一氧化氮对铝胁迫下烟草根系营养元素吸收和呼吸电子传递及内源激素含量的影响
2020-05-27刘强柳正葳刘召亮乔清华张玉
刘强 柳正葳 刘召亮 乔清华 张玉


摘要:为探明一氧化氮供体硝普钠(SNP)对铝胁迫下烟草幼苗生长和根系生理特性的促进作用,以耐铝型品种云烟100和铝敏感型品种云烟105为材料,采用水培法研究施加不同浓度SNP(0 μmol/L、50 μmol/L、100 μmol/L、200 μmol/L、400 μmol/L)对铝胁迫(200 μmol/L)下烟草幼苗生长、根系营养元素吸收、呼吸电子传递及内源激素含量的影响。结果表明,铝胁迫下烟草生长受到明显抑制,根系营养元素(钙、镁、铁、铜)含量、各呼吸途径的呼吸速率(总呼吸、细胞色素呼吸、交替呼吸)和根尖生长素、赤霉素含量均显著下降,而根尖活性氧(O2·-、H2O2)和脱落酸含量显著升高。加入适宜浓度的SNP能显著缓解铝对烟草根系和地上部生长的抑制,提高根系钙、镁、铁、铜营养元素含量和各呼吸途径的呼吸速率,促进根尖生长素、赤霉素和脱落酸合成,并降低根尖活性氧(O2·-、H2O2)含量。在试验浓度范围内以施加100 μmol/L的SNP对烟草铝毒害缓解效果最佳,而SNP为400 μmol/L时缓解效果受到抑制。综上所述,100 μmol/L的SNP可通过促进烟草根系对钙、镁、铁和铜营养元素的吸收,提高根系各呼吸途径的呼吸速率以降低根尖活性氧含量,以及维持根尖内源激素的平衡而缓解烟草铝毒害。
关键词:烟草;铝胁迫;一氧化氮;营养元素;呼吸作用;植物激素
中图分类号:Q945.78文献标识码:A文章编号:1000-4440(2020)02-0292-07
Abstract:Two tobacco cultivars of Yunyan 100 (aluminum-resistant) and Yunyan 105 (aluminum-sensitive) were selected to explore the effects of sodium nitroprusside (SNP, as the nitric oxide donor) with different concentrations (0 μmol/L, 50 μmol/L, 100 μmol/L, 200 μmol/L, 400 μmol/L) on plant growth, root nutrient elements uptake, respiratory electron transfer and endogenous hormones contents under aluminum stress (200 μmol/L) with solution culture. The results showed that aluminum stress significantly inhibited plant growth, decreased root nutrient elements (Ca, Mg, Fe, Cu) uptake, respiratory rate (total respiration, cytochrome respiration, alternative respiration), root apical auxin and gibberellin contents, but significantly increased the contents of reactive oxygen (O2·- and H2O2) and abscisic acid in root apex of tobacco seedlings. Application of SNP concentration dramatically relieved the aluminum-induced inhibition of root and aboveground growth, increased root nutrient elements (Ca, Mg, Fe, Cu) uptake, respiratory rate, the contents of auxin, gibberellin and abscisic acid in root apex, but dramatically decreased reactive oxygen (O2·- and H2O2) contents. In this experiment, SNP at 100 μmol/L had the best effects on relieving aluminum toxicity in tobacco seedlings, while the alleviating effects were suppressed when SNP concentration was 400 μmol/L. In conclusion, treatment with SNP at 100 μmol/L could alleviate aluminum toxicity in tobacco seedlings through promoting root nutrient elements (Ca, Mg, Fe, Cu) uptake, increasing respiratory rate in order to decrease reactive oxygen (O2·- and H2O2) accumulation, maintaining balance of endogenous phytohormones in root apex.
Key words:tobacco;aluminum stress;nitric oxide;nutrient elements;respiration;phytohormones
鋁是限制酸性土壤上作物生长的最主要因素之一[1]。铝毒害可破坏植物根冠细胞超微结构,影响根系发育和营养元素的吸收,导致根伸长生长的快速抑制。铝会不同程度抑制植物根系对钾、钙、镁、铁、锰、铜、锌等营养元素的吸收和运输[2],导致根系呼吸功能紊乱[3],生成O2·-和H2O2等活性氧造成植物氧化伤害[4]。此外,铝对植物体内激素代谢平衡也会产生显著影响。铝胁迫下黑麦和小麦根系脱落酸含量显著升高,赤霉素含量显著下降,生长素含量则有升有降[5],然而目前仍不清楚铝诱导的激素含量变化是否只是铝毒害的次生效应。
一氧化氮(NO)作为重要的生物活性分子广泛参与植物各种生理活动以及植物对生物和非生物胁迫的应答过程[6]。近年来,NO被认为是介导植物耐铝响应的重要信号分子之一,已成为研究热点。外源NO可通过提高植物抗氧化胁迫能力[7]、促进根系有机酸分泌[8]、影响细胞壁组分和性质[9]、诱导抗氧化酶和细胞壁合成相关蛋白质基因表达[10]等途径增强植物耐铝性。但也有报道表明外源NO加剧了铝诱导的饭豆根伸长抑制[11]。这种差异可能与NO处理浓度、处理时间及植物对铝敏感性不同等因素有关[12]。
烟草是普遍栽培的经济作物,也是遗传背景清晰的模式植物。中国烤烟主产区在南方酸性土壤上有大面积分布,铝毒害问题较为严重,常造成作物大幅减产[13]。前期研究发现,外源NO可通过调控烟草叶片叶绿体和线粒体内能量代谢以降低细胞内活性氧生成而增强烟草耐铝性[14-15],但关于NO对铝胁迫下烟草根系营养元素吸收、呼吸电子传递和内源激素含量的影响,以及与植物耐铝性的关系尚不清楚。本研究以2个耐铝性差异明显的烟草品种为试验材料,分析外施不同浓度NO供体硝普钠(SNP)对铝胁迫下烟草生长、根系营养元素吸收、呼吸作用、活性氧和内源激素含量变化的调控作用,旨在为外源NO施用提高植物耐铝性提供一定的科学依据和实践应用基础。
1材料与方法
1.1材料与试剂
供试烟草(Nicotiana tabacum L.)品种为前期筛选得到的耐铝型云烟100和铝敏感型云烟105。铝和NO分别以AlCl3·6H2O和硝普钠([Na2Fe(CN)5]·NO)形态提供,分析纯。
1.2试验方法
烟草种子经1%NaClO溶液消毒30 min,用去离子水3次清洗后浸种24 h。之后将烟草种子均匀播种于育苗基质(草炭∶珍珠岩=3∶1,体积比)中进行培养,期间用1/5 Hoagland(pH 4.5)营养液浇灌。待幼苗长至3~4片真叶时,选取长势一致的幼苗移栽到1 L含1/5 Hoagland(pH 4.5)营养液的盆钵,每盆移栽8棵幼苗。预培养7 d后,采用完全随机设计共设置6个处理,每个处理3次重复,试验设计如下:①对照CK(AlCl3 0 μmol/L+SNP 0 μmol/L);②Al(AlCl3 200 μmol/L+SNP 0 μmol/L);③Al+50(AlCl3 200 μmol/L+SNP 50 μmol/L);④Al+100(AlCl3 200 μmol/L+SNP 100 μmol/L);⑤Al+200(AlCl3 200 μmol/L+SNP 200 μmol/L);⑥Al+400(AlCl3 200 μmol/L+SNP 400 μmol/L)。以上处理液均含1/5 Hoagland营养液,AlCl3和SNP均为直接加入营养液中,最终将各处理液均调节pH为4.5。处理液每3 d更换1次,每天通气2 h。处理15 d后测定植株根系和地上部生物量,同时取整株根系测定钾、钙、镁、铁、锰、铜、锌等元素含量、总呼吸、细胞色素呼吸和交替呼吸速率,取主根根尖0~1 cm测定O2·-和H2O2含量以及生长素、赤霉素和脱落酸含量。
1.3测定方法
元素含量测定:根系样品用HNO3-HClO4进行湿式消解,获得的待测液采用8000-DV型电感耦合等离子发射光谱仪(美国PE公司产品)测定钾、钙、镁、铁、锰、铜、锌等7种营养元素含量。根系总呼吸、细胞色素呼吸和交替呼吸速率根据Hu等[16]的方法采用Clark型氧电极(英国Hansatech公司产品)测定。根尖O2·-和H2O2含量测定采用Zhou等[17]的方法。根尖内源生长素、赤霉素和脱落酸含量的提取采用He等[5]的方法,测定采用酶联免疫法(ELISA)。
1.4数据处理
采用Kyplot软件制图,采用SPSS 19.0软件对数据进行最小显著差数法统计分析。
2结果与分析
2.1外源NO對铝胁迫下烟草根系和地上部生物量的影响
由图1可知,铝胁迫下,云烟100和云烟105根系生物量与对照相比分别下降30.4%和48.4%,地上部生物量分别下降23.4%和35.7%,说明云烟100耐铝性强于云烟105,且根系受铝毒害更为明显。施加SNP后,烟草根系和地上部生物量随SNP浓度增加呈先升高后下降的趋势。当SNP浓度为100 μmol/L时,云烟100和云烟105植株生物量升幅最大,根系生物量较铝胁迫处理分别升高24.5%和38.0%,地上部生物量分别升高18.5%和30.5%。但400 μmol/L SNP则显著加剧了铝诱导的云烟105生长抑制而对云烟100影响不显著(图1)。
2.2外源NO对铝胁迫下烟草根系营养元素吸收的影响
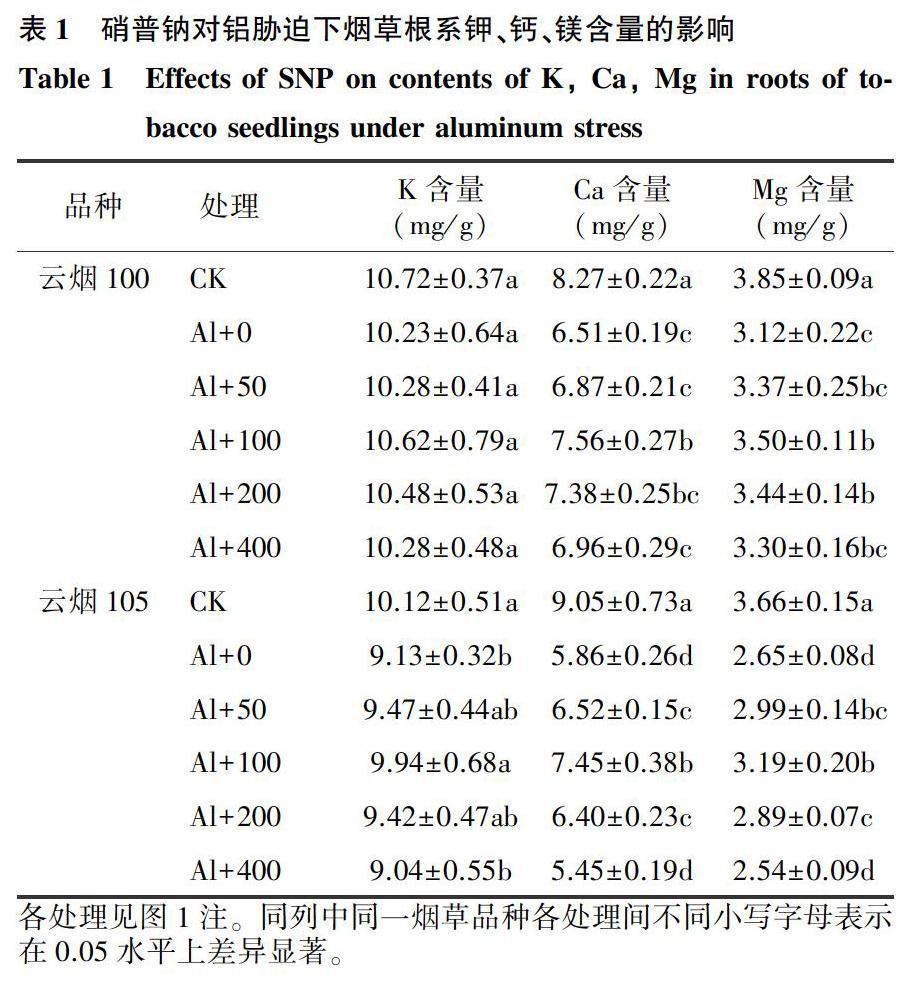
2.2.1外源NO对铝胁迫下烟草根系钾、钙、镁大量营养元素吸收的影响由表1可知,铝胁迫导致云烟100和云烟105根系钾含量分别下降4.6%和9.8%,钙含量分别下降21.3%和35.2%,镁含量分别下降19.0%和27.6%,说明铝对烟草根系钙、镁吸收的影响大于钾。外施SNP后,烟草根系对钾、钙、镁营养元素的吸收随NO浓度增加呈先升高后下降的趋势,并均在SNP 100 μmol/L时达到最大值。与Al处理组相比,Al+100处理云烟100和云烟105根系钾含量分别升高3.8%和8.9%,钙含量分别升高16.1%和27.1%,镁含量分别升高12.2%和20.4%(表1),表明NO可显著促进烟草根系对钙、镁元素的吸收。
2.2.2外源NO对铝胁迫下烟草根系铁、锰、铜、锌微量营养元素吸收的影响由表2可知,与对照相比,铝胁迫导致云烟100和云烟105根系铁含量分各处理见图1注。同列中同一烟草品种各处理间不同小写字母表示在0.05水平上差异显著。
别下降23.6%和38.2%,铜含量分别下降13.7%和26.9%,而云烟100根系锰和锌含量无显著变化,云烟105则显著下降。外施SNP后,烟草根系对铁、锰、铜、锌微量营养元素的吸收随SNP浓度增加呈先升高后下降的趋势。其中云烟100根系铁、铜含量在SNP 100 μmol/L时达到最大值,分别比铝处理组显著升高16.7%和9.2%,而锰、锌含量变化不显著;对于云烟105,在SNP 100 μmol/L时根系铁、锰、铜和锌含量最大,分别比铝处理组显著升高25.2%、8.9%、19.2%和9.4%;但当SNP为400 μmol/L时,云烟105根系铁含量比铝处理组显著下降10.1%(表2),表明高浓度SNP进一步抑制了铝诱导的敏感品种根系对铁的吸收。
2.3外源NO对铝胁迫下烟草根系呼吸电子传递的影响
2.3.1外源NO对铝胁迫下烟草根系呼吸速率的影响由表3可知,铝处理显著降低了烟草各呼吸途径的呼吸速率,云烟100和云烟105根系总呼吸速率比对照分别显著下降20.5%和31.6%,细胞色素呼吸速率分别下降26.7%和41.2%,交替呼吸速率分别下降8.1%和15.7%。SNP的施加提高了烟草根系各呼吸途径的呼吸速率,且随SNP浓度增加,2种烟草根系各呼吸途径的呼吸速率呈先升高后下降的趋势。其中云烟100在SNP浓度为200 μmol/L时总呼吸和细胞色素呼吸速率最大,较铝处理组分别显著升高13.4%和15.7%,而云烟105则在SNP浓度为100 μmol/L时总呼吸和细胞色素呼吸速率最大,较铝处理组分别显著升高17.6%和20.8%。同时云烟100和云烟105交替呼吸速率均在SNP浓度为100 μmol/L时达到最大值,分别较铝处理组显著升高7.53%和13.2%。当SNP浓度为400 μmol/L时,云烟105根系各呼吸途径速率较铝处理显著下降,而云烟100无显著差异(表3),表明高浓度SNP会进一步抑制铝敏感品种云烟105根系呼吸作用。
2.3.2外源NO对铝胁迫下烟草根尖活性氧含量的影响由图2可知,铝处理显著提高了烟草根尖活性氧含量,与对照相比,云烟100和云煙105根尖O2·-产生速率分别升高51.2%和89.3%,H2O2含量分别升高39.6%和65.2%。外源加入SNP后,烟草根尖活性氧含量随SNP浓度增加呈先下降后升高的趋势,并均在SNP浓度为100 μmol/L时达到最低值。此时,云烟100和云烟105 根尖O2·-产生速率较铝处理组分别下降14.3%和29.5%,H2O2含量分别下降16.9%和36.5%。而当SNP浓度为400 μmol/L时,云烟105根尖O2·-产生速率和H2O2含量显著高于铝处理组,但云烟100与铝处理组差异不显著(图2),表明高浓度SNP导致了铝诱导的铝敏感品种云烟105根尖氧化胁迫作用加剧。
2.4外源NO对铝胁迫下烟草根尖内源激素含量的影响
由表4可知,铝胁迫下,烟草根尖生长素和赤霉素含量显著下降,而脱落酸含量显著升高。云烟100和云烟105根尖生长素含量分别比对照下降17.6%和23.1%,赤霉素含量分别比对照下降25.1%和32.9%,而脱落酸含量分别比对照升高76.4%和65.2%。外源加入SNP后,烟草根尖生长素和赤霉素含量均随SNP浓度增加呈先升高后下降的趋势,云烟100和云烟105均在SNP浓度为100 μmol/L时达到最大值,生长素含量分别比铝处理组分别显著升高19.8%和26.6%,赤霉素含量分别比铝处理组显著升高50.4%和62.6%。云烟100和云烟105根尖脱落酸含量均随SNP浓度增加而升高,在SNP浓度为400 μmol/L时达到最大值,分别比铝处理组显著升高80.5%和90.4%(表4)。
3讨论
根尖是铝毒害的最初作用部位,其毒害症状主要表现为根伸长抑制,水分和养分的吸收能力减弱,从而导致植株生物量下降[18-19]。本研究中铝胁迫显著抑制了烟草幼苗生长,而外施50~200 μmol/L SNP均可有效缓解烟草铝毒害,其中以100 μmol/L SNP缓解效果最好,表明适宜浓度的NO可有效缓解植物铝毒害,这与在大豆[20]、小麦[21]等研究中取得的结果相一致。但当 SNP浓度升至400 μmol/L时,NO的缓解作用显著减弱甚至对铝敏感品种云烟105产生更严重的毒害作用。这可能与NO是一种重要的活性氮形态,其过量积累会导致硝化胁迫从而产生新的氧化伤害有关[22]。
铝毒害的一个重要机制是抑制根系对养分的吸收,最终导致植物体内营养元素的亏缺或失衡[23]。钙可维持细胞膜结构稳定性,镁是叶绿素组分元素,铁、锰、铜等微量营养元素则作为叶绿体和线粒体电子传递体的组分在光合和呼吸代谢中起着重要作用[24]。本研究中,铝处理对烟草根系钙、镁、铁和铜元素的吸收抑制显著大于钾、锰和锌等元素。在荞麦[2]、茶树[25]等研究中也发现铝显著抑制了根系对营养元素的吸收。其机制可能与铝阻塞了根原生质膜钙离子通道[26],将镁从根质外体结合位点置换[27]及降低根系呼吸作用[3]有关。外加100 μmol/L SNP则显著提高了2个烟草品种铝胁迫下根系钙、镁、铁和铜营养元素含量,表明适宜浓度的NO可在一定程度上通过增强根系对营养元素的吸收而有效缓解铝诱导的膜结构破坏和光合呼吸代谢生理紊乱等不利影响。而加入400 μmol/L SNP后,这种缓解效应显著减弱甚至对云烟105根系铁吸收产生新的抑制,可见高浓度NO会干扰烟草根系对微量元素的吸收。分析其原因可能与本研究中高浓度NO造成铝诱导的根系氧化伤害加剧和呼吸作用显著下降从而导致养分吸收受阻有关,具体原因有待进一步分析。
植物线粒体电子传递至O2主要由主路细胞色素途径和支路交替途径构成。逆境胁迫常造成细胞色素途径受抑,导致电子传递受阻致使部分电子可直接漏出对O2进行单电子还原形成O2·-[28]。本研究中发现,铝胁迫下烟草根系呼吸速率尤其是细胞色素途径呼吸显著下降,影响到线粒体呼吸链上电子的有效传递,从而导致O2·-产生速率和H2O2含量显著升高。在铝诱导的水稻根系[29]和烟草叶片[15]研究中也发现类似现象。外施SNP后,烟草根系各呼吸途径的呼吸速率显著升高,O2·-和H2O2含量显著下降,其中以100 μmol/L SNP效果最好,400 μmol/L SNP反而较差。这些结果说明,适宜浓度的NO能有效降低活性氧的生成,分析其主要机制:一方面可通过提高烟草根系呼吸作用使电子在呼吸链上顺利传递给O2而减少电子泄漏,另一方面可能与NO作为信号分子可直接猝灭活性氧或激活植物体内抗氧化系统有关[7]。
植物內源激素及其相互作用被普遍认为可调节各种环境条件下根系的生长和发育[30-31]。有研究者报道铝抑制紫花苜蓿生长与根尖内源生长素积累降低有关[32],低浓度外源脱落酸可有效缓解栝楼铝毒害[33],可见植物体内激素含量的变化与铝毒害/耐铝性密切相关。同时He等[5]也指出植物铝毒害/耐铝性不仅取决于内源激素含量,还与各激素之间比例的协调性有关。本研究中,铝处理抑制了烟草根尖生长素和赤霉素合成,而促进了脱落酸含量增加,这与铝胁迫下小麦[5]根尖生长素和赤霉素含量下降,大豆[34]、大麦[35]脱落酸含量升高等研究结果相一致。外施100 μmol/L SNP后,烟草根尖生长素和赤霉素含量升幅最大,脱落酸含量显著升高,此时烟草生长促进效应最明显,表明该浓度的NO可有效调节烟草根尖内源激素含量而有效缓解铝对植物的毒害。但当SNP浓度为400 μmol/L时,烟草根尖脱落酸含量进一步升高,其升幅远大于生长素和赤霉素,在一定程度上造成烟草根尖内源激素代谢平衡失调,尤以云烟105变化更明显,此时云烟100根系生长与铝处理组相比无显著变化,而云烟105则显著下降,这可能与2个烟草品种对铝和NO双重胁迫的不同反应有关。
综上所述,铝胁迫显著降低了烟草生物量。适宜浓度外源SNP可通过促进烟草根系对钙、镁、铁和铜营养元素的吸收,提高根系各呼吸途径的呼吸速率以降低根尖活性氧含量,以及维持根尖内源激素的平衡而缓解烟草铝毒害。SNP缓解效果因浓度不同而存在明显差异,其中以100 μmol/L SNP减轻铝毒害效果最佳。
参考文献:
[1]刘强,郑绍建,林咸永.植物适应铝毒胁迫的生理及分子生物学机理 [J].应用生态学报,2004,15(9):1641-1649.
[2]陈文荣,刘鹏,黄朝表,等.铝对荞麦铝和其它营养元素运输的影响 [J].水土保持学报,2006,20(3):173-176.
[3]YAMAMOTO Y,KOBAYASHI Y,DEVI S R,et al.Aluminum toxicity is associated with mitochondrial dysfunction and the production of reactive oxygen species in plant cells [J].Plant Physiology,2002,128(1):63-72.
[4]PEREIRA L B,MAZZANTI C M,CARGNELUTTI D,et al. Differential responses of oat genotypes: oxidative stress provoked by aluminum [J].Biometals,2011,24(1):73-83.
[5]HE H Y,HE L F,GU M H,et al. Nitric oxide improves aluminum tolerance by regulating hormonal equilibrium in the root apices of rye and wheat [J].Plant Science,2012,183(2):123-130.
[6]YU M,LAMATTINA L,SPOEL S H,et al. Nitric oxide function in plant biology: a redox cue in deconvolution [J].New Phytologist,2014,202(4):1142-1156.
[7]SUN C L,LIU L J,LU L L,et al. Nitric oxide acts downstream of hydrogen peroxide in regulating aluminum-induced antioxidant defense that enhances aluminum resistance in wheat seedlings [J]. Environmental and Experimental Botany,2018,145(1):95-103.
[8]WANG H H,HUANG J J,LIANG W H,et al. Involvement of putrescine and nitric oxide in aluminum tolerance by modulating citrate secretion from roots of red kidney bean [J].Plant and Soil,2013,366(2):479-490.
[9]ZHANG Z Y,WANG H H,WANG X M,et al. Nitric oxide enhances aluminum tolerance by affecting cell wall polysaccharides in rice roots [J].Plant Cell Reports,2011,30(9):1701-1711.
[10]YANG L M,TIAN D G,TODD C D,et al. Comparative proteome analyses reveal that nitric oxide is an important signal molecule in the response of rice to aluminum toxicity [J].Journal of Proteome Research,2013,12(3):1316-1330.
[11]ZHOU Y,XU Y Y,CHEN L Q,et al. Nitric oxide exacerbates Al-induced inhibition of root elongation in rice bean by affecting cell wall and plasma membrane properties [J].Phytochemistry,2012,76(1):46-51.
[12]WANG H H,HUANG J J,BI Y R. Nitrate reductase-dependent nitric oxide production is involved in aluminum tolerance in red kidney bean roots [J]. Plant Science,2010,179(3):281-288.
[13]尹永强,何明雄,邓明军.土壤酸化对土壤养分及烟叶品质的影响及改良措施 [J]. 中国烟草科学,2008,29(1):51-54.
[14]刘强,贺根和,柳正葳,等.外源一氧化氮对铝胁迫下烟草叶片光能利用和光保护系统及活性氧代谢的影响[J]. 湖南农业大学学报,2016,42(6):615-621.
[15]刘强,柳正葳,胡萃,等.外源一氧化氮对铝胁迫下烟草叶片呼吸作用和活性氧代谢的影响[J]. 河南农业科学,2017,46(1):42-47.
[16]HU W H,XIAO Y A,ZENG J J,et al. Photosynthesis, respiration and antioxidant enzymes in pepper leaves under drought and heat stresses [J]. Biologia Plantarum,2010,54(4):761-765.
[17]ZHOU Y H,YU J Q,HUANG L F,et al. The relationship between CO2 assimilation, photosynthetic electron transport and water-water cycle in chill-exposed cucumber leaves under low light and subsequent recovery [J]. Plant Cell and Environment,2004,27(12):1503-1514.
[18]王志颖,刘鹏,李锦山,等.铝胁迫下外源有机酸对油菜根系形态及叶绿素荧光特性的影响[J]. 江苏农业学报,2011,27(4):756-762.
[19]于力,孙锦,郭世荣,等.铝胁迫对豇豆幼苗根系生理指标的影响[J]. 江苏农业学报,2013,29(2):319-323.
[20]CAI M Z,ZHANG S N,WANG F M,et al. Protective effect of exogenously applied nitric oxide on aluminum induced oxidative stress in soybean plants [J]. Russian Journal of Plant Physiology,2011,58(5):791-798.
[21]SUN C L,LIU L J,YU Y,et al. Nitric oxide alleviates aluminum-induced oxidative damage through regulating the ascorbate-glutathione cycle in roots of wheat [J]. Journal of Integrative Plant Biology,2015,57(6):550-561.
[22]CORPAS F J,LETERRIER M,VALDERRAMA R,et al. Nitric oxide imbalance provokes a nitrosative response in plants under abiotic stress [J]. Plant Science,2011,181(5):604-611.
[23]RENGEL Z,ROBINSON D L. Aluminum and plant age effects on absorption of cations in the Donnan free space of rye grass roots [J]. Plant Soil,1989,116(2):223-227.
[24]潘瑞熾.植物生理学 [M]. 北京:高等教育出版社,2012.
[25]段小华,邓泽元,胡小飞,等.模拟酸雨和外源铝对茶树铝及一些营养元素吸收积累的影响[J]. 农业环境科学学报,2010,29(10):1936-1942.
[26]HUANG J W,PELLET D M,PAPERNIK L A,et al. Aluminum interactions with woltage dependant calcium transport in plasma membrane vesicle isolated from roots of aluminum-sensitive and resistant wheat cultivars [J]. Plant Physiology,1996,110(2):561-569.
[27]袁世力,刘星辰,樊娜娜,等.镁对铝胁迫紫花苜蓿幼苗生长和光合系统的影响[J]. 中国草地学报,2018,40(5):29-35.
[28]DAVIDSON J F,SCHIESTL R H. Mitochondrial respiratory electron carriers are involved in oxidative stress during heat stress in Saccharomyces cerevisiae [J]. Molecular and Cellular Biology,2001,21(24):8483-8489.
[29]谢国生,师瑞红,庞贞武,等.铝胁迫下水稻幼苗根系的生理特性[J]. 应用生态学报,2009,20(7):1698-1704.
[30]KOPITTKE P M. Role of phytohormones in aluminium rhizotoxicity [J]. Plant Cell and Environment,2016,39(10):2319-2328.
[31]赵爽,郑刚,季梦成,等. 山木通扦插生根过程中内源激素的动态变化[J].江苏农业科学,2018,46(3):148-150.
[32]任晓燕,周鹏,安渊.喷施IAA对紫花苜蓿幼苗铝毒害的缓解作用[J]. 草业科学,2014,31(7):1323-1329.
[33]李婧,刘星星,吴玉环,等.外源脱落酸对铝胁迫下栝楼叶绿素荧光及生理活性的影响[J]. 水土保持学报,2017,31(2):293-300.
[34]SHEN H,LIGABA A,YAMAGUCHI M,et al. Effect of K-252a and abscisic acid on the efflux of citrate from soybean roots [J]. Journal of Experimental Botany,2004,55(397):663-671.
[35]KASAI M,SASAKI M,TANAKAMARU S,et al. Possible involvement of abscisic acid in increases in activities of two vacuolar H+-pumps in barley roots under aluminum stress[J]. Plant Cell Physiology,1993,34(8):1335-1338.
(责任编辑:张震林)
