丝状真菌全局转录调控因子研究现状及发展
2020-11-24薛鲜丽刘博雅高紫君王德培
薛鲜丽,刘博雅,高紫君,王德培,*
(1.工业发酵微生物教育部重点实验室,天津300457;2.天津科技大学生物工程学院,天津300457)
丝状真菌的次级代谢是一个复杂且多层次的调控过程,其中涉及到真菌细胞感受外界环境(温度、pH值、渗透压、光照等)变化的适应、调控基因的表达、基因簇的转录激活和翻译修饰等过程。以上过程不仅受到途径特异性调控因子(如碳代谢调控因子CreA)调控,同时也被全局转录因子(如LaeA、VeA、McrA等)进行多重调控[1]。这些全局性转录因子的基因不以基因簇的形式存在,却可以调控一系列基因的表达[2]。丝状真菌在生长过程中很大程度上受外界环境条件的影响,当受到外界环境(如:强光照、高温、高渗透压、氧化胁迫等)刺激时,菌体通过基因调控表达相关基因来改变自身代谢方式,从而减少环境对微生物生长的不利影响,最终适应环境变化。研究发现,当菌株处于不利生长环境时,由全局转录因子参与的基因调控途径发挥着不可或缺的作用,这为提高菌株耐受性以应对不良环境提供了新的研究方向。针对丝状真菌而言,全局性调控因子通过调控相关基因的转录激活影响多种次级代谢物的产生,包括有益次级代谢产物(如:某些抗生素、降血脂药物洛伐他汀等)和有害次级代谢产物(如:黄曲霉毒、赭曲霉毒素等)[3-4]。目前,全局转录因子功能研究已经被应用于提高菌株耐受性、定向改造次级代谢物的产生、提高有益代谢物以及去除有害次级代谢物产生等。本文就全局性调控因子LaeA、VeA、McrA和LaeB对菌株耐受性提高、菌株生长形态改变及对次级代谢物产生影响等方面展开论述。
1 全局调控因子LaeA
1.1 laeA基因结构特征及作用机理
Bok等从构巢曲霉(Aspergillus nidulans)中分离鉴定到了laeA基因[5],随后的研究中发现laeA基因还广泛存在于其他丝状真菌中,如烟曲霉(Aspergillus fumigatus)、黄曲霉(Aspergillus flavus)、产黄青霉(Penicillium chrysogenum)、橘青霉(Penicillium citrinum)、米曲霉(Aspergillus oryzae)、黑曲霉(Aspergillus niger)等。由构巢曲霉laeA基因序列分析发现,该基因的启动子和编码区域分别有一个真菌毒素合成途径中的特异性调控蛋白AflR(黄曲霉毒素调控基因aflR)的结合基序,另外编码区的中游含有一个S-腺苷甲硫氨酸(S-adenosylmethionine,SAM)结合的保守的结构域,这个保守区域与其他曲霉中的laeA基因有较高同源性,如图1[1]。

图1 laeA基因结构示意图Fig.1 Structure diagram of laeA gene
研究发现,缺失SAM结合区域和缺失laeA全基因的菌株表型相同,由此推测laeA主要功能为一种甲基转移酶,其可以改变组蛋白甲基化水平。利用染色质免疫共沉淀方法鉴定出构巢曲霉中laeA会甲基化修饰组蛋白H3K9,进而使次级代谢产物柄曲霉素(sterigmatocystin,ST)的合成受到影响[6-8]。另外通过对序列的分析得出,laeA基因中AflR结合区域会影响次级代谢物的生成,基本机制是由于AflR对laeA具有负调控作用,并从转录水平得到验证,即通过转录水平分析发现AflR的过表达反而会下调laeA表达水平。众所周知,参与次级代谢和营养利用途径的基因均以基因簇的形式存在于真菌中。laeA则是黑曲霉次级代谢产物基因簇基因转录所必需的调控因子,但并不调控营养利用途径和spoC1孢子形成基因簇[8]。研究发现,△laeA突变株中,次级代谢产物柄曲霉素(ST)合成受阻,但在ST基因簇的外部额外拷贝一个aflR基因可以回补laeA缺失,使其调控的柄曲霉素(ST)的表达恢复。此外,将不受到laeA调控的argB基因(编码尿素合成过程中关键酶鸟氨酸氨甲酰基转移酶)整合到ST基因簇中,则会受到laeA的调控,进一步说明laeA调控下游基因簇与基因所在位置相关,而与具体基因功能关系不大[8]。近期研究表明,laeA对次级代谢物的调控具有位置偏好性,被其调控的基因簇多发生在染色体亚端粒区域[9]。除此之外,研究发现敲除LaeA导致曲酸的合成显著下降[10],其中kojR基因是曲酸合成基因簇中的关键基因[11],因此推测LaeA可能调控kojR基因的转录进而影响曲酸的产生,但这还需要进一步的研究确定。目前对LaeA调控机制已有大量研究,但是其调节机制细节还不是很透彻清楚。
1.2 LaeA对丝状真菌生长形态的影响
LaeA作为全局性调控因子,对多种丝状真菌生长进程和菌落形态有明显影响。Kale等的研究证明,LaeA影响菌核的形成,在A.flavus菌株中敲除或过表达laeA基因会相应导致无法产生菌核或者菌核的数量增加[12]。同样,Katarina等研究产黄青霉菌,敲除laeA会减少其孢子的形成[13];在A.fumigatus菌株中Bok等研究敲除或回补laeA基因对孢子影响与产黄青霉菌一致[14]。上述结果充分证明LaeA对丝状真菌生长和孢子形成存在不同影响。laeA缺失对丝状真菌生长形态及分生孢子的影响见图2。
如图2A和图2C所示,LaeA可影响红曲霉和橘青霉的菌落直径、孢子萌发和分生孢子形成,红曲霉△laeA的突变株(编号2)出现菌落异常(如图2A),比野生菌相同时间菌丝生长快、菌落直径大,而且未发现子囊孢子,分生孢子数量则是野生型的3倍(如图2B)。另外laeA基因缺失突变株分生孢子萌发率高于野生型[15],说明失去LaeA调控后,促进了红曲霉无性分生孢子的生成,同时失去有性繁殖子囊孢子的产生。周峰研究发现在橘青霉中过表达laeA基因,导致橘青霉在黑暗条件下的分生孢子产量较野生型降低9.9%(如图2D)[16]。但在构巢曲霉中缺失laeA基因不影响分生孢子的形成,可能机理是LaeA会调控Hülle细胞形成,该细胞功能为辅助分生孢子发育,laeA缺失突变株由于缺失该细胞会使得闭囊壳发育异常,与野生型相比较其所含的子囊孢子形状小且数量少仅为野生型的五分之一,但是其生育能力不受影响[17]。综上说明,LaeA对于不同丝状真菌中有性/无性繁殖孢子形成存在一定差异的影响,基于现有文献得到初步结论,LaeA对丝状真菌抑制无性分生孢子的产生,有助于有性生殖或子囊孢子的形成,但推测LaeA在参与调控有性/无性孢子形成时可能会存在不同调控机制,其相关参与的蛋白还有待于深入挖掘。

图2 laeA缺失对丝状真菌生长形态及分生孢子的影响Fig.2 Effection of ΔlaeA in filamentous fungi on morphology and conidiospores
1.3 LaeA对丝状真菌次级代谢产物的影响
LaeA的全局性调控功能对次级代谢产物的影响在很多丝状真菌中得到证实。目前由丝状真菌产生的具有生物活性的次级代谢物,被广泛研究,既包括对人类有益的如各种抗生素、降脂药物前体洛伐他汀等,也包括对人类有害的毒性或致癌性的次级代谢物,如:黄曲霉毒素、赭曲霉毒素等。研究人员发现增加laeA基因拷贝数可以激活沉默的次级代谢产物相关基因簇,进而使有益于人类的次级代谢物得到高产,例如在橘青霉中过表达laeA基因可使其次级代谢物美伐他汀的产量较对照提高5.42倍[16];在烟束曲霉(Aspergillus fumisynnematus)中laeA基因拷贝数的增加会提高环匹阿尼酸(cyclopiazonic acid,CPA)积累[18]。此外,运用敲除技术也可调控相关基因簇的表达进而抑制次级代谢物生成,在不同的丝状真菌中都有相关报道:Bok等研究发现在构巢曲霉中laeA基因的缺失,导致其不产生次级代谢产物柄曲霉素ST,如图3所示。

图3 LaeA参与次级代谢的调控Fig.3 Depicts of LaeA involvement in secondary metabolite regulation
黄曲霉毒素(aflatoxin,AF)是由黄曲霉或寄生曲霉产生的一类具有毒性和致癌性的次级代谢产物,在黄曲霉中敲除或过表达laeA基因会相应地抑制或促进真菌毒素AF的合成。其调控机制是,AF的合成是通过受控于G蛋白的信号通路,细胞外的刺激信号(如:氮源、碳源等)反馈于细胞核内,在核内受到全局性转录因子LaeA调控,经过AflR专一途径使其特异结合到AF基因簇的启动子区域,从而激活AF基因簇的表达和合成(如图3)[19]。同时相关研究表示,敲除laeA会影响细胞壁的疏水性,破坏了AF合成时所需要的细胞稳定性。综上所述,LaeA是通过影响了AflR蛋白表达进而影响AF合成启动,最终影响到AF的合成。研究表明在烟曲霉中laeA基因的缺失会减少其毒性次级代谢物胶囊毒素的产量[20];在产赭曲霉毒素的黑曲霉中敲除laeA也会减少赭曲霉毒素A(ochratoxinA,OTA)生产,尤其是在生长7 d后发生大幅度降低,在光照和黑暗条件下分别减少92%和99%左右,说明LaeA在OTA生产中起到了关键作用[21]。综合研究现状说明,在不同丝状真菌中敲除或过表达laeA能够显著影响不同的次级代谢产物(洛伐他汀等)的合成,而且都与AflR蛋白表达调节有关,但在不同的丝状真菌中可能存在的作用机制不尽相同,这还需要进一步地去挖掘影响次级代谢物的基因和条件。1

图4 H2O2对野生型和ΔlaeA突变菌株生长影响Fig.4 The effect of the H2O2on the growth of wild strain and ΔlaeA mutant strain
.4 LaeA对丝状真菌耐受性的作用
菌体在生长过程中往往会受到外界环境改变而带来的不利影响,如:温度、pH值、渗透压、光照及氧化胁迫等,环境改变会影响菌株的自身代谢过程进而导致其活性降低以及次级代谢物的生产力。目前,研究表明当菌体受到环境因素刺激,LaeA会参与环境胁迫时产生的应激反应,从而提高菌体的耐受性。例如,在黑曲霉的laeA敲除突变株,添加H2O2以构成氧化胁迫环境,其浓度依次为0、5、10 mmol/L,随着浓度的增加发现对环境的敏感性增加,如图4所示。
在0 mmol/L和5 mmol/L的浓度下,敲除株和野生型(WT)的生长状态较为相似,但是相同浓度条件下敲除株较对照生长明显受到抑制,且相比之下菌落直径也变小,而在10 mmol/L的浓度下敲除株的生长则完全受到抑制,而野生型仅表现为生长延迟;由于10 mmol/L的H2O2浓度下完全抑制敲除株,所以通过对5 mmol/L浓度下的缺失突变株和野生型的胞内活性氧自由基(reactive oxygen species,ROS)和抗氧化酶含量变化测定,进一步分析面对胁迫环境时胞内发生的防御机制,试验人员发现H2O2的添加使得野生株胞内ROS增加,抗氧化酶含量也相应增加,而相同条件下laeA缺失株的抗氧化酶含量较对照降低,说明laeA的缺失可能会影响抗氧化酶的活性使其在受到氧化胁迫时无法平衡过多活性氧带来的毒害,造成对胁迫环境敏感性的增加[21]。
2 全局调控因子VeA
2.1 VeA蛋白结构特征
Käfer等在构巢曲霉中首次发现全局性调控因子VeA[22]。构巢曲霉中的VeA蛋白由573个氨基酸组成,VeA蛋白N端包括一个核定位信号和核输出信号。研究表明,不同来源VeA同源蛋白的N端相对保守,C端具有种属特异性变化。经研究证实,VeA蛋白是一种光敏蛋白,它是Velvet蛋白家族的核心成员,其具有真菌特异性的高度保守的velvet结构域[23-25]。
2.2 VeA对丝状真菌生长形态的影响
HeeSeo Kim等对构巢曲霉的veA基因研究发现,其对有性繁殖正向调控,对无性繁殖进行反向调控[23]。这一发现为之后VeA对菌种生长发育和形态变化方面提供了不同研究方向。研究发现,不同来源veA基因对其生长形态的影响不尽相同,例如,Ana M.Calvo等试验发现在寄生曲霉(Aspergillus parasiticus)中菌核的形成仅发生在黑暗条件下,并且veA基因的缺失会导致无法形成菌核[26]。除此之外,还有对菌落形态、分生孢子产生及数量的影响,如在Aspergillus cristatus中敲除veA基因,较野生型仅进行有性繁殖相比,ΔveA突变株只生成灰绿色无性孢子,且无性孢子的数量是野生型的6倍多,如图5所示[27]。
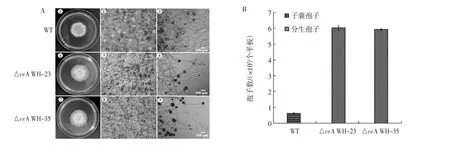
图5 野生型和ΔveA突变菌株在MYA(+1 mol/L NaCl)的平板上生长48 h生长形态及孢子的区别Fig.5 The differece of phenotypes and conidiation or cleistothecium production of wild strain and ΔveA mutant strain grown in MYA(+1 mol/L NaCl)for up to 48 h
橘青霉过表达veA基因会影响菌落形态,较野生型菌落褶皱增多且颜色变淡,在相同时间菌落直径与野生株没有区别,分生孢子数比野生株减少60%左右[28],表明veA基因的过表达抑制分生孢子形成,这与在构巢曲霉中过表达veA基因的试验结果相似。另外,在黄曲霉的研究中利用DNA微阵列分析以及实时荧光定量聚合酶链式反应(quantitative real-time polymerase chain reaction,qRT-PCR)等技术分析,得到缺失突变株和野生型之间有136个基因存在表达差异,并且有27个基因簇随时间变化表达差异显著,通过对基因簇的同源性和功能性分析,以及对可能依赖VeA的基因簇进行定量分析,发现VeA参与黄曲霉多个生长代谢的调控(如鞘脂类代谢),且敲除veA会提高无性产孢能力[29]。综上研究说明,VeA对不同丝状真菌的有性或无性繁殖的调控有一定的相似性,即抑制无性分生孢子的产生,有助于有性生殖或子囊孢子的形成,这一调控功能与LaeA功能正好一致。
2.3 VeA对丝状真菌次级代谢产物的影响
VeA作为全局性转录因子,在有益/有害次级代谢产物合成过程都有一定的影响。比如在橘青霉菌中,过表达可激活洛伐他汀合成基因簇中调控的基因mlcR和途径特异性调控基因mlcB,促进次级代谢物洛伐他汀的生成,通过转录组分析,mlcR和mlcB的表达量都明显高于对照;同时发现veA基因敲除会降低洛伐他汀的产量[28]。VeA可以抑制丝状真菌生长过程中有毒次级代谢物的产生,如在合成黄曲霉毒素的过程中aflR基因和aflJ基因转录表达是必不可少的,Ana M.Calvo等研究veA缺失黄曲霉突变株发现,aflR基因和aflJ基因都无法转录,推测VeA可能对aflR基因和aflJ基因转录有开关作用,进而影响AF的合成[29];此外,通过RT-PCR分析,发现黑曲霉veA敲除株赭曲霉素OTA合成关键基因pks表达水平降低,导致该突变株无法合成OTA[21]。
2.4 VeA对丝状真菌耐受性的作用
VeA在调控丝状真菌次级代谢产物合成的同时,也参与菌株对环境胁迫的适应性响应,主要是通过改变相应基因表达和改变代谢途径提高菌株对环境胁迫的耐受性。例如在黑曲霉中敲除veA基因会增加对氧化胁迫的敏感性,在添加过氧化氢构成氧化胁迫环境条件下,△veA黑曲霉突变株随着过氧化氢浓度的增加生长逐渐受到抑制,在10 mmol/L过氧化氢浓度下菌株生长被完全抑制,而对照组仅表现出随着过氧化氢浓度的增加生长迟缓,在相同条件下可以生长形成菌落[21]。另外,与构巢曲霉亲缘性很近的海洋灰绿曲霉(A.glaucus)中也存在全局转录因子VeA。海洋生物往往易受到盐浓度渗透压胁迫,相关研究表明,VeA在海洋灰绿曲霉中可响应盐度信号调控菌体生长和发育,无论是敲除突变株还是野生型,其菌落大小都会随着盐度的降低而变小,但是在相同的盐度条件下缺失株和野生型的菌落大小也存在差异。在发育过程中,处于人工海水和等渗溶液条件下,敲除株与对照株的菌落和生长速率较为相似,分生孢子的数量随盐度变化趋势也相似;但在去离子水的胁迫条件下,敲除突变株在生长发育的后期较野生型菌落边缘出现更多不规则的细小菌丝,且分生孢子数量较野生型增加。分生孢子的增加说明菌株倾向于无性繁殖。还有研究发现灰绿曲霉的无性繁殖可以促进次级代谢物抗肿瘤灰绿曲霉A的产生,这也为利用VeA全局调控A.glaucus在盐度变化的条件下高产灰绿曲霉A提供参考[30-31]。综上研究说明,利用全局转录因子VeA提高菌株在发酵过程中对不良生长环境的耐受性,对今后的生产实践具有重要意义和应用前景。
3 Velvet复合体中全局转录因子之间的协同作用
Bayram等在构巢曲霉中发现VeA作为桥梁连接LaeA和VelB(the velvet like B,属于Velvet蛋白家族),使其成为异源三聚体,命名为Velvet复合物,调控丝状真菌的形态发育和次级代谢,其调控机制如图6所示。
在光照条件下,VeA多保留在细胞质中,导致VeA无法作为桥梁连接LaeA和VelB形成Velvet复合物,游离VelB可促进无性孢子形成,此时LaeA表现出低活性且无法形成异源三聚体,从而抑制有性繁殖;当处于黑暗环境时,在α输入蛋白KapA和VeA的核定位信号相互作用下,使进入细胞核的VeA量增加,此外,VelB也随着VeA进入核内并增多,从而形成影响次级代谢物基因簇表达的Velvet复合物,进而促进有性繁殖和次级代谢产物的生成[32-33]。Velvet复合物在不同真菌中既有保守性又有一定的特异性,而复合物形成在调控代谢过程和生长发育方面的相互调节和协同作用还需要进一步的深入研究。

图6 Velvet复合体中全局转录因子之间的协同作用Fig.6 Synergy between global transcription factors in the velvet complex
4 全局调控因子McrA
4.1 McrA结构特征及对丝状真菌生长形态的影响
McrA是近期在构巢曲霉中发现的一类可能属于锌指结构转录因子的全局调控因子[34]。对于全局性调控mcrA基因的结构特征不如laeA基因的研究深入全面,目前只在构巢曲霉、黄曲霉[33]、青曲霉(Penicillium)和土曲霉(Aspergillus terreus)中有相关报道。在近期的研究中发现,黑曲霉中存在与构巢曲霉中McrA同源性极高的基因,在黑曲霉CBS513.88中,mcrA的缺失会对菌株的生长形态产生一定影响,例如敲除mcrA菌株在MEPA培养基中的表型由野生株所呈现的黑色菌落变为黄色菌落,并且菌落直径较野生型变小,在其他培养条件下(4种真菌培养基PDA、YG、CYA、YMEG),敲除株比野生株的菌落直径都有不同程度的减小,孢子颜色都发生改变而且颜色变化差异较大,证明敲除突变株的生长速率要低于野生型[35]。同时推测外界不同的环境刺激信号可能使McrA在调控同一个真菌时能产生不同的调控机制,作用于不同的基因导致孢子颜色和大小的差异。
4.2 McrA对丝状真菌次级代谢产物及耐受性的作用
mcrA基因不仅对菌株形态变化有影响,还具有调节次级代谢产物的功能。全局性调控因子McrA在构巢曲霉中被鉴定为是一种负调控因子,由转录组分析结果得到敲除mcrA显著改变了1 352个基因的转录,其中623个基因上调了5倍,且试验中发现mcrA基因缺失突变株至少上调了6个与次级代谢产物相关的转录因子,如上调AN7819转录因子,推测其为ST基因簇的一个调控基因,过表达则会抑制次级代谢物的产生,在青霉菌和土曲霉的相应研究中也得到一致的结果[33,35]。在对黑曲霉mcrA基因研究发现,敲除mcrA突变株产生的次级代谢物通过与对照组差异分析,比对得到衣康酸(hexylitaconic acid)的衍生物(3S,8R)-8-hydroxy-3-carboxy-2-methylenenonanoic acid和(3S)-9-hydroxy-3-carboxy-2-methylenenonanoate,以及具有抗菌活性的衣康酸衍生物曲衣康酸A-C(asperitaconic acid A-C),并且通过定量分析发现,McrA正调控前两种化合物,负调控后者,但是具体的调控机制还有待研究。在对抗环境胁迫方面,mcrA基因也起到一定的调控作用。黑曲霉mcrA基因缺失突变株的研究中发现,在高盐高渗的胁迫条件下,敲除突变株对Zn2+、Li+离子环境与野生对照组相比敏感性降低,对Na+环境敏感性升高。说明不同的胁迫环境可能在影响McrA调控黑曲霉时存在差异,其具体作用机理还有待于进一步发现[36]。
5 全局调控因子LaeB
前期的研究报道中发现,在植物内生菌无花果拟盘多毛孢存在一类全新的全局调控性因子RsdA[37],且在模式菌株构巢曲霉中也发现其同源基因laeB。该基因是由其表型(loss of aflR express)而命名,预测其包含同源性较低的两个区域:G-蛋白通路抑制因子和转录起始因子ⅡA结构域[38]。经过试验证明,LaeB调控构巢曲霉的次级代谢,缺失laeB基因会间接影响杂色曲霉素的合成。laeB对黄曲霉在不同培养基上菌体生长及分生孢子的影响见图7。

图7 laeB对黄曲霉在不同培养基上菌体生长及分生孢子的影响Fig.7 The effect of phenotypes and conidiation production of wild strain and ΔlaeB mutant strain grown in different culture
在黄曲霉中,laeB基因缺失突变株较野生型发生明显变化(如图7A),如在PDA培养基上培养的两个菌株的对比,△laeB突变株的分生孢子产量要明显多于对照(如图7B)。通过荧光定量PCR分析分生孢子发现突变株brlA基因的表达量上调,brlA则是分生孢子形成的关键基因,推测LaeB通过brlA调控基因进而影响菌落形态;而在GMM培养基的条件下,△laeB突变株的菌落显著变小,生长受到抑制。同时代谢产物中无法检测到黄曲霉毒素B1。荧光定量PCR分析表明,laeB基因缺失菌株影响黄曲霉毒素B1相关合成aflQ基因以及调控aflR基因等表达,这些基因的表达下调可能导致减少合成黄曲霉毒素B1[39-40]。目前laeB基因只在构巢曲霉和黄曲霉中有相关研究,在其它丝状真菌菌株中未见报道,因此对其具体调控机制以及对菌株形态变化的影响还需要进一步的研究。
6 总结与展望
丝状真菌是一类重要的微生物,其代谢产生的物质众多,目前已知微生物次级代谢产物占总数的一半左右,并应用于生物医药和食品发酵等领域,产生着巨大的实际应用价值,因此近年来丝状真菌及其代谢工程研究成为热点。目前,通过对丝状真菌大量的基础研究发现,其次级代谢途径是一个复杂且多层次的高精度调控网络,包含转录、翻译及表型遗传水平等多层次调节,其中每一层次都有多种蛋白质参与。随之直接或间接地调控这些蛋白质表达的全局转录因子被发现并得以应用。通过对全局转录因子的基因鉴定、结构分析以及为阐明其调控机制的研究,可以定向改造丝状真菌调控次级代谢物产生相关基因的转录表达。通常采用敲除或过表达基因的方法,激活丝状真菌中调控对人类有益的次级代谢物(如抗生素类药物、降血脂药物等)产生相关基因;抑制调控对人类有害的具有毒性和致癌性次级代谢物(如黄曲霉毒素、赭曲霉毒素等)产生的基因。这些次级代谢产物的研究及其在农业、食品工业以及医疗卫生等方面的应用具有重要价值。此外真菌在生长或发酵过程中往往会受到不同程度及条件的环境胁迫,而其生长发育的改变又会影响到次级代谢物的合成和产量。在工业生产中为避免不良环境影响菌株生长,常规是人为改变外界坏境条件,例如采取冷却水循环系统降低由夏季高温引起的不利影响,但是这同时也对生态坏境资源造成一定的浪费,增加了生产成本。因此利用全局转录因子改善菌株的耐受性以平衡由环境改变带来的不利影响以及减少对生态环境的危害也已逐渐成为领域内所关注的焦点。
根据上述的研究总结,不难发现对于全局转录因子间的相互调节和协同作用的研究还不够深入。因此laeA、veA基因和Velvet复合物等其具体的作用机制和调控代谢网络需要更加系统深入的研究,同时寻找和挖掘更多的新型全局转录因子(如LaeA的相似功能基因等),为丝状真菌在各领域更广泛地发展提供更多的可能。这就需要在今后的研究中采取更加全面、更加精确的技术来研究和完善全局转录因子的理论基础,如采用代谢组学、转录组学等方法以及结合生物信息学,对目的基因或基因簇进行检测分析和编辑,更加系统地设计改造基因,为调控次级代谢途径及次级代谢产物的产生提供新方法新思路,也为今后工业生产发酵提供新的理论基础和技术支持。
