抑制剂T0070907 对鸡PPARγ基因表达影响的研究
2020-06-01王宇祥翟桂影庞永佳孙宇航杜智恒
王宇祥,李 超,翟桂影,庞永佳,李 瑞,孙宇航,杜智恒
(1. 东北农业大学 动物科学技术学院,黑龙江 哈尔滨 150030;2. 农业农村部鸡遗传育种重点实验室,黑龙江 哈尔滨 150030;3. 黑龙江省普通高等学校动物遗传育种与繁殖重点实验室,黑龙江 哈尔滨 150030)
过氧化物酶体增殖物激活受体(Peroxisome proliferator-activated receptor,PPAR)是一类配体激活的核转录因子超家族成员。PPAR 包括PPARα、PPARβ和PPARγ 3 种表型,其中以PPARγ的研究最为深入[1]。PPARγ 主要在脂肪组织,血管内皮细胞、平滑肌细胞以及单核巨噬细胞中表达。PPARγ可以调节参与能量储存和分解相关基因的表达[2],并在脂肪细胞分化、脂肪生成、胰岛素抵抗、炎症反应、血管生成和动脉粥样硬化以及葡萄糖利用的调节中起到关键作用[3]。最近,PPARγ被发现在免疫炎症调控中发挥重要作用。PPARγ可以通过抑制自身反应性T 细胞、减少自身抗体的产生、调节T 细胞的功能及抑制炎症介质的表达来缓解自身免疫性疾病对机体的损害[4];PPARγ 还可通过调控细胞内多条炎症信号传导途径,抑制细胞因子、趋化因子、黏附分子等的基因表达,从而发挥抗炎作用[5-6];此外,激活PPARγ还有利于诱导免疫细胞凋亡和中性粒细胞麻痹[7-8]。可见开展PPARγ的功能研究对于预防和治疗炎症相关疾病具有重要的意义。
在研究基因功能的过程中,往往需要下调或抑制基因的表达,并观察由此引起的细胞或组织表型的变化,从而推测其影响机体生长发育的可能作用[9]。T0070907 是一种新型合成的PPARγ基因抑制剂,化学式为C12H8CIN3O3。众多研究表明,T0070907 是有效的PPARγ拮抗剂,对小鼠PPARγ基因具有较强的亲和力和较高的选择性,可以通过影响PPARγ基因的配体结合域第12 螺旋的构象,调节PPARγ与共激活因子的相互作用而抑制PPARγ的活化[10-11]。其对PPARγ基因的偏好超过对PPARα基因和PPARβ基因的偏好,T0070907 对PPARγ活性的抑制作用已在小鼠胰腺细胞的报告基因和功能检测中得到证实[12],目前已成为研究PPARγ基因功能的主要工具。然而,本实验室在前期针对鸡PPARγ基因的功能研究中偶然发现,T0070907 非但不能抑制鸡PPARγ的活性,反而对该基因的表达具有一定的促进作用。为验证这一现象的真实性,本研究利用荧光定量PCR、western blot 和报告基因技术,分别从mRNA 表达水平、蛋白表达水平及蛋白活性等方面检测了抑制剂T0070907 对DF-1 细胞中鸡PPARγ基因两种亚型[13]表达的影响。以期为选择合适的PPARγ抑制剂,开展该基因的功能研究奠定基础。
1 材料与方法
1.1 主要试剂 T0070907 购自MCE(Med Chem Express)公 司;TRIzol 购 自Invitrogen 公 司;FastStart Universal SYBR Green Master(Rox)购自Roche 公司;DMEM/F12 培养基、胎牛血清、胰蛋白酶和罗格列酮(Rosi)购 自Gibco 公 司;Lipofectamine®2000 购 自Invitrogen 公司;双荧光素酶报告基因检测试剂盒和反转录试剂盒购自Promega 公司;RIPA 缓冲液、辣根过氧化物酶(HRP)标记山羊抗小鼠IgG、ECL 化学发光检测试剂盒、鼠源β-actin 抗体均购自碧云天生物技术有限公司;鼠源HA 抗体,购自北京中杉金桥生物技术有限公司。
1.2 载体与细胞 PPARγ真核表达质粒、过氧化物酶体增殖物反应元件(Peroxisome proliferator response element,PPRE)报告基因质粒、报告基因活性对照质粒pRL-TK、DF-l 细胞为本实验室保存。PPARγ真核表达质粒、PPRE 报告基因质粒构建方法简述如下:根据鸡PPARγ基因序列分别设计引物(序列见表1),扩增鸡PPARγ1、PPARγ2 基因全长编码区;以鸡脂肪组织cDNA 为模板进行PCR 扩增,扩增产物酶切后与pCMV-HA 载体连接,构建成pCMV-HA-PPARγ1、pCMV-HA-PPARγ2 真 核 表 达 质粒。根据PPRE(AGGTCAAAGGTCA)和pRL-TK 质粒(Promega 公司)的相关序列,人工合成一段同时包含3 个PPRE 拷贝和TK 启动子的DNA 片段,使PPRE 序列位于TK 启动子的上游,并在DNA 片段5'和3'端分别引入SacI 和HindIII 酶切位点,然后将该序列与经过相同酶切的pGL3-Basic 载体相连,构建pGL3-3×PPRE 报告基因质粒。所有的表达质粒均经双酶切、测序鉴定后用于后续研究。
1.3 抑制剂T0070907 对PPARγ基因mRNA 转录水平的影响检测 按TRIzol 说明书提取DF-1 细胞总RNA,反转录为cDNA,以其为模板进行荧光定量PCR 检测T0070907 对DF-1 细胞内源性PPARγ基因及LPL 基因mRNA 转录水平的影响。反应体系为:FastStart Universal SYBR Green Master(Rox)(2×)5 μL,上、下游引物(10 μmol/L)各0.2 μL,cDNA 模板1 μL,ddH2O 3.6 μL,总 体 积10 μL;反 应 条 件:95 ℃10 s,95 ℃5 s,60 ℃34 s,共40 个循环;熔解曲线条件:95 ℃15 s,60 ℃10 min,95 ℃15 s。以鸡NONO 基 因 为 内 参,利 用2-ΔΔCt方 法 将 原 始Ct 值 转 换为相对基因转录水平。引物信息见表1。
1.4 抑制剂T0070907 对PPARγ蛋白表达的影响检 测 将pCMV-HA-PPARγ1、pCMV-HA-PPARγ2真核表达质粒分别转染DF-1 细胞,转染24 h 后,向细胞中分别添加5 μmol/L 的T0070907(处理组)及DMSO(对照组);添加48 h 后将DF-1 细胞用冷PBS洗涤后加入1 mL RIPA 缓冲液裂解细胞,提取细胞总蛋白;煮沸变性后经12.5%的SDS-PAGE 检测总蛋白的表达;以鼠源HA抗体(1∶1 000)和内参β-actin(1∶1 000)为一抗,山羊抗小鼠HRP-IgG(1∶3 000)为二抗室温孵育1 h 后,westen blot 检测T0070907 对PPARγ蛋白表达的影响。

表1 引物信息Table 1 Primers information in this study
1.5 抑制剂T0070907对PPRE报告基因活性的影响检测 将pGL3-3×PPRE 报告基因质粒转染DF-1 细胞,转染24 h 后添加不同浓度(0.1 μmol/L、0.5 μmol/L、1 μmol/L、2 μmol/L、5 μmol/L)的T0070907,24 h 后收集细胞,采用双荧光素酶报告基因检测试剂盒进行荧光素酶活性检测,分析T0070907 对PPRE 报告基因活性的影响;报告基因活性分析共进行3 次生物学重复,每次3 个技术重复,报告基因活性用Fluc/Rluc 比值表示。
1.6 同时添加抑制剂T0070907 和激动剂Rosi 对PPRE报告基因活性的影响检测 将pGL3-3×PPRE报告基因质粒转染DF-1 细胞,24 h 后分别添加20 μmol/L PPARγ特异性激动剂—罗格列酮(Rosi)、5 μmol/L T0070907 以 及20 μmol/L Rosi+5 μmol/L T0070907,24 h 后收集细胞,采用双荧光素酶报告基因检测试剂盒进行荧光素酶活性检测,分析单独添加Rosi、T0070907、共 同 添 加Rosi 和T0070907 对PPRE 报 告基因活性的影响;报告基因活性分析共进行3 次生物学重复,每次3 个技术重复,报告基因活性用Fluc/Rluc 比值表示。
1.7 统计学分析 采用GraphPad Prism 5软件对试验数据进行统计分析,试验数据以“平均值±标准差”表示,采用t 检验对试验结果进行差异显著性分析,p<0.05为差异显著,p<0.01为差异极显著。
2 结 果
2.1 抑制剂T0070907 对PPARγ基因mRNA 转录水平影响的检测结果 采用不同浓度的T0070907 处理DF-1 细胞,利用荧光定量PCR 检测T0070907 对细胞内源性PPARγ基因及LPL 基因转录水平的影响。结果显示,随着T0070907 浓度的增加,细胞内PPARγ基因的转录水平逐渐升高,呈现明显的剂量依赖性;在T0070907 浓度为1 μmol/L 时,处理组细胞内PPARγ 基因的转录水平显著高于对照组(p<0.05),至浓度为5 μmol/L 时两组差异达到极显著水平(p<0.01)(图1A)。但不同浓度的T0070907 对细胞内LPL 基因的转录水平无显著影响(图1B)。表明T0070907 能够特异性地促进DF-1 细胞中PPARγ基因的转录水平。
2.2 抑制剂T0070907 对PPARγ蛋白表达影响的检测结果 将过表达PPARγ1 或PPARγ2 的DF-1 细胞中分别添加不同浓度的T0070907,经western blot检测外源PPARγ蛋白的表达。结果显示,与对照组相比,T0070907 处理细胞后,PPARγ2 蛋白表达量明显增加,而PPARγ1 的蛋白表达水平无明显变化(图2)。表明T0070907 对PPARγ 蛋白表达有一定的促进作用,但仅限于促进PPARγ2 蛋白亚型的表达。

图1 不同浓度T0070907 对DF-1 细胞中PPARγ基因和LPL 基因转录水平的影响Fig.1 Effects of T0070907 at different concentrations on the transcription levels of PPARγ gene and LPL gene in DF-1 cells

图2 抑制剂T0070907 对PPARγ蛋白表达影响的检测结果Fig.2 Effect of T0070907 on expression of PPARγ in DF-1 cells
2.3 抑制剂T0070907 对PPRE 报告基因活性影响的检测结果 将转染pGL3-3×PPRE 报告基因质粒的DF-1 细胞中添加不同浓度的T0070907,经双荧光素酶报告基因技术检测细胞荧光素酶的活性。结果显示,与对照组相比,处理组随着T0070907 浓度的增加,PPRE 报告基因活性逐渐增强,并且在浓度为0.5 μmol/L 时显著高于对照组(p<0.05),浓度为2 μmol/L、5 μmol/L 时两组差异达到极显著水平(p<0.01)(图3A)。表明T0070907 能够显著促进PPRE 介导的荧光素酶的蛋白活性。
2.4 同时添加抑制剂T0070907 和激动剂Rosi 对PPRE 报告基因活性影响的检测结果 向转染pGL3-3×PPRE 报告基因质粒的DF-1 细胞中分别添加Rosi、T0070907 以及二者的混合物,采用双荧光素酶报告基因检测试剂盒检测。结果显示,单独添加T0070907 与Rosi 均可显著增强PPRE 报告基因活性(p<0.05),添加T0070907 并不能降低Rosi 对PPRE报告基因活性的促进作用(图3B)。表明T0070907与Rosi 均可促进PPRE 介导的荧光素酶的蛋白活性,并且T0070907 不能抑制Rosi 对PPRE 介导的荧光素酶蛋白活性的激活作用。
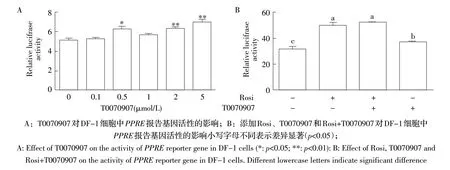
图3 抑制剂T0070907 对PPRE 报告基因活性的影响Fig.3 Effect of inhibitor T0070907 on PPRE reporter gene activity
3 讨 论
LEE 等研究发现,作为一种特异性的、高亲和力的PPARγ拮抗剂,T0070907 在生化和细胞检测中能够阻断PPARγ的活性[10]。T0070907 对PPARγ活性的抑制作用已在小鼠胰腺细胞、人结直肠癌细胞、宫颈癌细胞的功能检测中得以证实[12,14-15],T0070907还可以通过一种PPARγ依赖的方式有效抑制脂肪生成[16]。然而本研究的结果显示,T0070907 对DF-1 细胞中PPARγ的表达无论是mRNA 转录水平还是蛋白水平,以及蛋白活性均具有正向调节作用。
彭璇等研究显示,在猪血管上皮细胞中,随着T0070907 浓度的增加,PPARγ基因的表达量呈显著下降[17];Ge 等研究表明,T0070907 能够抑制PPARγ表达的上调和补骨脂二氢黄酮甲醚(Bavachinin)诱导的PPARγ向细胞核的迁移[18]。但本研究结果却显示DF-1 细胞内,PPARγ基因的表达量随着T0070907 浓度的增加逐渐升高,并在浓度为5 μmol/L 时极显著高于对照组(p<0.01),而同样浓度T0070907 的处理对细胞内LPL 基因的表达没有影响,表明T0070907可以特异性地促进PPARγ基因的表达。同时在蛋白水平上,本研究结果也显示T0070907 对PPARγ的过表达效果有一定的促进作用,表明T0070907 确实对PPARγ的表达有正调控作用。但T0070907 仅对DF-1细胞中的PPARγ2 的表达有效,对PPARγ1 的表达无影响,推测PPARγ的两种蛋白亚型可能存在功能上的差异。
Zaytseva 等研究表明,T0070907 虽然不改变人乳腺癌细胞内PPARγ 蛋白的表达量,但可增加PPARγ的磷酸化水平,并显著降低PPARγ与DNA 应答元件的结合活性[19]。为进一步确认T0070907 对PPARγ表达的作用,本研究利用含PPRE 反应元件的报告基因质粒来分析T0070907 对PPARγ活性的影响。PPARγ是核受体PPAR 子集的同种型,其还包括PPARα和PPARβ。PPAR 被其配体激活后,可与维甲酸X 受体(Retinoic acid X receptor,RXR)形成异二聚体,再与靶基因启动子中的一段特定的DNA 序列—PPRE 结合,实现对靶基因的转录调控作用。PPRE 是由2 个以AGGTCA 为核心的正向重复序列组成,中间大多被1 或2 个核苷酸隔开,分别与PPARs 和RXR 结合。这种PPRE 存在于参与脂质代谢和体内平衡的基因中,常被用来克隆、构建报告基因质粒,检测PPAR 的调控作用或配基筛选[20]。因此,本研究通过比较PPRE 报告基因质粒的荧光素酶活性变化,结果显示T0070907 可以显著增强PPRE 介导的荧光素酶的蛋白活性,并且T0070907不能抑制Rosi 对PPRE 介导的荧光素酶蛋白活性的激活作用,提示T0070907 能够促进PPARγ的蛋白活性,使其与靶基因的PPRE 结合,增强对靶基因的表达调控作用。
综上,本研究结果表明T0070907 对DF-1 细胞中PPARγ基因的mRNA 转录水平、PPARγ2 蛋白表达以及PPARγ的蛋白活性均具有正向调节作用。该结果为开展鸡PPARγ的功能研究提供了有益的数据,并提示,与哺乳动物不同,鸡PPARγ可能具有其独特的表达方式和调控机制。
猜你喜欢
——一道江苏高考题的奥秘解读和拓展
