论植物杂种优势的研究途径
2020-07-27潘春桂

摘 要:植物群体叶面积指数与平均单位叶面积截获光能的反相关导致群体光能截获杂种优势形成的必然性,各种养分环境消耗进入临界点同步性改善导致养分获取杂种优势形成的必然性。植株质量或作物产量的杂种优势主要由双亲群体光能截获和各种养分获取的特征及差异所决定。遗传与环境的相互作用决定了杂种优势的形成过程。动态跟踪植株群体光照结构、植株质量或产量形成过程、根际环境中各种养分的浓度等因子有助于理解杂种优势形成过程,单独研究双亲遗传差异不能获得杂种优势形成的一般机制。
关键词:光能;养分;反相关;同步性;必然性;杂种优势
Abstract:The inverse correlation between the leaf area index of plant population and the light energy intercepted per unit leaf area leads to the inevitability of the formation of heterosis of population light energy intercepted, and the improvement of synchronicity of various nutrient environmental consumption into the critical point leads to the inevitability of the formation of heterosis of nutrient obtained. The heterosis of plant weight or crop yield is mainly determined by the characteristics and differences of light interception and various nutrients acquisition of parents. The interaction between heredity and environment determines the formation of heterosis. Dynamic tracking of light structure, plant weight or yield formation process, concentration of various nutrients in rhizosphere environment and other factors is helpful to understand the formation process of heterosis. The general mechanism of heterosis formation can not be obtained by studying the genetic difference between parents in isolation.
Key words: light energy; nutrient; inverse correlation; syncronicity heterosis; inevitability; heterosis
从达尔文时代就已经认识到杂种优势现象,人类利用杂种优势增加经济效益以及研究杂种优势的形成机理也有近一个世纪。对其机理的解释,经典遗传学提出了显性、超显性、上位性效应假说,分子遗传学认为基因差异表达、表观遗传修饰包括DNA甲基化和组蛋白修饰参与杂种优势的形成[1],以上假说也获得了相关试验的支持。另外,一些学者也提出了遗传平衡、活性基因遗传振动活成、基因网络系统及自组织理论等假说[2],不难看出,以上假说共同性在于将杂种一代内在相关过程的特异性等价为杂种优势的形成机理或产生原因,实际上它们仍然为杂种优势的表现或结果,就像杂种一代营养体较大,生物或经济产量较高一样,不能作为杂种优势的形成原因。可能这些研究潜在地认为亲本两套遗传物质相互作用就产生了杂种优势,排斥或弱化了环境因素在杂种优势形成中的地位。相同遗传类型在不同环境中具有生理、生化、形态过程的差异性,表明环境效应从遗传表达到形态建立各个环节中均产生重要影响。假设各基因位点加性表达,在外部环境作用之下,这种遗传过程是不是能够转变呢?从能量平衡而言,杂种一代的优势应该与获得更多的光能和养分有关。本研究从上述基本假设出发,论述杂种优势形成机理可能的研究途径。
1 光能截获的杂种优势
如果一个乘积性状等于两个单因子性状的乘积,两个单因子大小在双亲间反向变化,并假设它们均为加性遗传,则乘积性状具有正向中亲优势或杂种优势;单因子差异越大,杂种优势越大。类似地,如果两个单因子在双亲间同向变化,则存在负向杂种优势,两个单因子差异越大,杂种优势越小[3]。当至少有一个单因子双亲相等时,杂种优势为零;任一单因子双亲均乘以相同系数时,不改变杂种优势大小。由于加性遗传仅是理论上成立的,以上均为理论杂种优势。在实际情形中,如果以上条件近似满足时,结论也是适用的。
植物群体光能利用率随群体密度而变化,当群体密度较小或叶面积指数较小时,群体冠层光能反射率较低,而透光率较高,各层叶片均可获得较好的光照。当群体叶面积指数过大时,群体冠层光能反射率较高,而透光率下降,上部叶片经常处在饱和点之上,下部叶片可处在光补偿点之下,平均单位叶面积截获光能下降。这是一个必然的过程,群体叶面积指数越大,叶片或叶层互相遮掩越严重。在密度相同的情况下,以单株叶面积代替群体叶面积,单株截获光能等于单株叶面积与单位叶面积截获光能的乘积,可见这个乘积性状两个构成单因子具有反相关倾向。由于单位叶面积截获光能和单株叶面积、叶片大小、形状等有关,如果认为单株叶面积和单位叶面积截获光能均近似为加性遗传,则从上文分析可知,单株截获光能的杂种优势是一个本能过程[3]。下文从一些引文数据的计算结果来分析光能截获和产量形成杂种优势之间关系。司洪华[4]的15个玉米杂交种试验表明,当以单株叶面积与单位叶面积籽粒产量的乘积来模拟上述单株光能截取过程时,光能截获理论杂种优势与实际籽粒产量杂种优势相关系数为0.523 3(P<0.05)。吴才君等[5]的6个芸薹种蔬菜交种试验表明,当以单株叶面积与单位叶面积株重的乘积来模拟光能截获时,光能截获理论杂种优势与单株重实际杂种优势相关系数为0.843 4(P<0.05),原文结果亦表明,利用CDNA-AFLP技术研究双亲与杂交种的基因差异表达,未发现任何一种表达类型比例与株重杂种优势有显著相关性。由于植株体单株分枝数与单株叶面积存在线性关系,当以单株分枝数与单个分枝的产量的乘积来模拟单株光合产物时,赵禹凯等[6]的18个桔梗杂交种试验表时,光合产物理论杂种优势与桔梗果实数实际杂种优势相关系数为0.471 3(P<0.05)。余政军等[7]棉花5×6完全双列杂交试验表明,这里以正反交平均产量表示组合产量,当以单株果枝数与单个果枝皮棉产量的乘积来模拟光合产物时,光合产物理论杂种优势与实际皮棉产量杂种优势相关系数为0.526 4(P<0.05);杨祥波[8]30个大豆杂交种试验表明,当以单株分枝数与单个分枝的籽粒产量乘积来模拟光合作物时,光合产物理论杂种优势与产量实际杂种优势相关系数为0.860 2(P<0.001),其研究结果显示,SSP标记的分子距离与单株粒质量杂种优势的相关系数为-0.42(P<0.05)。李素萍[9]的试验数据显示,如果构建一个乘积性状,向日葵单株粒质量=(单株叶片数×生育期)×[单株粒质量/(单株叶片数×生育期)],亲本间以上两个乘积因子相关系数为-0.672(P<0.05),单株粒质量理论杂种优势与实际杂种优势相关系数为0.443 6(P<0.05),以上结果并不总是出现的,因为没有考虑到产量形成过程或叶片功能期长短等的遗传差异,虽然如此,以上结果也足够提示了光能截获这个乘积性状的深入研究对于理解产量杂种优势机制的重要性。以上光合产物或产量这个乘积性状2个构成单因子均直接与光能截獲有关,由于可有多种方法将光合产物或产量分解为2个单因子乘积,例如棉花皮棉产量可以为籽棉产量与衣分的乘积,也可以分解总成铃数与单铃质量的乘积;水稻产量可以为总穗数与单穗质量的乘积,也可以为总粒数与粒质量的乘积。然后按照单因子加性遗传来计算乘积性状理论杂种优势,这与上文从光能截获角度来研究杂种优势是否有同等效果呢?这值得进一步研究。
2 养分获取的杂种优势
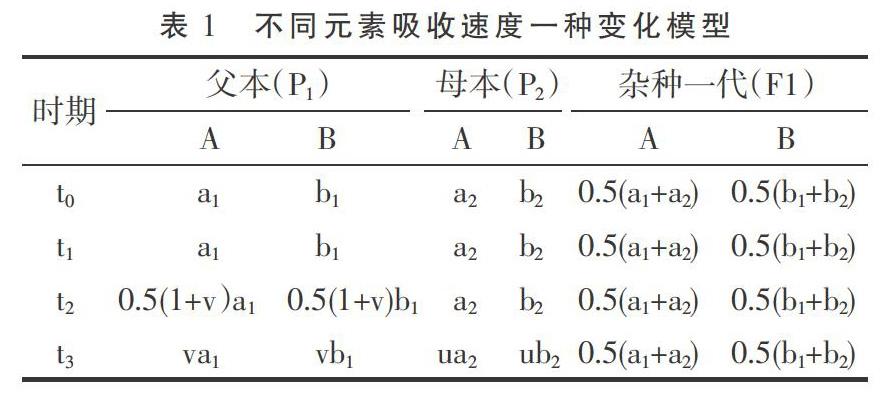
杂种优势的形成也与吸收更多的营养元素有关。植株体对外部环境中养分吸收速度与遗传、环境养分浓度及植株体大小等有关,可用Logistic曲线来模拟植株生长过程[10-11]。可近似认为,在一定条件之下,(1)植株体对各种营养元素的吸收存在饱和浓度或饱和期,此时增加任何一种养分的浓度或环境供给,植株体对各种元素的吸收速度不再增加[12],这里假定不考虑盐害浓度;(2)在饱和期杂种一代对各种元素的吸收速度为加性遗传;(3)植株体对各种元素吸收速度为固定比例,只与遗传类型有关;(4)各种营养元素均存在临界环境浓度或临界点,对某种元素而言,此时减少其环境浓度,植株体对这种元素的吸收速度会下降,其他元素也协同下降,但吸收速度仍保持固定比例。基于以上假设,如果一些元素饱和期吸收速度在双亲间存在差异,则最大元素吸收速度被双亲所具有,因此,经过一段生长时间后,双亲均因为各自具有最大元素吸收速度可能比杂种一代更早地通过临界点,其后杂种一代对各种元素吸收速度便超过中亲值,因此,各器官或单株干质量的杂种优势便开始形成。
根据上文假设条件,易得2F1/(p1+p2)>1,即此时段已形成正向中亲优势或杂种优势。当然,对于某一种元素而言,不同生长时期饱和吸收速度存在差异,临界环境浓度也不同,以上分析假设t0-t3时间较短或各种养分饱和吸收速度不变。
以上述养分吸收模型可产生如下推论。(1)在饱和期,理论上是不存在杂种优势的;或者说,在植株生长很早时期和环境养分供给极端充分时,杂种优势平均值较小或趋于零,变化也较小。可能这个时期很短,以致不易被分辨出来。(2)如果生长时间足够长或各种养分已进入相对胁迫期,植株体养分吸收与环境供给相容性好易获得较大的质量(鲜质量或干质量,下文同),相容性与质量呈正向关系。相容性好指各种养分有较大的饱和吸收速度、和环境供给有较好的协调性即各种养分进入临界点时间相近以及各种养分有较大的(临界)环境消耗率等。由于以相同的外部环境作共同参照,双亲质量越大,则遗传相似性越好;双亲质量越小,由于存在各类不相容性,双亲总体上有较大的遗传差异,但不同组合间可有一定变化。环境相容性好的双亲从通过临界点到其后延续时间里,各种营养元素吸收速度的衰减(相对于饱和吸收速度衰减,下文同)和杂种一代相近,衰减期也和杂种一代较同步,因此不易获得较大的杂种优势;环境相容性差且遗传差异较大时,可导致双亲间各养分平均衰减幅度更大,平均衰减时间更早,易形成较大的杂种优势。因此在大多数情况下,双亲质量之和与杂种优势易呈显著的负相关。(3)植株体质量的变化与饱和养分吸收速度及其衰减均有关。如果前期一直处于饱和期,在临界期附近;或植株生长很早时期和养分供给相对充足条件下,植株质量主要与各种养分尤其是大量元素饱和吸收速度有关,双亲质量之差可一定程度上反映遗传差异;其较大时,双亲之一或双亲养分吸收速度在后续时间内有较大的衰减,衰减时间也较早;因此,此时刻双亲质量之差与邻近较短时段内植株体质量增加量杂种优势可呈正相关。(4)由于植株体质量取样时间、试验环境、遗传背景的巧合,质量较小的亲本或因饱和期养分吸收速度慢未通过或较迟通过临界点,或因饱和期养分吸收速度慢通过临界点后衰减也慢,也可能它们遗传相似性好;质量较大的亲本则相反。但通过临界点之后延续时间均不长,饱和期养分吸收速度主要决定了植株体质量。这时双亲质量之和与杂种优势可呈弱正相关或正相关。但不排除一些不同试验条件下可产生相似的遗传过程。下文通过文献数据的计算结果来证明以上推论。马谦[13]的试验数据表明,大田条件下水稻播种后第20天的幼苗干质量杂种优势为-5.8±19.4(%),而第30天为23.41±19.1(%),第20天到第30天幼苗干质量增加量杂种优势与第20天双亲干质量之差绝对值呈正相关,相关系数为0.379 9(P<0.05),与第30天干质量双亲之和呈负相关,相关系数为-0.527 2(P<0.005)。另外,第20天幼苗干质量杂种优势与同时刻双亲干质量之和相关系数为-0.044 5,第30天幼苗干质量的杂种优势与此时双亲之和呈弱负相关,相关系数为-0.323 7(P<0.1)。以上结果表明,第20天时几乎所有亲本仍处于饱和期或在临界点附近。第30天时,所有亲本均已通过临界期,但其后延续时间均较短或各种养分环境胁迫仍不严重。马谦[13]还有一个黑暗条件之下水稻种子发芽4天后幼芽干质量数据,幼芽干质量的杂种优势与双亲之和呈负相关,相关系数为-0.677 8(P<0.05),杂种一代幼芽干质量和中亲值之差(离亲值,下文同)与中亲值相关系数为-0.603 4(P<0.1),由于发芽时种子贮藏的干物质是唯一能量来源,说明第4天时,各种营养元素已存在明显的胁迫。余丽霞[14]棉花杂交种愈伤组织培养试验数据计算表明,愈伤组织质量杂种优势与同时刻双亲之和的相关系数在第35天、第65天、第80天分别为-0.597 4(P<0.002)、-0.675 0(P<0.001)、及-0.696 5(P<0.001);杂种一代愈伤组织重量离亲值与同时刻中亲值相关系数在第35天、第65天及第80天分别-0.601 0(P<0.002)、-0.670 0(P<0.001)、-0.700 6(P<0.001);第35天到第65天愈伤组织质量增加量杂种优势与第35天、第65天、第80天双亲质量之和相关系数分别为-0.508 2(P<0.01)、-0.714 1(P<0.001)、-0.708 9(P<0.001),说明在第35天时,各种养分早已进入协迫阶段。第65天到第80天愈伤组织质量增加量杂种优势与第35天、第65天、第80天双亲质量之和相关系数分别为-0.544 2(P<0.005)、-0.533 1(P<0.01)、-0.590 3(P<0.002),可能生长后期外界养分已较严重亏缺,杂种一代对各种养分吸收速度也衰减严重,杂种优势变得较不活跃,双亲遗传差异大小与愈伤组织增加量杂种优势相关性减弱。王章奎[15]营养液栽培小麦试验表明,拔节期单株根系干质量杂种优势与双亲之和相关系数为-0.750 5(P<0.001),杂种一代单株根系干重离亲值与中亲值相关系数为-0.704 9(P<0.001),说明这时营养液各种养分的供给已进入较重的胁迫阶段。温玲[16]甜瓜杂交种试验表明,4叶一心期单株根系干质量杂种优势与双亲之和相关系数为0.743 5(P<0.05),这与上文推论第(4)点有关。同时也表明取样时各种养分的环境供给可能仍比较充裕。蔡丽艳等[17]苜蓿杂交种数据也可能属于同样性质,苜蓿干草单株质量的杂种优势与双亲之和相关系数为0.388 3(P<0.05)。大田作物经过一个生长季节,土壤养分已处于相对亏缺状态,这时作物产量杂种优势与双亲之和可为显著或极显著负相关。林欣立等[18]萝卜杂交试验表明,单根质量杂种优势与双亲之和相关系数为-0.583 8(P<0.05)。潘春桂[19]计算结果表明,皮棉产量杂种优势与双亲之和相关系数为-0.696 6(P<0.001)。徐静斐和汪路应[20]的水稻试验表明,水稻产量的杂种优势与双亲之和相关系数为-0.523 4(P<0.002)。司洪华[4]玉米杂交试验表明,玉米产量杂种优势与双亲之和相关系数为-0.840 3(P<0.001)。吴利民等[21]小麦杂种试验,小麦产量杂种优势与双亲之和相关系数为-0.483 3(P<0.05)。梁耀平等[22]西瓜杂交试验表明,西瓜产量杂种优势与双亲之和相关系数为-0.715 7(P<0.001)。阎冬生[23]大豆杂交试验表明,大豆产量杂种优势与双亲之和相关系数为-0.848 5(P<0.001)。桑世飞等[24]的油菜杂交试验表明,油菜产量杂种优势与产量双亲之和相关系数为 -0.523 4(P<0.001)。由于杂种优势与双亲之和相关性受到自相关的影响,进一步计算出杂种一代产量离亲值与中亲值的相关性,绝大多数仍呈显著或极显著负相关,只有林欣立、司洪华的试验中两者呈弱负相关,这里未列出相关系数。另外,上文已经提及,双亲之和越小时,双亲遗传差异在不同组合间可变化较大。因此,从线性相关的计算原理可知,由于亲本选配的随意性,有些情况下去除双亲之和最小的一些组合,可显著提高杂种一代离亲值或杂种优势与中亲值相关性。当然与以下原因有关。如果杂交试验中存在很多如下类型组合,因各种养分的吸收速度或(临界)环境消耗率较小导致双亲质量(之和)越小,杂种一代质量由于各种养分进入临界点同步性改善而获得的离亲值反向同步增大受到限制,因此离亲值与中亲值或双亲之和的反相关性减弱或不存在。这在公开发表的文献中十分常见。例如桑世飞等[24]油菜杂交试验原有组合46个,去除产量双亲之和最小的6个组合,杂种一代产量离亲值与中亲值相关系数由原-0.454 8(P<0.002)变为-0.564 3(P<0.005)。王建军等[25]油菜杂交试验原有杂交组合36个,去除籽粒产量双亲之和最小的2个组合,籽粒产量离亲值与双亲之和相关系数由原-0.326 3(P<0.1)增加为-0.474 3(P<0.005)。唐梅[26]水稻杂交试验重庆试点原有组合35个,现去除单株粒质量双亲之和最小的8個组合,离亲值与双亲之和的相关系数由原-0.183 0增加为-0.539 4(P<0.005)。由于植株体主要由大量元素构成,但它对大量元素和少(微)量元素的获取是相互协同和相互制约的关系,因试验环境、遗传背景、目标性状及取样时间不同,杂种优势与亲本生物体质量的相关性存在可解释的变化,但在生长后期或环境养分相对亏缺时,负相关性几乎是必然的,有兴趣的读者可以去验证。
3 结论与讨论
光能截获与环境养分获取两种模型在解释杂种优势形成时可存在兼容性。例于在某种条件下大田作物生长早期,群体叶面积指数很小,单株叶面积与单位叶面积截获光能在双亲间不易形成反向变化关系,甚至由于叶片形态等的差异,这两个因子在双亲间呈同向变化。因此,根据前文的结果,这时杂种优势大多数组合为零,甚至出现负向杂种优势。从养分获取模型来说,由于此时大多数亲本仍处于养分饱和期或临界点附近,所以也不易形成正向杂种优势。大田作物要获得很高的产量,必须群体叶面积、叶片功能期、光合效率均表现较好;产量很低时,可允许这3个因子优劣的不同组合。从光能截获模型考虑,双亲产量越高时越不易形成杂种优势;甚至存在一些组合双亲产量之和较大,表现为一个亲本产量很高,另一个产量一般,光能截获构成因子在双亲间同向变化从而形成负向杂种优势。双亲产量减小时,易获得光能截获的反极因子效应[3],形成杂种优势;从养分获得模型考虑,双亲产量越高,养分吸收和环境供给相容性越好,遗传相似性越好,越不易获得杂种优势,正如上文所述。从上文不难看出,光能截获的反极因子效应和养分获取的杂种优势形成机理使植物体整体或部分质量存在正向中亲优势或杂种优势近似于一种本能或必然的自然现象。这从公开发表的各种作物杂交试验的相关文献中能够发现[3-9,18-26],几乎所有杂交试验中,各组合中亲优势优势平均值大于零或者正向中亲优势的组合比例大于50%,占很大比例的试验中均接近100%。但对于多年生乔本植物而言,杂种一代早期群体光能截获的杂种优势不能适时地转换为生殖器官相关性积累,绝大部分用于加快群体营养生长或叶面积指数增加,以致群体过于繁茂引起的平均单位叶面积截获光能下降或低于双亲,经过较长时间或数年后,植株营养生长量例于主茎高度及茎围、材积等性状正向中亲优势或杂种优势获得的必然性显著下降,各组合杂种优势平均值或趋于零[27-28]。目前,对于杂种优势产生机理的各种解释,如显性、超显性、上位性效应、基因网络、遗传振动等假设,都未能触及杂种优势的必然性,更由于它们所涉及的遗传方式均具有随机性,也不能形成杂种优势的必然性。与目前所有研究与杂种优势相关性的研究方法相比[4-5,8,15-16,21-26]本文所揭示的杂种优势与有关因子的相关性显著度大幅度提高,而且是恒定的,或是可解释的变化。这是因为本文所研究的相关性是揭示杂种优势內在机理的必然相关性,而不是在以往研究中从寻求相关性过程中去试图了解杂种优势形成机理,是两个相反的过程。本文研究不设定遗传背景和试验环境,强化环境效应在杂种优势形成中的地位,避免了以往研究当试验条件改变时结论不再适用的情况。很显然,环境相容性包括养分的供给数量、空间分布及供给时期等方面,还包括温度、光照、病害、土壤湿度等方面。笔者认为,本文的研究方法或是杂种优势形成机理终极揭秘一个全新的重要视角。
参考文献:
[1]张义荣,姚颖垠,彭惠茹,等.植物杂种优势形成的分子遗传机理研究进展[J].自然科学通讯,2009,19(7):697-703.
[2]何光明,邓兴旺.植物杂种优势分子机理研究:机遇和挑战[J].中国基础科学植物科学专刊,2016(1):28-34.
[3]潘春桂.基于反极因子的杂种优势形成途径[J]. 陕西农业科学,2018,64(4): 71-73,76.
[4]司洪华 .利用玉米杂交当代杂种优势预测成株期杂种优势的研究[D].沈阳:沈阳农业大学, 2006.
[5]吴才君,曹家树,董德坤,等. 芸薹种蔬菜杂交种及其亲本莲座期基因差异表达与杂种优势的关系[J].中国农业科学,2004,37(11):1654-1659.
[6]赵禹凯,魏建和,杨成民,等.桔梗主要农艺性状的杂种优势分析[J]. 中药材,2010, 33(10):1523-1527.
[7]余政军,邵小强,肖新宇,等.棉花主要性状的遗传相关性分析[J].湖南农业科学,2014(1):17-20.
[8]杨祥波. SSR分子标记遗传距离预测大豆亲本杂种优势的初步研究[D].长春:吉林农业大学,2008.
[9]李素萍.食用向日葵杂种优势及配合力研究[D].呼和浩特:内蒙古农业大学,2006.
[10]李秋元,孟德顺. Logistic曲线的性质及其在植物生长分析中的应用[J].西北林学院学报,1998,8(3): 81-86.
[11]莫红,陈瑛.基于Logistic方程的植物生长过程模型与最优分析[J].焦作大学学报, 2006(4):70-71.
[12]罗林会.不同配方水培营养液对皱叶留兰香生长发育的影响[J].江苏农业科学, 2009(3):227-228.
[13]马谦.水稻苗期杂种优势分析及赤霉素与苗期杂种优势生物学基础的关系研究[D].武汉:华中农业大学,2011.
[14]余丽霞.陆地棉主要经济性状的遗传及杂种优势研究[D].武汉:华中农业大学,2005.
[15]王章奎 .普通小麦杂种与亲本间根系基因表达差异与杂种优势分子机理[D].北京:中国农业大学, 2004.
[16]温玲. 甜瓜杂种优势早期预测指标的筛选[D]. 哈尔滨:东北农业大学,2003.
[17]蔡丽艳,石凤翎,陈海玲.不同苜蓿雄性不育杂交组合牧草产量杂种优势分析[J].中国草地学报,2013,35(2):24-30.
[18]林欣立,李寅安,王美菊.萝卜杂种优势与配合力的研究[J].中国农业科学,1986,19(20):54-60.
[19]潘春桂. 一种提高作物产量杂种优势的亲本选配方法—以棉花为例[J].安徽农业科学,2017,45(7):28-29,99.
[20]徐静斐,汪路应.水稻杂种优势与配合力的初步研究[EB/0L].安徽农学院学报, http://www.cnki.com.cn/Article/CJFD
Total-ANHU197900002.htm.
[21]吴利民,倪中福,王章奎,等. 小麦杂种及其亲本苗期叶片家族基因差异表达及其与杂种优势关系的初步研究[J]. 遗传学报,2001,28(3):256-266.
[22]梁耀平,王世杰,陈豫梅,等.西瓜亲本遗传差异与杂种优势的相关性[J].江苏农业学报,2012, 28(3):622-626.
[23]阎冬生. 大豆籽粒基因差异表达模式与杂种优势关系的研究[D].长春:吉林大学,2008.
[24]桑世飞,王会,梅德圣,等.利用全基因组SNP芯片分析油菜遗传距离与杂种优势的关系[J].中国农业科学, 2015,48(12):2469-2478.
[25]王建军,雍洪军,张晓聪,等.12几个外来玉米群体与我国主要种质配合力和杂种优势分析[J].作物学报,2012,38(2):2170-2177.
[26]唐梅.中籼杂交水稻杂种优势的分子际记研究[D].重庆:西南大学,2000.
[27]张一,周志春,金国庆,等.马尾松双列杂交亲本遗传距离与杂种优势相关性分析[J]南京林业大学学报, 2010,34(1):9-14.
[28]王赵民,陈益泰.杉木主要性状配合力分析及杂种优势利用[J].林业科学研究, 1998,11(6):614-624.
