EuCAD基因不同位置片段RNAi对烟草木质素合成的调控效果
2020-10-21陈博雯肖玉菲李军集覃子海张晓宁刘海龙
陈博雯, 肖玉菲, 李军集, 张 烨, 覃子海, 张晓宁, 刘海龙
(广西壮族自治区林业科学研究院 国家林业局中南速生材繁育实验室/广西优良用材林资源培育重点实验室, 广西 南宁 530002)
桉树(Eucalyptus)是桃金娘科桉属植物的总称,原产地为澳大利亚[1-2]。目前,全世界有100多个国家种植桉树,在我国热带、亚热带地区引种种植已有100多年的历史[3-5]。由于桉树速生丰产并且材质优良,已经成为短周期原料林的主要造林树种,被广泛用于制浆造纸、人造板和纤维板等,特别是在造纸木浆生产中具有重要地位[6-8]。
在传统选育优良纸浆材品种的基础上,利用基因工程手段改良木材品质是国际上研究的热点[9-10]。尾叶桉GLU4无性系(Eucalyptusurophyllaclone GLU4)是广西广泛种植的桉树良种,以此树种为基础,采用转基因技术培育低木质素桉树品种,具有良好的应用前景。为此,广西林科院已经率先在尾叶桉良种GLU4的基础上开展了木质素调控的转基因技术研究,且经过前期研究已取得一定进展,在一系列木质素合成途径基因中,已锁定应用尾叶桉肉桂酰乙醇脱氢酶基因(EuCAD)转化烟草可抑制木质素合成[11-12]。在前述对EuCAD基因研究中使用的是基因全长序列构建RNA干扰载体,序列较长,载体构建和应用均不便捷,而RNAi技术的优势之一,就是利用较短的干扰片段进行转录后调控。为此,选择模式植物烟草,采用生物信息学工具分析其EuCAD基因序列,根据编码蛋白保守结构域的分布情况分别选择EuCAD基因的部分序列构建RNA干扰抑制表达载体并转化植物,利用遗传转化体系检测分析不同位置选取的干扰片段对烟草CAD基因表达及木质素合成的调控效果,筛选鉴定出EuCAD中RNAi效果最佳的基因片段,以期为尾叶桉GLU4无性系肉桂酰乙醇脱氢酶基因(EuCAD)调控其木质素的合成提供技术参考。
1 材料与方法
1.1 材料
1.1.1 组培苗及菌株 烟草无菌组培苗、DH5α菌株和农杆菌LBA4404菌株,由广西优良用材林资源培育重点实验室保存。
1.1.2 试剂盒 RNA prep Pure多糖多酚植物总RNA提取试剂盒,天根生化科技有限公司;RNA LA PCR反转录试剂盒,大连宝生物工程有限公司;SG Fast qPCR Master Mix(2X),购自上海生工生物公司。
1.1.3 仪器 Nikon Eclipse80i光学显微镜,尼康公司;LightCycler480荧光定量PCR仪,罗氏公司。
1.2 方法
1.2.1 RNAi载体的构建 利用Blastp、ProtParam、psipred、SWISS-MODEL等对EuCAD基因序列分析其编码蛋白的保守结构域,其分布在40~300氨基酸。根据保守结构域的分布情况,分别选择包含NAD(P)结合位点、底物结合位点和Zn结合催化位点的S1片段;包含多个Zn结合结构位点的S2片段;包含多个NAD(P)结合位点的S3片段,以及带有NAD(P)结合位点和二聚体结合界面序列的S4片段用于构建RNAi载体。每条选定片段利用Vector NTI分析筛选出40 bp左右片段,序列信息为S1-gaatttgccacagtgacattcaccagatcaagaatgatcttggcgc,S2-tggttgggtgccgcagaagctgtggcccttgcaattcggaccagctg,S3-atgtgcgctggtgtgaccgtgtacagtcctctggtgcgctttgg,S4-ccatggttatgcttgggaggaagtcaatcactgggagtttcata。利用序列设计成反向重复序列,在每条序列两端附加XhoI酶切位点后,交由上海生工生物公司合成序列,得到携带目标片段的克隆载体pUC57-S1、pUC57-S2、pUC57-S3和pUC57-S4。利用XhoI酶分别消化pUC57-S1、pUC57-S2、pUC57-S3和pUC57-S4,通过凝胶电泳回收得到目标序列,同时利用XhoI酶消化pCAMBIA3301载体,同样经过凝胶电泳回收后,得到去除bar基因序列的载体骨架。分别将载体骨架与4条合成序列连接,连接产物转化DH5α,应用载体上的测序引物3301-F(5'-CCCTTATCTGGGAACTACTCAC-3')/3301-R(5'-CGCTGAAATCACCAGTCTCTC-3')对转化进行PCR鉴定,对可扩增得到目标长度片段的克隆进行测序,测序结果与目标序列比对,序列一致即为构建成功的RNAi载体。RNAi载体构建成功后,按常规方法导入农杆菌LBA4404中用于转化。
1.2.2 烟草转化植株的培育 利用RNAi载体转化获得的4个转基因烟草植株,经过愈伤组织培养和芽再生培养,经抗生素筛选获得阳性植株,移栽后置于温室内培养,同时保留每个植株的组培继代芽,以备后续扩繁培养转基因植株之用。移栽60 d后,取叶片提取基因组总DNA。应用载体上的测序引物3301-F/3301-R,对转基因植株进行PCR鉴定,同时对长度相符的PCR扩增产物测序以验证插入的RNAi序列准确性。
1.2.3 转基因烟草植株CAD基因的表达水平 4个转基因烟草植株移栽培养240 d后,以野生型(WT)植株为对照,自茎尖向下第3节处取叶片,提取RNA用于CAD基因表达水平检测。
采用烟草actin基因作为内参基因,烟草CAD(X62343.1)基因为目标检测基因,利用Vector NTI设计引物序列如下:
actin-F:CTGGAATCCATGAGACTACTTACAA;actin-R: AACCGCCACTGAGCACAATA;CAD-F: GTATGGCACCAGAACAAGCAG;CAD-R: CCAATGCCTCTTGTCTCTTCTTAT。
植物组织样品采用试剂盒提取并反转录合成cDNA。PCR检测条件为95℃持续3 min,40次循环,95℃持续7 s,55℃持续10 s 和72℃持续15 s。每个处理设3个生物重复及3个技术重复。
1.2.4 指标测定
1) 木质部厚度测定。取温室培育270 d的4个转基因烟植株,以野生型(WT)植株为对照,自茎部顶端向下数,选取其茎部第4节和第6节茎部组织,按常规方法制成石蜡切片,并用番红-固绿法染色[13]。制好的切片用Nikon光学显微镜进行观察,NIS-Elements拍照并测量茎部直径。测量时每个样品选取1张切片,在茎部截面的各个方向分别采集60个茎部直径数据和木质部厚度数据。
2) 木质素及纤维素含量测定。取温室培育270 d的4个转基因烟植株,以野生型(WT)植株为对照,选择自顶端向下第3~10节茎部,去除叶片后65℃烘干24 h,粉碎后过40目筛,用于木质素和纤维素含量检测。
木质素含量:参照GB/T20808-2006要求采用酸性洗涤方法测定。称取1 g样品,加入2%CTAB和2~3滴正辛醇后煮沸1 h。消煮液用玻璃砂漏斗抽滤后,将漏斗转移至50 mL烧杯中,加入15℃的12.0 mol/L硫酸至半满,将结块打碎并搅拌呈糊状,20℃保温3 h,期间根据流出量不断补充12.0 mol/L硫酸。消解后抽滤并用热水洗涤至pH试纸检测呈中性。取出漏斗和残余物置于105℃烘箱中烘干至恒重。
木质素含量=烘干后木素残渣质量/绝干样品质量×100%
纤维素含量:采用硝酸法测定。称取1 g样品置于锥形瓶中,加入25 mL硝酸-乙醇(4∶1)混合液,装上回流冷凝器,沸水浴1 h,期间随时震荡烧瓶。静置至残渣沉积后小心倒出上清液至恒重的玻璃滤器中,用真空泵吸干滤液并将流入滤器中的残渣移回锥形瓶中。量取25 mL硝酸-乙醇混合液分数次将滤器及锥形瓶口附着的残渣移入瓶内。再次装上回流冷凝器,沸水浴1 h。3次重复后,将锥形瓶内容物全部移入滤器,用10 mL硝酸-乙醇混合液洗涤残渣,再用热水洗涤至洗液中性,最后用乙醇洗涤2次,吸干滤液,滤器置于105℃烘箱中烘干至恒重。
纤维素含量=烘干后纤维素残渣质量/绝干样品质量×100%
1.3 数据处理
采用SPSS 19.0进行统计与分析。
2 结果与分析
2.1 RNAi载体的构建
从图1看出,pCAMBIA3301载体上的测序引物3301-F/3301-R分别位于XhoI酶切位点旁侧约50 bp处,XhoI酶切位点之间的bar基因序列分别被替换为S1~S4的目标序列,PCR产物长度从650 bp左右缩短至约200 bp。每个载体都检测得到扩增出目标特异性条带的阳性克隆,对阳性克隆提取质粒测序结果显示,插入序列与预期序列一致,RNAi载体构建成功,分别命名为pCAMBIA3301-S1、pCAMBIA3301-S2、pCAMBIA3301-S3和pCAMBIA3301-S4。
2.2 烟草转化植株的培育
从图2可见,RNAi载体转化获得的4个转基因烟草植株基因组上插入RNAi载体的转基因植株中可检测到长度200 bp的特异性片段,而未整合RNAi载体的植株则未扩增到目标条带。阳性植株经进一步PCR扩增产物测序结果显示,其序列与目标RNAi序列一致,获得的转基因烟草植株分别命名为pS1、pS2、pS3和pS4。
2.3 转基因烟草植株CAD基因的表达水平
从图3看出,pS1、pS2、pS3和pS44个转基因烟草植株的CAD基因表达水平与野生型(WT)相比,均呈显著下降趋势,pS1与pS2间差异不显著,pS1和pS2显著低于pS3和pS4,pS3显著低于pS4。其中,pS1和pS2降幅最大,分别为87.48%和87.30%;pS3降幅其次,为67.93%;pS4降幅最小,为49.70%。从基因表达调控效果看,RNAi片段S1和S2的调控效果最好。
2.4 转基因烟草植株茎部直径及木质部的厚度
从表1可知,野生型和4个转基因烟草植株茎部直径及木质部厚度的变化。直径:第4节和第6节分别为6 194.57~6 634.29 μm和8 293.15~8 576.63 μm,依次分别为WT>pS2>pS4>pS1>pS3和WT>pS4>pS1>pS2>pS3,不同植株间差异均不显著。木质部厚度:第4节为283.21~320.91 μm,依次为WT>pS4>pS3>pS2>pS1,WT与pS3和pS4差异不显著,但显著厚于pS1和pS2;pS1、pS2、pS3和pS4间差异不显著。第6节为384.53~427.53 μm,依次为WT>pS3>pS4>pS2>pS1,WT显著厚于pS1,与其余3个植株间差异不显著;pS1、pS2、pS3和pS4间差异不显著。
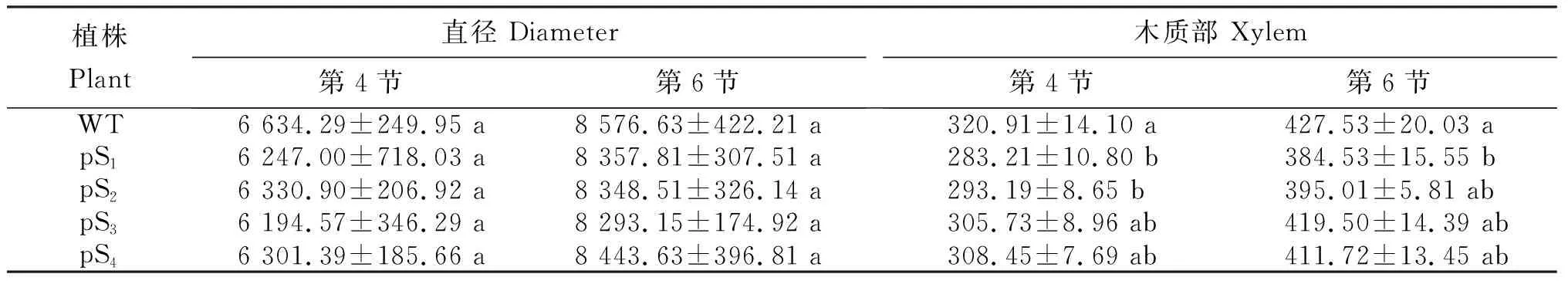
表1 转化植株茎部直径及木质部的厚度Table 1 Diameter and xylem thickness in the stem of transformed plants μm
2.5 转基因烟草植株茎部木质素及纤维素的含量
从表2可知,野生型和4个转基因烟草植株木质素及纤维素含量的变化。木质素:WT最高,为13.37%;pS4其次,为12.36%;pS1最低,为11.33%;WT显著高于其余植株,pS1显著低于其余植株,其余植株间差异显著。pS1、pS2、pS3和pS4分别较WT降低15.26%、11.44%、10.62%和7.55%。纤维素:pS3最高,为36.08%;WT其次,为35.16%;pS1最低,为31.43%;pS3显著高于其余植株,pS1显著低于其余植株,其余植株间差异显著。pS1、pS2和pS4较WT分别降低10.61%、5.03%和3.10%,pS3较WT提高2.62%。

表2 转基因烟草植株的木质素及纤维素含量Table 2 Lignin and cellulose content in transgenic tobacco %
3 结论与讨论
由于调控靶序列的特异性和高效性,RNAi已成为转录后调控的首选方法[14]。在对目标基因序列不清楚时,RNAi常采用基因全长序列构建RNAi载体,这种常规研究策略在许多植物中取得了较好的效果。但也有研究者指出,干扰片段过长,会使自身复制过程中碱基错配几率增大,同时较长的序列更容易与其他非目标基因序列匹配,脱靶风险更高[15]。若选择目标基因的部分片段用于RNAi,所选择的位置至关重要。HOLEN等[16]根据目标基因的不同位置设计4条21~23 bp的RNAi片段研究发现一种可能的位置效应,调控效果最佳的RNAi片段hTF167i和hTF372i能够抑制85%~90%的基因活性,而 hTF562i片段只表现出一种中间效应,hTF478i的活性则很低;且进一步深入研究发现,针对靶点hTF167设计的干扰片段hTF158i和 hTF161i与hTF167i序列仅相差几个碱基,几乎完全丧失调控效果。张斌等[17]对OsPUT1基因的RNA干扰研究也提出类似的位置效应,基因中段序列的干扰效果优于首尾片段。由此推断,针对目标基因选择RNA干扰片段时,既应考虑片段在基因CDS中的位置,也应考虑片段包含的特异性位点。
鉴于CAD蛋白在木质素合成中的重要功能,在许多植物中都对其展开了研究,目前为止,NCBI已收录2 428条陆生植物CAD蛋白序列,不同植物中的CAD具有高度的同源性。对CAD蛋白序列比对分析发现,几乎所有CAD都具有三类典型的保守的结构域。第一类是辅酶和底物结合结构域,包括NAD(P)结合位点和底物结合位点。第二类是酶活性中心结构域,包括Zn结合催化位点和Zn结合结构位点。第三类是二聚体结合结构域。对CAD蛋白三级结构分析表明,该蛋白以二聚体形式存在,涉及到二聚体结合界面的结构域对二聚体的形成与稳定具有极其重要的作用[18]。
研究分别选择包含NAD(P)结合位点、底物结合位点和Zn结合催化位点的S1片段,包含多个Zn结合结构位点的S2片段,包含多个NAD(P)结合位点的S3片段,以及带有NAD(P)结合位点和二聚体结合界面序列的S4片段用于构建RNAi载体。在将4条RNAi片段分别转化烟草后发现,S3和S4片段的调控效果不理想,多项表型检测结果与野生型相比降幅较小或无差异。S1和S2片段对木质素的调控效果则最为明显,从基因表达水平到木质素含量和木质部厚度均较野生型降低。
从序列编码蛋白的结构域看,S1片段编码蛋白兼具辅酶、底物结合结构域和酶催化结构域,生物学功能重要;S2片段编码蛋白则几乎涵盖CAD中所有预测的Zn结合结构位点,缺失前述任一段蛋白片段势必极大地影响酶学活性。从基因序列位置分析,S1片段近于转录翻译的起始端,S2片段也位于基因序列的前1/3处,若RNA干扰在S1或S2片段发生,CAD酶几乎无法转录或翻译,对酶活性的抑制将更明显。前述分析结果与试验结果相一致,说明S1和S2片段是较为理想的RNAi片段。
S3和S4片段在设计时涵盖了大多数的预测NAD(P)结合位点和二聚体结合界面的结构域,但S3和S4片段的调控效果却甚微,可能是由于S3和S4片段引发RNA干扰时,由于片段位于基因中后部,大部分序列仍可翻译为残损的CAD蛋白,残损蛋白虽缺失上述片段编码的结构域,但蛋白的其他NAD(P)结合位点和二聚体结合界面的结构域位点可能仍具有活性,所以残损蛋白仍具有生物学活性。此外,研究中选取的片段长度为44 bp,在后续研究中应进一步优化缩短干扰片段,有望获得高效RNAi片段。
研究结果表明,植物茎部结构特点是从上到下木质化程度逐渐提高,而在pS1和pS2植株中,随着木质化程度的提高,通过抑制CAD基因表达调控木质素合成的效果逐渐弱化。造成此变化的原因可能是CAD对木质素合成的调控主要发生在木质化初期,在木质化中后期,其调控效果被其他代谢通路中的基因应答作用所掩盖。同时,pS1和pS2植株中木质部厚度的降幅均在10%左右,降幅较低,较小的降幅可能在木质化过程中被逐渐积累的木质部厚度稀释,造成降幅差异逐渐减小。对此,在后续研究中,可尝试使用底物标记等手段,分阶段解读木质化过程,有望深入揭示抑制CAD基因调控木质素合成的生理生化机制。
