3株鸽源新城疫病毒的分子特征及对鸽的致病性
2021-03-09杨少华黄庆华黄艳艳许传田
杨少华,崔 宁,张 琳,黄庆华,黄艳艳,许传田
(山东省农业科学院 畜牧兽医研究所 山东省畜禽疫病防治与繁育重点实验室,山东 济南 250100)
鸽新城疫是引起鸽急性、热性的高度接触性传染病之一,对养鸽业的危害巨大,其病原为鸽源新城疫病毒(NDV),又称鸽Ⅰ型副黏病毒(pigeon paramyxovirus type Ⅰ,PPMV-1)[1-4]。在分类上属于单股负链RNA病毒目(Mononegavirales)、副黏病毒科(Paramyxoviridae)、副黏病毒亚科(Paramyxovirinae)的禽腮腺炎病毒属(Avulavirus)[5-6]。NDV属于单股负链、不分节段RNA病毒,基因组长度约为15.2 kb,基因排列方式为 3′-NP-P-M-F-HN-L-5′,分别编码6种结构蛋白:核衣壳蛋白(NP)、磷蛋白(P)、膜蛋白(M)、融合蛋白(F)、血凝素-神经氨酸酶(HN)和大分子蛋白(L)[7]。NDV只有一个血清型,但基因型较多[1,8]。根据遗传进化分析可将NDV分为2类,即Ⅰ类(ClassⅠ)和Ⅱ(ClassⅡ)类,Ⅰ类NDV主要从野鸟中分离,多为弱毒株,基因组长度为15 198 nt,Ⅱ类NDV包含强毒和弱毒株,可分为18个基因型(Ⅰ~ⅩⅧ)[9],Ⅰ~Ⅳ基因型主要为1960年前分离的毒株,基因组长度为15 186 nt,Ⅴ~Ⅷ基因型主要为1960年后分离的毒株,基因组长度为15 192 nt[5,8,10-11]。从鸽中分离的NDV多为ClassⅡ基因Ⅵ型毒株,该基因型又进一步分为Ⅵa~Ⅵi共9个亚型[9,12-14]。Ⅵb、Ⅵf、Ⅵg亚型是我国2000年左右流行的主要基因型,常从鸽、鸡或其他家禽分离到[15-16],近年流行的毒株主要为Ⅵb基因型[17-19],Ⅵf、Ⅵg型很少分离到[4,20]。
多年来,鸽新城疫一直是危害养鸽业的重要传染病,发病率为30%~70%,死亡率为40%~80%,有时高达100%[1-4]。为了解山东鸽群NDV的分子流行情况,本研究对2011―2013年从山东分离的3株鸽NDV进行了序列测定和比对分析,并评估了其抗原性和致病性,以期为鸽NDV的防控提供科学的理论依据。
1 材料与方法
1.1 病毒株与单克隆抗体2011-2013年在新城疫的流行病学调查中从鸽场采集的疑似发病鸽的泄殖腔棉拭子中分离鉴定3株NDV:pigeon/China/SDS/2011、pigeon/china/SD01/13、pigeon/china/SD03/13,分别简称为SDS、SD01、SD03,对其进行纯化后-20℃保存备用。针对HN的单克隆抗体C3B3、1E5、2F10由我室制备并保存[21]。
1.2 主要试剂反转录试剂盒、pEASY-T3 Vector、DNA Marker、琼脂糖及胶回收试剂盒购自大连宝生物公司;DH5α感受态细胞购自北京全式金生物公司;TRIzol 购自Invitrogen 公司。
1.3 致病指数检测利用F蛋白裂解位点的氨基酸序列对3个分离株的致病性进行预测,同时按照OIE推荐的方法对3个分离株在10日龄鸡胚的致鸡胚平均死亡时间(mean death times,MDT) 和1日龄雏鸡的脑内接种指数(intracerebral pathogenicity indexes,ICPI)进行测定。
1.4 与单抗的反应性单抗C3B3 、1E5、2F10以PBS按1∶10 稀释,然后分别与200 TCID50的SDS、SD01、SD03病毒以等体积混合,37℃作用40 min,接种CEF细胞于37℃培养72 h ,测定细胞培养液的HA效价,以HA ≥2log2判定为阳性反应。
1.5 F和HN基因序列测定和分析TRIzol法从尿囊液中提取病毒RNA,所用引物及RT-PCR方法见文献[22]。PCR产物纯化后连接pEASY-T3 载体,转化DH5α感受态细胞,鉴定并挑取阳性克隆送公司测序。参照GenBank中发表的鸽NDV代表毒株序列,并根据F基因高变区(47~420 nt)核苷酸序列采用临位相连法(neighbor-joining)构建系统进化树。
1.6 分离株SDS的全基因组特征为了更好地了解分离株的遗传特性,选取毒株SDS,对其进行了全基因组测序并进行了序列分析。测序所用引物及克隆测序方法见文献[23],对SDS分段克隆测序,并进行拼接。应用Lasergene v7.1和MAGA 4.1基因分析软件对拼接的全基因组序列与GenBank中已发表的NDV代表毒株序列进行比对分析,并根据F基因高变区(47~420 nt)构建系统进化发生树。
1.7 分离株SDS的致病性30只NDV抗体阴性的健康试验鸽随机分为3组,每组10只,2个试验组分别以滴鼻和肌注方式接种0.1 mL 106EID50的病毒,对照组以0.1 mL灭菌生理盐水滴鼻,3个组分别于3个隔离器内饲养,每天观察记录采食、精神和粪便等情况。攻毒后3,5,7,14,21 d分别取喉头、泄殖腔棉拭子,常规处理后接种10日龄SPF鸡胚,检测其排毒情况;攻毒后7,14 d翅静脉采血,血凝和血凝抑制试验(HA-HI)检测NDV抗体水平;对病死鸽进行解剖,观察病理变化。
2 结果
2.1 生物学特性血清学试验显示,3个分离株与NDV标准阳性血清反应为阳性,与禽流感H5和H9亚型阳性血清反应均为阴性。SDS、SD01 和SD03分离株的MDT分别为89.5,71.2,62.2 h;ICPI分别为1.1,1.2,1.4;根据国际兽疫局(OIE)推荐的毒力判定标准SDS、SD01和SD03分离株均为中等毒力毒株。通过与HN特异性单克隆抗体1E5、C3B3、2F10的反应发现,疫苗株LaSota与3个单克隆抗体均反应,3个分离株仅与C3B3反应,与另外2个单抗1E5、2F10不反应,表明与疫苗株LaSota相比,3个鸽源NDV毒株的抗原性发生了改变(表1)。
2.2 SDS的全基因组序列特性与遗传进化分析通过测序和拼接,获得了除5′和3′引物区域外全基因组序列,其GenBank登录号为JQ993431。SDS基因组全长15 192 nt,基因排列方式为3′-NP-P-M-F-HN-L-5′。与NDV早期基因型(基因型Ⅰ、Ⅱ、Ⅲ、Ⅳ)相比,该分离株基因组在NP基因5′非编码区1 647~1 648 nt处插入6 bp碱基,插入序列为CCCCAA。3′端leader和5′端trailer序列分别为55 nt和114 nt,这与多数NDV毒株相同。NP-P、P-M、M-F、F-HN 和HN-L基因间隔区的序列长度依次为2,1,1,32,48 nt(图1)。通过全基因组序列同源性分析,SDS与2011年黑龙江分离株Pi/CH/CHLJ/131237(KJ607165)和山东分离株pigeon/China/SDLC/2011(JQ979176)的同源性较高,分别为99.2%和98.8%,与国外毒株PPMV-1/Belgium/98-238/199 (JX9011098)的同源性为95.8%,与经典强毒株F48E8及疫苗株LaSota、V4、Mukteswar的同源性分别为85.4%,83.0%,85.0%,85.7%。

A.基因组简图;B.基因起始和基因末端序列;C.基因间隔区序列
2.3 F、HN基因序列与遗传进化分析3个分离株的F基因全长1 792 bp,编码553个氨基酸,包含6个潜在的糖基化位点(85NRT87、191NNT193、366NTS368、447NIS449、471NNS473) 和13个保守的半胱氨酸残基(25,27,76,199,338,347,362,370,394,399,401,424,523)。融合肽(fusion peptide,117~141 aa)存在2处氨基酸替换:V121I和A132S,3个七肽重复区(heptad repeat region,HR)存在2处氨基酸替换,分别为V179I(HRa,143-185aa)和K480R (HRc,471~500 aa),而主要跨膜区(501~521 aa)相对保守。3个分离株F蛋白裂解位点均为112RRQKRF117,符合NDV强毒的分子特征。将分离株F基因高变区(47~420 bp)序列与41株NDV参考株序列进行遗传进化分析,结果显示3个毒株与2011―2017年山东、安徽、上海、广西、广东、云南、青海、黑龙江等地的鸽源NDV分离株的遗传关系较近,属于同一分支(图2),为基因Ⅵb型毒株;2013年的2个分离株SD01和SD03聚于一簇,遗传距离最近。
HN基因全长2 002 bp,编码571个氨基酸,含有5个潜在的糖基化位点(119NNS121,341NNT343,433NKT435,481NHT483,508NIS510)和13个半胱氨酸残基(123,172,186,196,238,247,251,344,455,461,465,531,542)且位置保守。与疫苗株LaSota相比,3个分离株HN蛋白上的5个抗原中和位点(193~201位、345~353位、513~521位、494位及569位氨基酸区域)存在多处氨基酸替换:R197I、N263K、E347G、D349E、R356K、G494D、D569E(表1),表明3个分离株的抗原性可能发生了一定的变化。

图2 分离株F基因高变区(47~420 bp)系统进化分析

表1 分离株HN蛋白抗原中和位点及与单克隆抗体的反应性
2.4 致病性试验结果分别通过滴鼻、肌注的方式用分离株SDS进行攻毒,攻毒后每天观察鸽的精神、食欲及发病死亡等情况,连续观察21 d。结果2个攻毒组试验鸽自攻毒后4 d采食量开始下降,精神沉郁;5 d临床症状明显,表现为翅膀下垂,流泪,扭头观星状,排黄绿色稀粪,采食量下降。滴鼻组和肌肉注射组的发病率为100%,死亡率分别为60%和70%。病死鸽剖检,可见脑膜充血,有点状出血点,腺胃乳头出血,腺胃肌胃交界处条纹状出血,肠道出血,肝脏肿大,脾有淤血斑。对照组鸽精神、食欲正常,无临床症状。
滴鼻组在攻毒后5,7,14 d喉头和泄殖腔有排毒,而肌肉注射组在攻毒后3,5,7,14 d喉头和泄殖腔均有排毒,排毒高峰在7 d,攻毒后21 d两个试验组均检不出排毒(表2)。分别于攻毒后7,14 d每组随机取5只鸽采血,测抗体,结果攻毒后7 d 3个组血清NDV抗体均为阴性,14 d时滴鼻组血清抗体为5~7 log2,肌肉注射组为7~8 log2,对照组仍为阴性。
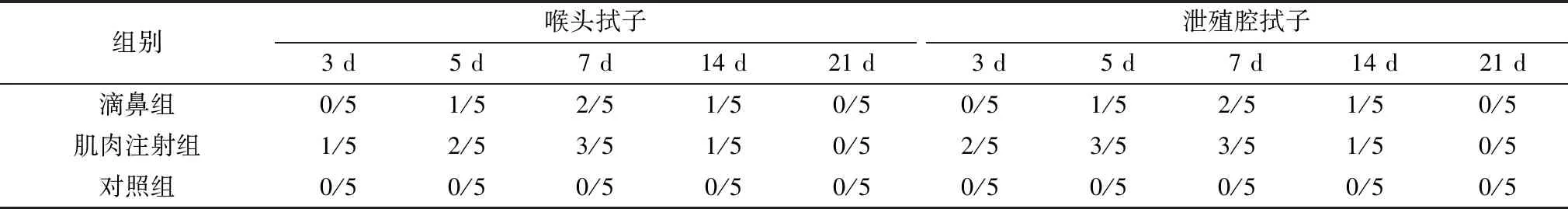
表2 试验鸽攻毒后不同时间喉头和泄殖腔排毒情况
3 讨论
本研究于2011-2013年从山东发病鸽群分离到3株鸽源NDV(SDS、SD01和SD03),根据F基因进行系统发育分析鉴定为Ⅵb基因型,与2011年黑龙江分离株Pi/CH/CHLJ/131237有很高的同源性。在过去的20年里,Ⅵb型NDV是世界范围内流行的主要基因型[1-3],目前分离到的鸽源NDV多为该型,其他基因型如:Ⅵa、Ⅵd、Ⅵf、Ⅵg较少分离到[4,18]。3株NDV的F蛋白裂解位点氨基酸序列均为112RRQKRF117,具有典型的强毒株特征;MDT分别为89.5,71.2,62.2 h,ICPI分别为1.1,1.2,1.4,比强毒株毒力低;根据OIE推荐的毒力判定标准SDS、SD01和SD03为中等毒力毒株,因此F蛋白裂解位点不是NDV毒力的唯一标准。
与疫苗株LaSota相比,3个分离株在F蛋白的融合肽和七肽重复区存在多处氨基酸替换:V121I、A132S、V179I和K480R,糖基化位点、半胱氨酸残基、蛋白跨膜区则比较保守;HN蛋白糖基化位点半胱氨酸残基高度保守,抗原中和表位存在多处氨基酸替换,例如:R197I、N263K、E347G、D349E、R356K、G494D和D569E。单克隆抗体细胞中和试验发现,与LaSota相比3个分离株与1E5和2F10的反应性发生改变,这可能与HN抗原位点突变有关,疫苗(如LaSota)产生的免疫压力可能是导致这一突变的主要原因,而抗原位点的突变又将导致疫苗的免疫失败。
为了了解Ⅵb型NDV对鸽的致病力,选取SDS分离株进行了鸽的攻毒试验。分别经滴鼻和肌肉注射攻毒鸽,感染后5 d鸽开始发病,症状与临床感染相似,表现为翅膀下垂,流泪、扭头观星状,排黄绿色稀粪,呼吸道症状不是很明显。滴鼻组鸽死亡率为60%,肌肉注射组为70%。而仇旭升等[4]报道用Ⅵb型鸽源NDV分离株ND132和ND167经口、肌肉注射、静脉注射攻毒鸽,鸽仅表现一过性临床症状,但感染后21 d仍可检出排毒。表明不同地区的鸽源NDV毒株存在遗传变异,致病力差异明显。临床上有的鸽带毒不发病,有的发病率和死亡率却很高,除了气候、环境、饲养条件和共感染等因素外,分离株的毒力差异也是一个重要原因。
近年来,随着新城疫的免疫防控措施的加强和养禽规模化集约化程度的提高,新城疫的发病率呈现逐年下降趋势,当前流行的NDV主要为基因Ⅵ型和Ⅶ型,其中基因Ⅵ型主要存在于鸽群中,基因Ⅶ型主要在鸡群和鹅群流行[24]。鸽或鸡感染鸽源NDV后均不同程度地通过喉头和泄殖腔向体外排毒,有的感染后21 d仍能检出排毒[4,20],成为新的传染源。鸽源Ⅵb型NDV曾导致20世纪80年代新城疫的第3次大流行[25-26],目前鸽源NDV分离株在疫苗免疫压力下抗原性正在逐渐发生变化,许多分离株与疫苗株LaSota间抗原性存在着显著性差异[4,18],这将对NDV防控构成一定的威胁,因此有必要采取一定的措施,加强鸽源NDV的防控。
