棉花角斑病菌V8高效电转化条件的探索
2020-08-28王清孟凡奇张勇跃缪卫国刘志坚
王清 孟凡奇 张勇跃 缪卫国 刘志坚
摘要:对去甲基化氮胞苷(5-AZA)的最佳浓度以及影响棉花角斑病菌[Xanthmonas campestris pv.malvacearum (Xcm) lV8电转化效率的诸多因素进行逐一研究,以期获得高效的电转化条件。结果表明,选取100 limol/L的5-AZA驯化处于对数中期的V8制备感受态细胞;保证电压2.1 kV、50 μg/mL质粒7μL、4℃预冷复苏培养基SOC、振荡复苏培养4-5 h时,V8的电转化效率高达5×102/μg DNA。
关键词:棉花角斑病菌[Xanthmona scampestris pv.malvacearum (Xcm) ]V8;菌株驯化;电转化条件;电转化效率
中图分类号:Q78;S435.621.2+5
文献标识码:A
文章编号:0439-8114( 2020)12-0175-04
D01:10.1408 8/j .cnki.issn0439-8114.2020.12.039
开放科学(资源服务)标识码(OSID):
在中国,棉花是重要性仅次于粮食的作物,棉花细菌性角斑病是多数棉花种植区重要的细菌病害之一,对棉花整个生产过程造成了较大的负面影响[1]。该病菌的病原是野油菜黄单胞菌锦葵致病变种[Xanthmonas campestrispv. malvacearum (Xcm)][2]。黄单胞菌属是植物病原菌中较大的类群[3],随着细菌基因组学的发展,主要致病菌的hrp基因功能[4]等研究也更加深入。限制性修饰系统能抑制细菌基因组外来DNA交换引起的变化,降解异源DNA进而保护细菌[5],导致Xcm的8号小种V8电转化效率为零,推测这可能是hpaXm及hpaXm-IP基因通过电转化而定点敲除失败的原因。
通過去甲基化氮胞苷(5-AZA)驯化野生菌株[ 5],突破了V8电转化效率为0的限制,成功对V8进行电转化,并证实5-AZA使用浓度在100 μmol/L时驯化菌株最高效。以从V8基因组中克隆出的基因hpaXm及其信号肽类似序列hpaXm-IP为靶标基因,以前期构建的两个重组质粒pK18mobsacB::N1012和pK18mobsacB::m762为待转质粒,对影响V8电转化的一系列因素,如细胞生长状态、电压强度、质粒浓度及用量、复苏方式及时间、复苏培养基种类及温度等进行分析,为后续V8其他基因的定点敲除提供参考。
1 材料与方法
1.1 材料
菌株为棉花角斑病菌V8,从棉花上分离。质粒为pK18mobsacB:: N1012及pK18mobsacB:: m762,前期构建。主要溶液为去甲基化氮胞苷溶液。
1.2 方法
菌株驯化参考李岗[5]的方法。
1.3突变菌株V8△hpaXm和V8△hpaXm-lP的获得
电击转化获得转化子,具体验证参考王清等[6]的方法。
1.4去甲基化氮胞苷驯化V8的浓度筛选
采用去甲基化氮胞苷驯化V8,将V8电转化的条件及各参数基本保持一致,具体为采用对数期V8制备感受态、电压2.5 kV、振荡复苏3h、常温复苏培养基NB等,筛选5-AZA的最佳使用浓度。
1.5 不同条件对V8电转化效率的影响
将每个可能影响电转化效率的因素当作1个处理,每个处理3次重复,电转化后置于28℃恒温培养约4d,计算电转化效率。电转化效率=转化子总数/μg DNA;转化子数目=单菌落数×转化原液总体积×稀释倍数/涂板菌液体积[7]。
2 结果与分析
2.1不同浓度的去甲基化氮胞苷对V8电转化效率的影响
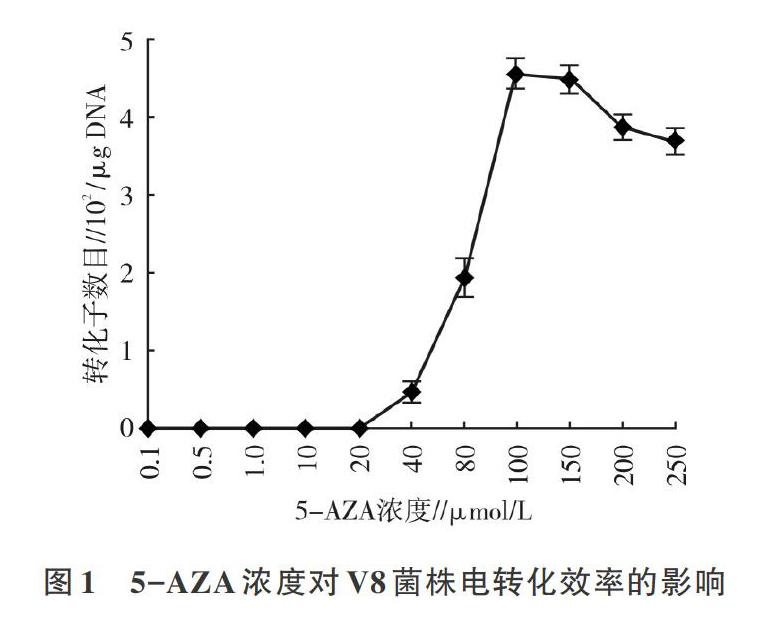
由于在用去甲基化氮胞苷驯化V8菌株之前,不清楚使V8菌株转化效率最高的5-AZA浓度,故配制了l mmol/L的母液,采用逐代驯化的模式,摸索出V8电转化效率与5-AZA浓度的关系。当5-AZA浓度大于20 μmol/L时,开始出现转化子,直至5-AZA浓度达100 μmol/L时,V8菌株的电转化效率相对较高,约为4.5x102/μ DNA,随后继续提高5-AZA的筛选压力,V8菌株的转化效率有所下降,说明100 μmol/L左右的5-AZA筛选的限制修饰系统突变菌株V8比较适合进行遗传转化(图1)。
2. 电转化中各因素对V8电转化效率的影响
由“2.1”试验结果可知,选取去甲基化氮胞苷处理菌株的方式,完成对V8的驯化,以此处理的菌株作为制备后续电转化的感受态细胞,并以此对电转化中涉及的各因素进行逐个研究。
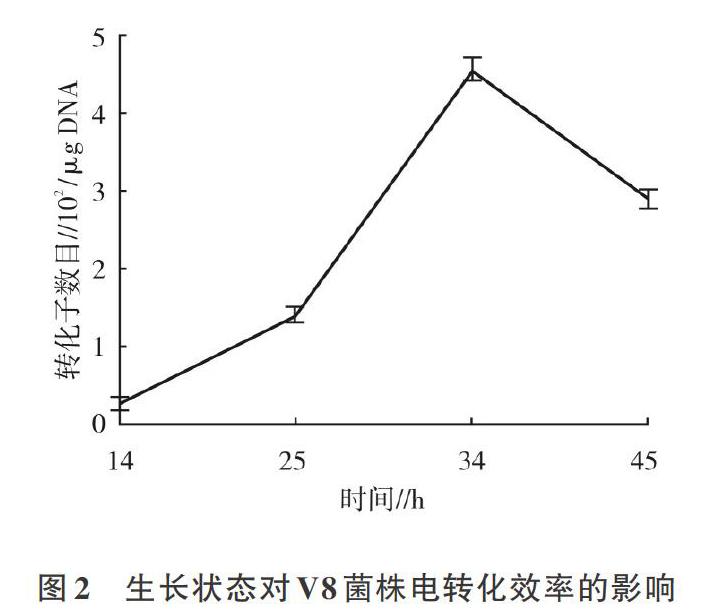
2.2.1 细菌生长状态对V8菌株电转化效率的影响V8培养到34 h左右,电转化效率达到最高,此时对应的菌液OD600mm为0.6左右,说明对数中期的V8生长旺盛,种群数量多,菌体粗壮,适合制备用于电转化的感受态细胞(图2)。
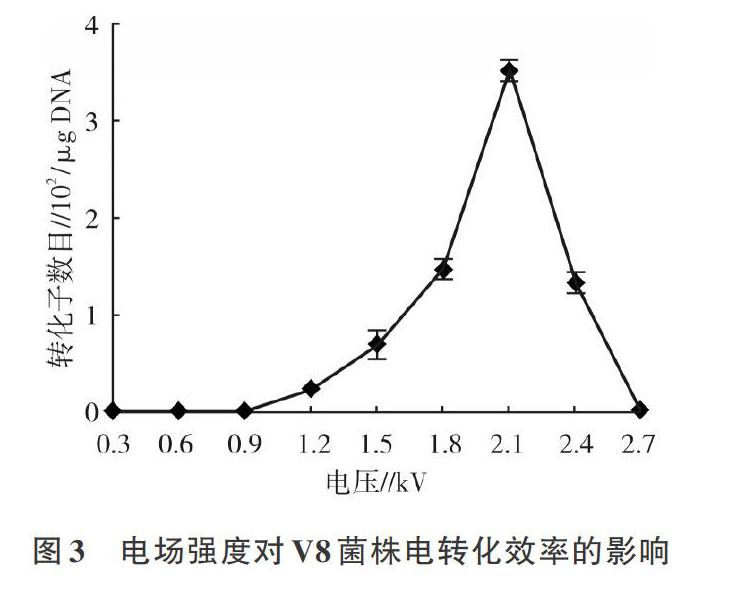
2.2.2 电场强度对V8电转化效率的影响 在电压小于2.1 kV时,电压与V8的电转化效率呈正相关,此时V8对外源质粒的吸收能力随电压强度增强而变大;但当电压超过2.1 kV后,二者之间的关系由正相关转变为负相关,甚至在电压达2.7 kV时,由于过高的电压会导致感受态细胞大量死亡,导致电转化效率降为0。说明2.1 kV是适合V8电转化的最佳电压,此时电转化效率达3.5x102/μgDNA(图3)。
2.2.3 质粒浓度对V8电转化效率的影响 质粒浓度与电转化效率的关系呈正相关,但无限制的提高质粒浓度,会出现电转化杯的“爆杯”,因此,为保证试验安全,质粒浓度维持在(50+10) μg/mL,可保证V8电转化效率约2.4x102/μg DNA(图4)。
2.2.4质粒用量对V8电转化效率的影响 在质粒用量小于或等于7μL时,质粒量与电转化效率呈正相关,随着质粒量的提高,电转化效率逐渐升高;但当质粒量超过7μL时,二者之间的关系转变为负相关。表明7 μL的质粒即可以使V8获得3.1x102/μg DNA的电转化效率(图5)。
2.2.5复苏培养基种类对V8电转化效率的影响SOC作为复苏培养基时,V8电转化效率最高,其次是NA,二者作为复苏培养基的电转化效率是LB的200倍左右。说明SOC和NA相对适合作为V8电转化复苏培养基(图6)。
2.2.6复苏培养基温度对V8菌株电转化效率的影响 4℃预冷SOC的转化效率约是常温复苏培养基电转化效率的1.5倍。说明预冷的SOC是V8电转化时适合受伤感受态细胞恢复活力的培养基(图7)。
2.2.7复苏方式对V8电转化效率的影响 采用在电击结束后对感受态细胞不复苏培养直接涂板、3h的静置复苏培养和振荡复苏培养的3种复苏方式,不同复苏方式使V8电转化效率差异明显。由图8可以看出,感受态细胞不复苏培养后电转化效率为0,感受态细胞静置复苏培养后也几乎没有转化子的产生。相反,振荡复苏培养感受态细胞可使V8电转化效率提高350倍左右。说明振荡培养使V8受伤的细胞在有限时间内尽可能地接触复苏培养液,通过降低细胞死亡率而提高电转化效率。
2.2.8复苏时间对V8电转化效率的影响5h是感受态细胞复苏时间与转化效率关系的分界点,在此之前V8的电转化效率与复苏时间呈正相关,转化效率高达4.Ox102/μg DNA;过长的复苏时间可能伴随细菌细胞生长活力的下降,电转化效率并未提高,V8的复苏时间维持在4-5 h比较适宜(图9)。
3 小结与讨论
去甲基化氮胞苷(5-AZA)驯化菌株的方法已在黄单胞菌PX099A上应用成功[5],研究表明V8经过去甲基化氮胞苷驯化后,获得抗100 μmol/L 5-AZA的限制性修饰系统突变菌株,电转化效率最高约为4.5x102/μ DNA,但5-AZA的浓度不能无限制增加,过高的浓度可能会破坏细胞的某些组织甚至因细胞死亡而导致电转化效率的下降[8]。
SOC作为复苏培养基时,电转化效率会相对较高,可达2.9x102/μg DNA,可能SOC含的氯化镁在一定程度上减小了黄单胞菌表面粘多糖的干扰[9]。适宜时间的复苏培养能够使受到电击穿孔的受损细胞壁发生愈合,促使受损的细胞进行细胞分裂,增大电转化效率,但是过长的复苏时间反而因为细胞分裂产生的一些有毒物质而降低电转化效率[10],因此V8最佳的复苏时间应该维持在4.5 h左右。
通过对电转化一系列因素的摸索,得到了V8菌株电转化的最优条件,为V8的遗传操作提供了一种方便、快捷的途径。前期通过细菌本身的无细胞胞体外甲基化待转质粒,也成功打破了V8电转化效率为零的限制,在保证V8和重组质粒双重驯化的前提下,进行电转操作时得到的最高电转化效率和最优电转化因素是否会不同,有待后续进一步研究。
参考文献:
[1]张明月.棉花角斑病菌由ClpA_ATPase介导的逆境胁迫相关基因的功能[D].河南开封:河南大学,2017.
[2]缪卫国.转hpalXoo基因棉花抗病虫防卫反应与全基因组转录谱分析及棉花角斑病菌hpaXm基因的功能[D].南京:南京农业大学,2009.164-167.
[3] XU B,ZHANC Z.GE Q,et al.A study on classification of Xan-thomonas by isoeleclric focusing IJl.Wei sheng wu xue bao= Actamicrobiologica sinica. 1993 .33(1):7-12.
[4]张鼎鼎,邹丽芳,赵梅勤,等.hrc0基因决定水稻条斑病菌在非寄主烟草上的过敏性反应和在寄主水稻上的致病性[J].中国水稻科学,2011,25(1):11-18.
[5]李 岗中国水稻白叶枯菌致病多样性和遗传多样性分析与avrBs3/pthA家族新成员介导的表型研究[D]南京:南京农业大学.2008.122-123.
[6]王清,梁鹏,李响,等.棉花角斑病菌遗传驯化体系的建立[J].江苏农业科学,2016,44(3):153-157.
[7]李鹏伟.哈氏噬纤维菌电转化条件的优化及表达体系的构建[D].济南:山东大学,2012.
[8]曲露露,冯献启,赵洪国,等.不同浓度的5一氮杂胞苷对RP-MI8226细胞系的诱导凋亡作用分析[J].现代生物医学进展,2013. 13( 13): 2437-2439.2444.
[9]吴东方副猪嗜血杆菌突变系统的初步研究[D].武汉:华中农业大学,2010.23-25.
[10]李爽.D-乳酸产生菌株的基因敲除[D].天津:天津大学,2010.13-14.
基金项目:国家甘薯产业技术体系项目( CARS-IO-C-13);国家重大基础研究计划项目(20IICB111612);国家农业产业技术体系建设项目(CARS-34-GW8)
作者简介:王清(1988-),女,河南项城人,实习研究员,硕士,主要从事植物保护等工作,(电话)15890213237(电子信箱)1173823088@qq.com;
通信作者,劉志坚(1966-),男,河南漯河人,研究员,主要从事甘薯品种选育及利用工作,(电话)0395-3781568(电子信箱)liuzj66@126.com。
