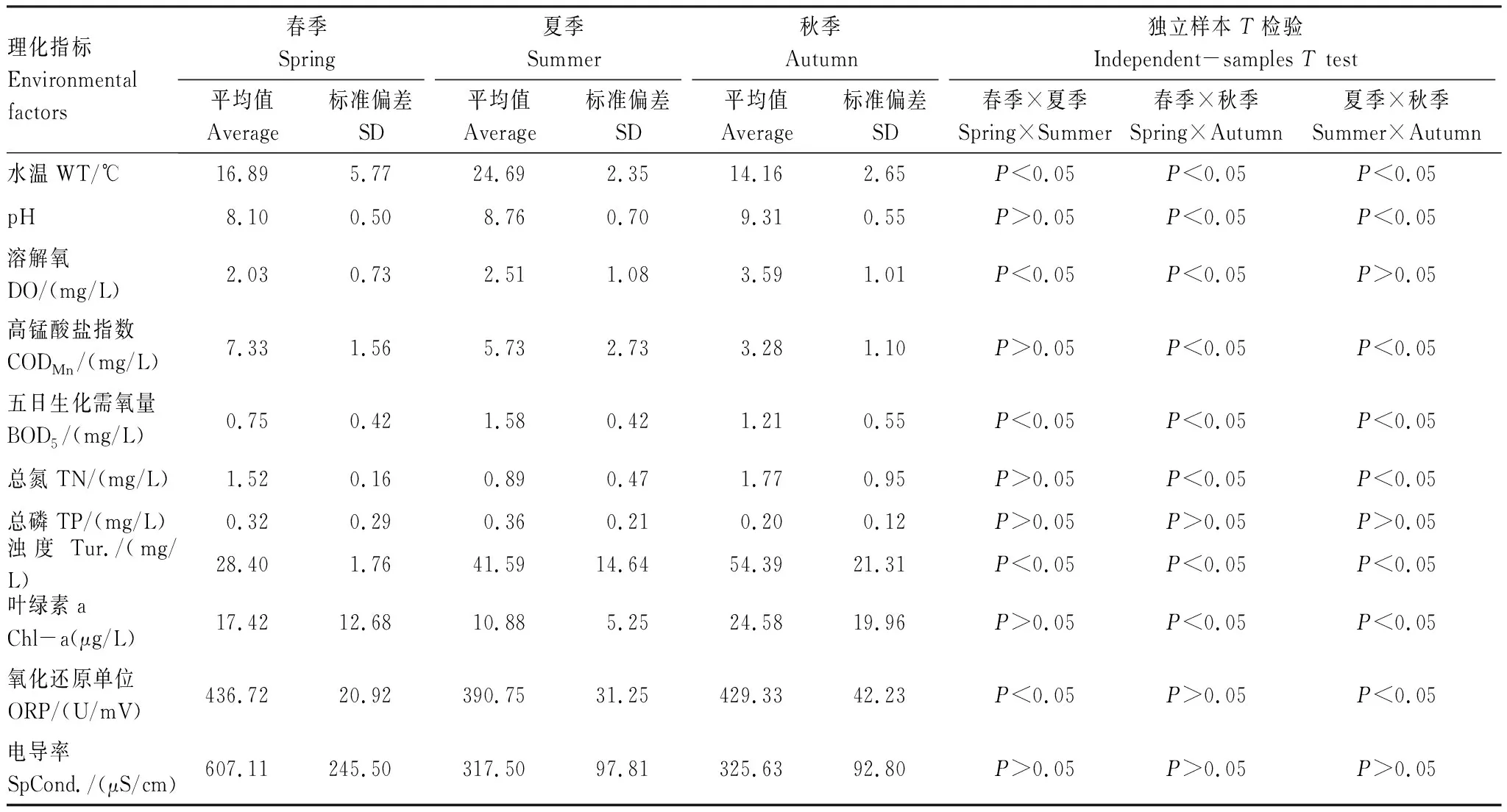
表1 呼兰河口湿地水体理化因子季节变化
2.2 呼兰河口湿地浮游植物种类组成
在研究期间共鉴定浮游植物243个分类单位,包括10变种,隶属于7门9纲18目32科75属。绿藻门(Chlorophyta)种类最多,为104种及8变种,占浮游植物种类总数的46.09%;裸藻门(Euglenophyta)种类46种及1变种,占19.34%;硅藻门(Bacillariophyta)种类44种及1变种,占18.52%;蓝藻门(Cyanophyta)种类27种,占11.11%;甲藻门(Dinophyta)种类7种,占2.88%;隐藻门(Cryptophyta)种类3种,占1.23%;金藻门(Chrysophyta)种类仅2种,占0.82%。在空间尺度上,呈现湿地保护区域(188种)>入水口区域(183种)>出水口区域(172种)>农田区域(135种)的规律;在时间尺度上,呈现夏季(198种)>秋季(152种)>春季(143种)的规律。
2.3 呼兰河口湿地浮游植物丰度的时空变化
浮游植物丰度在时间尺度上表现为夏季平均值最高,变化范围在(0.67—26.26)×106个/L之间;其次为秋季,变化范围在(0.39—20.15)×106个/L之间;春季最低,变化范围在(0.50—27.78)×106个/L之间。
浮游植物平均丰度呈现出湿地保护区域>入水口区域>农田区域>出水口区域的规律。湿地保护区域内的浮游植物平均丰度最高,变化范围在(15.92—95.44)×106个/L之间;入水口区域浮游植物丰度次之,变化范围在(4.47—25.40)×106个/L之间;农田区域浮游植物丰度变化范围在(1.28—27.78)×106个/L之间;出水口区域浮游植物丰度最低,变化范围在(2.44—11.33)×106个/L之间(图2)。

图2 呼兰河口湿地浮游植物丰度分布特征 Fig.2 Phytoplankton abundance spatial and temporal distribution of Hulanhe Wetland
2.3 FG、MFG和MBFG功能类群演替
根据Reynold和Padisák等提出的FG功能类群划分方法,共划分FG功能类群25个,分别为B/C/D/E/F/G/H1/J/LM/Lo/M/MP/N/P/S1/S2/SN/T/TB/W1/W2/X1/X2/X3/Y。其中代表性功能类群为B/D/F/J/Lo/MP/P/S1/W1/W2/X1/Y(表2,表3)。出现频率超过50%的功能类群为B/D/F/H1/J/Lo/MP/P/S1/W1/W2/X1/X3/Y,出现频率在30%—50%的功能类群为E/G/LM/X2,出现频率小于30%的功能类群为C/M/N/T/TB/S2/SN(图3)。FG功能类群季节演替特征为春季B(梅尼小环藻CyclotellameneghinianaKützing)/D(谷皮菱形藻Nitzschiapalea(Kützing) W.Smith)/Y(啮蚀隐藻CryptomonaserosaHering)→夏季B/D/S1(湖泊假鱼腥藻Pseudanabaenalimnetica(Lemmermann) Komárek)→秋季B/X1(狭形纤维藻AnkistrodesmusangustusBernard)(图4)。
根据Salmaso等对功能类群的划分[11],本次研究中呼兰河口湿地共划分MFG功能类群20个,分别为1b/1c/2c/2d/3a/3b/5a/5d/5e/6a/6b/6c/7a/8a/9a/9b/10a/11a/11b/11c。代表性功能类群为1c/2c/2d/5a/6a/6b/7a/9b/10a/11a/11b(表3)。出现频率大于50%的功能类群为1b/1c/2c/2d/5a/5d/5e/6a/6b/7a/8a/9b/10a/11a/11b,出现频率大于30%且小于50%的功能类群为11c/3b/6c/9a,功能类群3a出现频率小于30%(图3)。MFG功能类群季节演替特征为春季2d(隐藻门小个体)/7a(中心纲小个体)/6b(羽纹纲大个体)→夏季5a(颤藻目丝状体)/6b/7a/→秋季7a/10a(绿藻纲丝状体)(图4)。
基于Kruk等研究结果[12],在呼兰河口湿地共划分MBFG功能类群6个,分别为Ⅰ/Ⅲ/Ⅳ/Ⅴ/Ⅵ/Ⅶ(表2,表3)。研究期间6种功能类群均出现,其中Ⅴ类群在三个季节中都占据主要地位,出现频率100%,Ⅳ和Ⅵ出现频率超过98%,Ⅲ功能类群出现频率最低但也超过了50%以上(图3)。MBFG功能类群季节演替特征为春季Ⅵ(具有硅质外壁且无鞭毛的种类)/Ⅴ(具鞭毛中到大型的单细胞种类)→Ⅵ/Ⅳ(不具有明显特征的中等大小种类)→Ⅵ/Ⅳ(图4)。

表2 呼兰河湿地浮游植物功能群ANOSIM分析及SIMPER分析

表3 呼兰河口湿地三种浮游植物功能类群的代表性属种
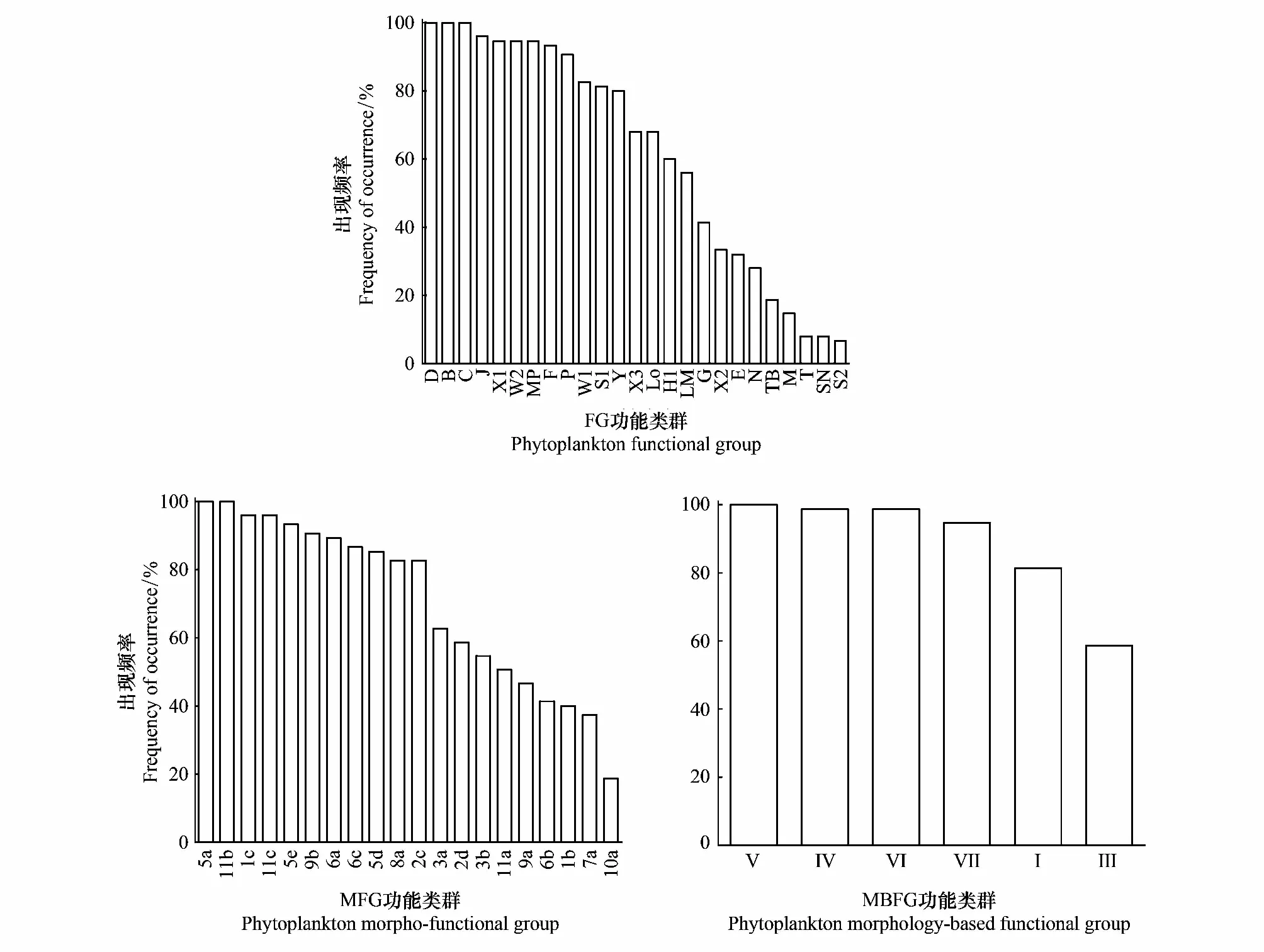
图3 呼兰河口湿地浮游植物功能类群出现频率Fig.3 Frequency distribution of functional groups in Hulanhe Wetland

图4 呼兰河口湿地三种功能类群丰度相对百分比Fig.4 Relative percentage of abundance of three functional groups in Hulanhe Wetland
基于FG、MFG和MBFG功能类群群落相似性分析(ANOSIM),表明季节变化对浮游植物群落演替驱动作用明显(P<0.05)。SIMPER分析表明,B/SI/Y/W2/X1/D/Lo、7a/5a/10a/2d/6b/2c/11a和Ⅵ/Ⅳ类群,分别为不同季节之间FG、MFG和MBFG类群演替的主要贡献者。
2.4 多元统计分析
对呼兰河口湿地的11种环境变量(WT、pH、SpCond.、DO、ORP、Tur.、Chl-a、TP、TN、CODMn与BOD5)与FG、MFG和MBFG功能类群进行排序分析,不同功能类群对环境变化的响应模式进行识别。三种功能类群DCA分析显示最长长度梯度分别为3.003和0.920和1.222,因此选择线性模型RDA对优势功能类群与环境变量进行排序分析。经蒙特卡洛置换检验表明,呼兰河口湿地三种功能类群的生态分布受多种环境因子共同作用影响,驱动功能类群生态分布的环境因子为:CODMn、WT、TP、SpCond.、pH、BOD5、Tur.、ORP和DO。

图5 呼兰河湿地植物功能类群与环境变量的RDA分析Fig.5 RDA of representative functional groups and environmental variables in Hulanhe wetland
3 讨论
3.1 呼兰河口湿地浮游植物功能类群演替特征
浮游植物功能类群方法从个体生态学角度出发,基于物种生态适应性角度分析种群的生态选择机制[30]。不同功能类群内的浮游植物种类具有其自身独特的生态特征,其演替通常与水体营养状态、光照及水温密切相关[31],在“上行效应”和“下行效应”两种机制调控下,当栖息地水环境发生变化时,浮游植物自身的形态、生理适应特征会主动或被动的适应环境特征的改变,以演替的形式来维持湿地的生态平衡[32]。
呼兰河口湿地受温带季风气候影响,季节之间水体理化环境特征变化明显[14]。FG功能类群将物种的生态属性划分到功能类群当中,其演替模式被认为是响应水环境变化的有效指标[18]。在本研究中FG功能类群季节演替特征为,春季B/D/Y类群演替为夏季B/D/S1类群,最后被秋季的B/X1类群所取代。B类群是硅藻门中的小环藻属,为α-ms(中营养)指示种,其生活环境适宜在温度20℃左右[33- 34]、高营养盐的中小型较浅的水体中。水体中营养盐含量的升高是导致富营养化的根本原因,其变化会直接影响浮游植物的丰度。在高营养盐的水体中,浮游植物的丰度通常会受光照的限制,光的可利用性也会影响浮游植物的竞争和群落多样性[35],因此耐受低光环境的类群通常会占据优势地位。呼兰河口湿地在春季期间,水温较低,日照时间相对较少,处于营养盐积累阶段,低水温的条件会限制大部分浮游植物的生长,与此同时B类群具有高捕光效率和高相对表面积比的特点,因此在栖息地的适宜性及较低光照的基础上,B类群在群落中占据优势地位[36]。Y类群的代表性物种为卵形隐藻和啮蚀隐藻,适宜生活在磷浓度较高的水体中,对低光耐受,为广适应性种类[32]。由于春季湿地保护区域人为干扰较少,水体流动性差,磷浓度较高,同时Y类群物种具有鞭毛,有较强的运动性,使其能在水体中垂直运动,可以更好地躲避捕食者,进而在资源竞争中占据有利地位[35]。因此,B和Y类群能在春季占据优势。D类群适应于营养盐含量较高的浅水水体、耐受于低光条件。呼兰河口湿地夏季水体温度升高,溶解氧含量和pH(平均值pH=8.76)增加,水体呈碱性。在一定范围内,温度和溶解氧的升高会促进B/D类群的生长[37]。同时,水体的弱碱性为D类群提供了良好的生活条件,因此D类群能在夏季占据优势地位[38]。呼兰河口湿地夏季降雨充沛,周边土地中的营 养物质通过淋溶作用进入水体,入水口区域、农田区域和湿地保护区域具有较多的人为活动干扰,因此导致水体变浑浊(浊度平均值为41.59 mg/L),限制光照摄入,进而影响耐受低光环境的B/D类群生长。营养盐是水体中重要的生源要素,由于呼兰河口湿地水体氮磷比小于16∶1,水体受氮限制明显。低氮的水体限制了大部分浮游植物的生长,而蓝藻的生长与水体的氮磷比密切相关[39],S1类群中的丝状蓝藻具有固氮能力,因此营养盐水平降低可能是S1类群中的丝状蓝藻(中富营养)占据优势的重要原因。本文SIMPER分析也证明S1类群的变化是影响不同季节之间FG功能类群差异的主要功能类群之一。从生物因素的角度看,“上行效应”和“下行效应”对不同营养类型的水体作用是不一样的,这是由浮游植物的粒径大小导致的[40]。S1类群中的丝状蓝藻可抵御浮游动物的捕食,在低牧食压力的情况下容易占据优势,因此S1类群代替了春季广适应性的Y类群,在夏季浑浊且温暖的水体中与B/D类群共同占据主要优势地位。呼兰河口湿地夏、秋两季之间存在总氮含量明显升高的过程(P<0.05),X1类群的代表性种为狭形纤维藻,为α-ms(富营养)指示种,能够耐受低温环境,适宜在含氮量高的水体中生长[35]。因此替代了夏季的蓝藻门S1类群。陆欣鑫等[22]对呼兰河口湿地的研究也证明了X1类群在秋季占据优势地位。
MFG功能类群反映了浮游植物运动能力、大小、形状、营养物质喜好、营养盐和碳的获取能力等[40]。本次研究中,MFG功能类群季节性演替特征为由春季的2d/7a/6b类群演替为夏季的5a/7a/6b类群,最后由秋季的10a/7a类群所替代。MFG功能类群的季节演替特征所反映的水环境变化过程与FG功能类群差异不大。MBFG功能类群方法是利用浮游植物对水环境变化特征的响应为标准,反映某一特定生境模块的状况。ANOSIM分析表明,以MBFG功能类群为研究手段分析季节之间群落结构差异效果不明显。季节之间SIMPER分析也表明,在不同季节之间影响群落结构差异的类群无变化。其原因可能是Ⅴ类群(具鞭毛中到大型的单细胞种类)和Ⅵ类群(具有硅质外壁且无鞭毛的种类)为春季的主要优势功能类群,两类群主要为隐藻和硅藻。春季低温且低光照,具有鞭毛的隐藻根据自身的形态特点以及对低温低光的耐受性,使其能在春季具有很强的竞争能力,与Ⅵ类群的硅藻在春季共同占据优势地位。夏季和秋季主要为Ⅵ类群的硅藻种类和Ⅳ类群中不具有明显特征的中等大小种类,Ⅳ类群种类繁多,包含大部分除硅藻以外的其他浮游植物种类,此类群具有广适应性的特点,因此在夏、秋两季占据优势地位。由于呼兰河口湿地夏、秋两季Ⅳ/Ⅵ类群对资源具有很强的竞争力,导致Ⅴ类群在水体中的生长被限制,这可能是Ⅴ类群在夏、秋两季不占据优势的原因之一。
对比FG、MFG和MBFG三种功能类群的演替模式不难发现,FG与MFG类群在描述呼兰河口湿地浮游植物演替特征上有重叠部分,差异较小;MBFG与其他两种功能类群相比,在表征季节间水环境的变化过程中效果并不理想。应用功能类群方法对温带典型水库中的研究曾指出,MBFG功能类群分类方法在水生态研究中虽然具有简单、高效的优势,但对环境的解释度不够[41]。FG功能类群将浮游植物的定量和定性数据与多种水环境数据相结合,很大程度上提高了浮游植物对环境的响应能力,将包含多种环境因子的复杂生态环境与浮游植物的耐受性和敏感性相关联,进而说明浮游植物与环境因子之间的相关性,同时预测研究水体的营养状态。虽然MFG弱化了对生态环境的考虑,导致对环境的解释不如FG功能类群清晰,但仍然能够揭示呼兰河口湿地浮游植物群落季节变化特征。MBFG在响应温带湿地季节间环境变化梯度的分析中,效果没有FG和MFG功能类群理想。
3.2 影响呼兰河口湿地浮游植物功能类群演替的环境因子
大量研究表明,温带湖泊、水库及湿地水体中浮游植物群落的演替受浮游动物、滤食性鱼类通过下行效应影响,另一方面,水温、光照、营养盐及资源可利用率等上行效应更是驱动群落演替的主要原因[39]。通过对中国3个典型湖泊浮游植物研究发现,浮游植物功能类群对不同营养类型的湖泊响应明显[42]。基于MBFG功能类群对亚马逊河流域浮游植物群落特征研究,得出MBFG类群能有效的指示水体电导率和pH分布特征[43]。呼兰河口湿地是中型恢复性湿地,其入水口区域与农田区域受季节性的人为活动干扰较大。浮游植物功能类群对湿地栖息环境的时空异质性响应存在独特模式,优势类群的支配地位会随着空间异质性的改变而被其他类群所替代。营养盐、pH、水温、浊度和水动力特征等都是影响浮游植物出现或者消失的重要因素,同时影响各种功能类群的组成。RDA分析得出三种功能类群的演替是由多种环境因子共同作用的结果。经过蒙特卡洛置换检验分析表明,pH、Tur.、BOD5和CODMn是影响呼兰河口湿地浮游植物功能类群演替的最主要因子。
CODMn为指示浮游植物栖息环境有机污染的重要指标之一,其数值越高说明水体富营养程度越高[44]。CODMn的变化往往会影响浮游植物的演替[45]。在研究期间,呼兰河口湿地CODMn值在(0.82—10.50)mg/L之间变化,三种功能类群在三个季节间CODMn差异性都呈显著水平(P<0.01)。RDA排序图显示,FG功能类群中的Y/D/W2类群、MFG功能类群中的2d/2c类群和MBFG功能类群中的Ⅴ类群都与CODMn值变化密切相关,并都呈现出随着CODMn值升高各类群丰度上升的趋势,这些类群主要在春季和夏季中具有较高的权重。春季研究区域水体较浅且浑浊,营养物质积累,由于春季农田区域与入水口区域人为活动干扰,造成水体中有机污染物增多,因此春季的CODMn值最高,平均值为7.33mg/L。呼兰河口湿地春季较高的CODMn值也伴随着相对较高的磷含量,Y/D/2d/Ⅴ类群对水体中的磷浓度需求较大,因此在春季水体中高磷浓度的条件下,促进了这些类群的生长。同时春季水温和光照较低,耐受低光和低温的B/D/7a/6b/Ⅵ类群在浮游植物群落中也占据优势地位。在一定程度上,CODMn值变化所反映出的水体有机物质含量的改变也影响了FG功能类群的B/D/Y/S1/X1类群的生长与演替,此结果与同气候带的另一湿地(金河湾湿地)的研究结果近乎一致[16]。BOD5与CODMn共同反映水体中的有机物含量,RDA排序分析表明,S1/X1/5a/10a/Ⅶ类群的分布与BOD5含量密切相关,随着BOD5的升高,其种类丰度逐渐升高。BOD5值在夏、秋两季较高,平均值分别为1.58mg/L和1.21mg/L,水体呈富营养状态,因此S1/X1/5a/10a类群较适应夏、秋两季的水体环境。Ⅶ类群体内可贮存营养盐,多为具有外胶被和较小表面积比的绿球藻目、颤藻目的群体,属于冗余耐受生长策略[26],通过形成较大群体,以抵御外部的捕食压力。同时BOD5与Chl-a也密切相关,在适当的范围内,有机物质会促进浮游植物的代谢[39],浮游植物通过自身光合作用会产生大量的有机物释放到水体中,使水体中的BOD5进一步升高,这一结论在对巢湖相关的功能类群研究中也得到证明[46]。可见,BOD5所反映的水体中有机物含量的变化,对湿地中S1/X1/5a/10a/Ⅶ类群的生长、演替和分布都有着明显的影响。
pH与浮游植物的分布关系密切,特别是与一些硅藻种类[47]。相关研究表明,pH变化可以通过影响胞内部分酶活性或跨膜质子梯度来影响浮游植物的光合作用过程,进而影响浮游植物的生长[48]。RDA排序表明,FG功能类群的D/Y类群、MFG功能类群的6b类群和MBFG功能类群的Ⅴ类群均与pH呈负相关。浮游植物的光合及呼吸作用日变化研究表明,通常情况下浮游植物夜间进行呼吸作用会释放大量的CO2,造成水体pH降低,本研究样品采集时间多为清晨至上午,这也是pH与浮游植物丰度呈负相关的原因之一[42];MBFG功能类群的Ⅳ/Ⅵ类群与pH变化密切相关,浮游植物通过光合作用利用水中无机碳(H2CO3),产生氢氧根离子,导致水体pH升高,随着其上升,导致水体pH与浮游植物丰度呈正相关,因此水体中的pH适度的升高会影响Ⅳ/Ⅵ类群浮游植物的生长[49]。浊度是由有机与无机漂浮物、微生物和尘土等微小颗粒悬浮物质所共同引起的[40],与光照强度、水体理化性质以及气候条件具有密切的关系,水体的浊度直接会影响浮游植物的丰度。RDA排序分析表明,Y/6b/Ⅴ类群随着浊度的升高表现出丰度下降的趋势。由于夏、秋两季降雨充沛,营养盐通过淋溶作用进入水体,与此同时水体扰动较大,导致水体的浊度逐渐升高[39],较高的浊度会影响浮游植物的光合作用,使浮游植物产生的有机物质减少,由于Y/6b/Ⅴ类群的个体相对较大,在相对面积的光照射下,这些类群的浮游植物光合作用更弱,从而导致Y/6b/Ⅴ类群在夏、秋两季高浊度的水体中处于不利地位。B/10a/Ⅵ类群丰度的变化也与浊度密切相关,由于浊度的增加会使水体的透明度下降,进而限制了光照的射入,由于B类群的梅尼小环藻以及10a/Ⅵ类群的部分个体相对较小,在相同光照条件下,这些类群的光合作用更强,因此10a/Ⅵ类群更具有竞争优势。因此B/10a/Ⅵ类群在浊度升高的条件也会占据优势地位。呼兰河口湿地是正在恢复中的湿地,本次研究与近8年前研究相比[21- 22],虽然优势种略有改变,但水生植物整体群落结构较为稳定。综合来看,呼兰河口湿地浮游植物群落演替受多种环境因子共同影响,其中pH、Tur.、BOD5和CODMn与三种浮游植物功能类群演替密切关系。本研究通过分析不同功能类群分布特征及环境相关性,以期进一步完善湿地水生态系统的评价体系,这对呼兰河口湿地水生生物多样性保护与生态恢复具有一定的理论与指导意义。
4 结论
(1)呼兰河口湿地在调查期间共鉴定浮游植物7门243个分类单位。共划分FG功能类群25个,MFG功能类群20个,MBFG功能类群6个。
(2)相较于MBFG功能类群,FG和MFG功能类群能更好的响应呼兰河口湿地水环境的空间异质性。
(3)CODMn和BOD5、pH、Tur.是驱动呼兰河口湿地浮游植物群落结构空间变化的主要环境因子。
