福建主栽非洲菊品种遗传多样性ISSR分析
2021-03-28夏朝水江斌邓永生陈玮婷曹奕鸯
夏朝水 江斌 邓永生 陈玮婷 曹奕鸯


摘 要:利用ISSR技术对35份福建主栽非洲菊资源进行遗传多样性分析。从100条ISSR引物中筛选出多态性好的15条随机引物,分别对35份非洲菊资源基因组DNA进行扩增,共扩增得到78条清晰可辨条带,其中多态性条带70条,多态位点百分比达89.7%。利用DPS软件统计分析,结果表明,各品种间的遗传相似系数介于0.2968~0.7500,当阈值为0.4200时,可将35个非洲菊分成4类。当遗传相似系数为0.4914时,第I类非洲菊又可分为4个亚类。聚类结果表明,花心颜色和重瓣类型可以作为非洲菊亲缘关系分类的一种依据,同一搜集来源的品种有聚类到一起的趋势,这与农艺性状的聚类结果有一定的对应关系。
关键词:非洲菊;种质资源;简单重复序列间扩增;遗传多样性
中图分类号:S 682.1 文献标志码:A 文章编号:0253-2301(2021)01-0009-06
DOI:10.13651/j.cnki.fjnykj.2021.01.002
Abstract:Genetic diversity of 35 major cultivars of Gerbera jamesonii Bolus in Fujian was analyzed by ISSR technology. 15 random primers with good polymorphism were selected from 100 ISSR primers, and were amplified on the genomic DNA of 35 Gerbera jamesonii Bolus. A total of 78 clear and distinguishable bands were obtained, of which 70 were of polymorphism, and the percentage of polymorphic sites reached 89.7%. The DPS software was used for the statistical analysis, and the results showed that the genetic similarity coefficients among the varieties ranged from 0.2968 to 0.7500. When the threshold value was 0.4200, the 35 Gerbera jamesonii Bolus could be divided into four categories. When the genetic similarity coefficient was 0.4914, the Class I Gerbera jamesonii Bolus could be further divided into four subclasses. The clustering results showed that the color of flower heart and the type of double petals could be used as a basis for the classification of genetic relationship of Gerbera jamesonii Bolus, and the varieties from the same source tended to cluster together, which had a certain corresponding relationship with the clustering results of agronomic traits.
Key words:Gerbera jamesonii Bolus; Germplasm resources; ISSR(inter-simple sequence repeat); Genetic diversity
非洲菊Gerbera jamesonii Bolus,又名扶郎花、太陽花,为菊科大丁草属多年生宿根草本花卉,是世界五大鲜切花之一,在国内外市场上非常畅销,备受消费者的青睐[1-3]。福建主栽非洲菊品种大多从外地引进,经常由于对品种习性不甚了解,或者品种已经开始退化,盲目引种导致巨大的经济损失。因此,采取各种育种技术方式,及早培育出具有自主知识产权新品种(系)非常必要。为培育超亲性状的非洲菊新品种,需分析亲本资源的亲缘关系才能更好地配置杂交组合,而分子标记目前广泛用于种质资源亲缘关系及遗传多样性分析,因此,利用分子标记辅助育种技术分析非洲菊品种资源之间的亲缘关系可为非洲菊遗传改良提供更快速、可靠、稳定的辅助选择手段。
近年来,部分学者采用分子标记技术对非洲菊进行初步研究。李宗菊等[4]以改良的CTAB法为基础,在第2次用氯仿/异戊醇抽提时,加入‘PCN溶液,可明显去除多糖;在材料研磨时加入配制的化学物质,可以明显提高DNA纯度及ISSRPCR扩增质量;利用该法所提取的DNA稀释30倍后,进行ISSRPCR扩增可得到清晰谱带。吴莉英等[5]以非洲菊的嫩叶、老叶为材料,利用改进的CTABFREE洗涤法提取纯度、浓度都符合AFLP分析要求的DNA,获得非洲菊清晰AFLP指纹图谱。李达等[6]对非洲菊SRAPPCR反应体系的影响因子进行探讨,得到了最佳的SPARPCR反应体系。此外,张伟等[7]、张玉进等[8]分别改进了非洲菊不同组织总RNA的提取方法。沈阳农业大学的聂京涛等[9]对75份非洲菊品种资源进行了遗传多样性的ISSR分析,发现非洲菊具有丰富的遗传多样性;将非洲菊品种的花色、管状花颜色、花径、花型结合聚类结果,发现其存在一定的相关性,这为非洲菊的良种选育在分子水平上奠定了理论基础。福建省农业科学院钟海丰等[10]设计100条ISSR引物,对外引的22个非洲菊品种进行遗传多样性的ISSR分析,表明非洲菊品种间具有较丰富的遗传多样性,花瓣及花心颜色可以作为区分非洲菊品种分类及分析亲缘关系的初步依据。因此,本研究利用ISSR分子标记辅助育种技术分析福建主栽非洲菊品种资源之间的亲缘关系,为新品种的选育提供依据。
1 材料与方法
1.1 试验材料
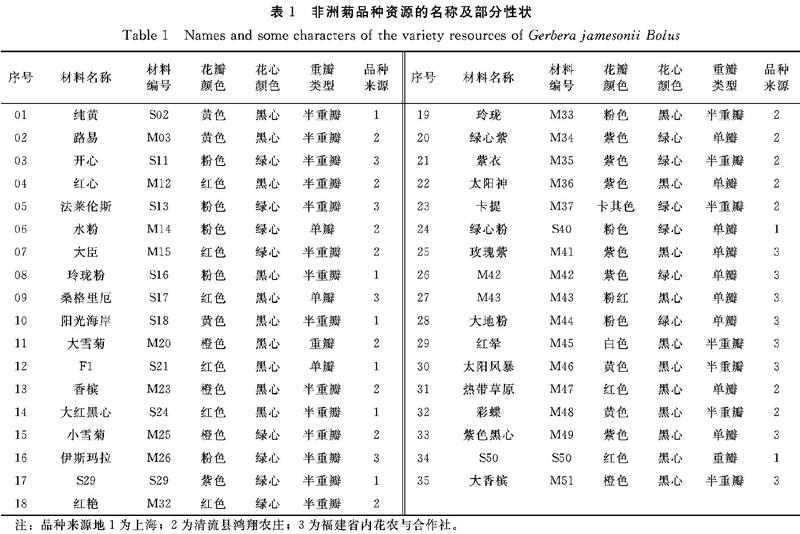
供试材料为三明市农业科学研究院收集的福建省内主栽35个非洲菊品种(表1)。其中8份材料引自上海,15份材料引自清流县鸿翔农庄,12份材料引自福建省内花农与合作社。试验每个品种随机选取3株采集,每株采1片嫩叶,装于自封袋,保存于-4℃冰箱中备用。
1.2 试验方法
1.2.1 基因组DNA的提取 非洲菊基因组DNA的提取采用CTAB法,方法参考《植物遗传手册》植物基因组总DNA提取方法[11],试验所提取的基因组DNA用1%琼脂糖凝胶电泳检测,浓度采用紫外分光光度计法进行测定,基因组DNA于-20℃保存备用。
1.2.2 引物篩选与PCR扩增 ISSR引物的筛选是以田间植物性状表现差异较大且DNA质量较高的1号与12号材料对从上海生物工程有限公司提供的100条ISSR随机引物中共筛选稳定性好、条带清晰、多态性好的引物用于非洲菊遗传多态性试验[12]。
ISSRPCR反应体系参考沈颖等[13]和聂京涛等[9]步骤稍做修改,15 μL ISSR反应体系中包含2 μL 10×PCR Buffer(Mg2+ Plus)、1 μL dNTPs、0.15 μL Taq酶(5 U·μL-1)、1 μL模板DNA(20 ng·μL-1)、1 μL引物、9.85 μLddH2O。PCR反应在Master Cyclergradient 96梯度PCR仪(Eppendorf)上进行,反应程序为:94℃预变性4 min;94℃变性45 s,52℃退火45 s,72℃延伸2 min,共40个循环;72℃延伸5 min ,于4℃保存PCR产物。扩增产物用1%的琼脂糖凝胶电泳检测,凝胶板置于5% EB(溴化乙锭)溶液中染色后,在BIORAD公司的GelDoc 1000型凝胶成像分析系统上进行数据采集、分析处理、保存及结果输出。
1.3 统计分析
对35份材料获得的DNA条带进行统计,每条DNA条带记为一个单位,在相同迁移位点无条带的赋值为0,有条带的赋值为1,得到的数据用Excel录入电脑中。数据分析采用DPS统计软件,然后用类平均聚类法(UPGMA)对其进行聚类分析,生成聚类图。
2 结果与分析
2.1 ISSR随机引物的确定
以试验选出田间试验性状表现差异较大且DNA质量较高的1号与12号材料对100条ISSR引物进行筛选,最终确定15条多态性高、条带清晰的引物用于分析材料的多态性。其引物名称与序列见表2,用1号与12号材料筛选出的15条引物PCR扩增带型见图1。
2.2 ISSR扩增产物的多态性
试验筛选出15条多态性高的引物用于35份非洲菊材料的遗传多样性研究,PCR共扩增出78条清晰条带,其中多态性条带70条,多态位点百分比为89.7%,平均每条引物可扩增4.7个多态性条带(表3)。图2~3为引物I34和I55对所有非洲菊品种的电泳图谱。引物扩增出多态性条带最多的为7条,最少的为3条。15个引物中有1个是四核苷酸重复序列,2个是三核苷酸重复序列,其余12个引物都为二核苷酸重复序列,表明非洲菊基因组以二核苷酸重复居多。
2.3 聚类分析和亲缘关系
采取DPS软件进行数据统计,用UPGMA法对其进行聚类分析,构建亲缘关系树状图(图4),结果显示,35份非洲菊品种资源之间的遗传相似系数为0.2968~0.7500,其中阳光海岸和大雪菊相似系数最大,为0.7500,表明它们之间的遗传关系最近,其花色分别为黄色黑色与橙色黑色;红晕和太阳风暴的相似系数最小,为0.2968,表明它们之间的亲缘关系最远,花色花心分别为白色黑心与黄色黑心。
以遗传相似系数0.42为阈值,非洲菊35份资源可分成4类:第I类包含纯黄、阳光海岸、大红黑心、玲珑、热带草原等30份资源;第Ⅱ类有M42和大地粉2份资源,遗传相似系数为0.4894,花色花心分别是紫色绿心和粉色绿心;第Ⅲ类只有玫瑰紫1份资源,其遗传相似系数为0.3095,花色花心为紫色黑心。第Ⅳ类有红晕和太阳风暴2份资源,遗传相似系数为0.2968,花色花心分别为白色黑心与黄色黑心。
当第Ⅰ类以遗传相似系数0.4914为阈值时,又可分为4个亚类,第Ⅰ亚类包含纯黄与绿心粉2个品种。第Ⅱ亚类包含22个品种,分2小类,第1小类包含18个品种,其中阳光海岸和大雪菊的亲缘关系最近,其遗传相似系数为0.7500,花色花心分别为黄色黑心和橙色黑心;伊斯玛拉的遗传相似系数为0.5343,花色花心为粉色绿心。第2小类包含开心、红心、法莱伦斯和香槟4个品种,其中红心和法莱伦斯的亲缘关系最近,遗传相似系数为0.6667,花色花心分别是红色黑心和粉色绿心;香槟的遗传相似系数最小,为0.5437,其为橙色黑心品种。第Ⅲ亚类包括热带草原、彩蝶、S50、紫色黑心和大香槟5个品种,其中紫色黑心和大香槟的亲缘关系最近,遗传相似系数为0.7179,花色花心分别是紫色黑心和橙色黑心;热带草原的相似系数最小,为0.4687,其为红色黑心品种。第Ⅳ亚类只有S29,其遗传相似系数为0.4299,为紫色绿心品种。
3 讨论
在ISSR引物的多态性筛选上,本试验选择15条多态性好的引物共扩增出多态性条带70个,平均每条引物4.7个多态性条带,相比前人研究结果[6],研究扩增出的多态性条带数相对较少,这可能与ISSRPCR反应体系的组成DNA或模板量的浓度有关。试验由材料1号和12号筛选确定的15条多态性高、条带清晰的ISSR引物中,有1个是四核苷酸重复序列,2个是三核苷酸重复序列,其余12个引物都为二核苷酸重复序列,表明非洲菊基因组以二核苷酸重复居多。这与缪恒彬等[14]、聂京涛等[9]的研究结果相同。
在非洲菊遗传聚类亲缘关系上,聂京涛等[9]利用ISSR分子标记技术对非洲菊材料进行聚类,结果表明非洲菊各品种之间的遗传关系与其花色、花型、管状花颜色、花径呈现一定的相关性。钟海丰等[10]利用ISSR分子标记技术对外引的22个非洲菊品种进行遗传多样性及聚类分析,得出花瓣和花心颜色可作为非洲菊品种间的亲缘关系分类的一种初步依据,这些研究都为非洲菊优良品种的选育奠定了一定的分子理论基础。本试验通过对35份非洲菊品种资源聚类分析得出,品种间的遗传相似系数为0.2968~0.7500,表明各个材料的遗传背景和亲缘关系复杂,存在较大的遗传分化,这可能与非洲菊品种存在自交不亲和性及长期的无性繁殖有关[15]。本研究根据ISSR聚类图可将35份非洲菊资源分为4大类,虽然花色、花心颜色、重瓣类型等性状没有完全聚到一块,但花心颜色和重瓣类型却聚集在各个分支上,且分支上的相似度很高,说明花心颜色和重瓣类型可以作为非洲菊亲缘关系分类的一种依据;特别值得注意的是,不同搜集来源的品种有聚集到一起的趋势,其中从清流鸿翔农庄搜集的15个品种绝大部分聚集到第Ⅱ亚类,而从农户或合作社收集的品种却分散到各个类别中,这或许可能是因为农户和合作社购买种苗的来源地不同,有云南、广东、上海等地的种苗,这就导致了其亲缘关系可能存在较大差别;而这些与农艺性状的聚类结果有一定的对应关系。
本试验通过ISSR分子标记结合农艺性状的聚类研究,发现非洲菊品种在农艺性状方面差异较大,且遗传多样性丰富,在分子水平上形成较大的遗传变异,这可能是由于非洲菊品种间存在自交不亲和性以及长期的无性繁殖有关。本研究通过了解非洲菊各品种的遗传和亲缘关系,为非洲菊杂交亲本的选配与良种选育提供技术支持,从而加速育种进程。
参考文献:
[1]杨先芬.工厂化花卉生产[M].北京:中国农业出版社,2002:160-162.
[2]王意成.最新图解草本花卉栽培指南[M].南京:江苏科学技术出版社,2007:144-145.
[3]程用谦.中国植物志(第79卷)[M].北京:科学出版社,1996:30-47.
[4]李宗菊,熊丽,桂敏,等.非洲菊基因组DNA提取及ISSRPCR扩增模板浓度优化[J].云南植物研究,2004,26(4):439-444.
[5]吴莉英,唐前瑞,李达,等.非洲菊的AFLP指纹图谱构建[J].湖南林业科技,2008,(4):8-19,23.
[6]李达,吴莉英,唐前瑞,等.非洲菊SRAP标记的DNA模板制备及反应体系优化[J].湖南农业大学学报(自然科学版),2008,34(4):196-199.
[7]张伟,曾黎琼,李媛娅,等.非洲菊不同组织总RNA提取[J].西南农业学报,2006,19(4):705-708.
[8]张玉进,孟祥春,潘瑞炽,等.非洲菊花瓣总RNA提取方法的改进[J].植物学通报,2001,18(6):722-726.
[9]聂京涛,潘俊松,何欢乐,等.非洲菊部分品种资源遗传多样性的ISSR分析[J].上海交通大学学报,2011,29(3):76-82.
[10]钟海丰,黄宇翔,钟淮钦,等.22个非洲菊品种遗传多样性ISSR分析[J].南方农业学报,2012, 43(1):1-4.
[11]孙勇如. 植物遗传手册[M]. 长沙:湖南科技出版社, 1989, 580-588.
[12]曾兵,张新全,范彦,等.鸭茅种质资源遗传多样性的ISSR研究[J].遗传,2006,28(9):1093-1100.
[13]沈颖,徐程,万小风,等.ISSR-PCR在石斛种间鉴别中的应用[J]. 中草药,2005,36(3):423-427.
[14]缪恒彬,陈发棣,赵宏波.85个大菊品种遗传关系的ISSR分析[J].园艺学报,2007,34(5):1243-1248.
[15]LEFFRING L.Flowers production in gerbera:I.Correlation between shoot,leaf and flower formatiom IN seedlings[J].Scientia Hortaiculturae,1973(1):221-229.
(責任编辑:柯文辉)
