酿酒酵母中基于CRISPR/dCas9 的基因转录调控工具的开发与应用
2020-05-13陈永灿张建志司同
陈永灿 张建志 司同
(中国科学院深圳先进技术研究院,深圳合成生物学创新研究院,深圳 518055)
酿酒酵母被用于制作面包和酿酒已有数千年的历史,是与人类关系最密切的微生物之一。由于其具有遗传背景清晰、生长迅速、抗逆性高、无噬菌体污染等优势,在生物制药、食品、化工和能源等领域均有广泛应用[1-2]。同时,酿酒酵母还是现代分子和细胞生物学研究的真核模式生物。
基因编辑和转录调控工具在酿酒酵母的基础研究和工业应用中起着十分重要的作用。细菌和古细菌中存在CRISPR/Cas9(Clustered regularly interspaced short palindromic repeats/CRISPR-associated protein 9)系统,可以特异性地降解外源遗传物质,进行适应性免疫防御。近年来,该系统被广泛开发应用于包括酿酒酵母在内不同物种的基因编辑[3-6]。CRISPR/Cas9 系统由核酸内切酶Cas9(CRISPRassociated protein 9)、CRISPR RNA(crRNA)和反式激活RNA(trans-activating crRNA,tracrRNA)组成。其中,crRNA 与tracrRNA 形成二级结构并招募Cas9蛋白;crRNA 和tracrRNA 也可以融合为向导RNA(guide RNA,gRNA)以简化操作。Cas9 蛋白与核酸结合的复合体可以特异性结合到PAM(Photospacer adjacent motif,NGG)位点相邻并与crRNA 互补的DNA 序列。接着,Cas9 内切酶的HNH 结构域切割与crRNA 互补的DNA 链,RuvC 结构域切割另一条DNA 链,形成DNA 双链断裂(double-strand break,DSB),断裂位点位于PAM 位点上游三个核苷酸的位置[7]。同源重组(Homologous recombination,HR)是酿酒酵母中主要的DNA 修复方式。同源片段存在时,通过CRISPR/Cas9 特异性引入DNA 双链断裂,可对酿酒酵母基因组进行精准的敲除、敲入和突变[8-9]。
除基因编辑外,CRISPR 系统在转录调控、表观遗传编辑、碱基编辑和细胞成像等方面也有广泛应用[10-12]。这些应用主要基于核酸内切酶活性缺陷的dCas9(Nuclease-deficient Cas9)蛋白,而这一突变体通过在Cas9 的RuvC 和HNH 结构域分别引入D10A 和H840A 突变获得[13]。酿酒酵母中,将dCas9 靶向基因的启动子区域,即有一定的转录抑制作用;但通过融合或募集不同类型的转录调控结构域,如转录抑制因子、激活因子或表观遗传修饰酶等[13-17],并结合精巧的gRNA 设计[18-19],可以进一步实现多模式(Multi-mode)、多位点(Multiplex)、正交性(Orthogonal)的基因转录调控。
本文针对酿酒酵母中基于CRISPR/dCas9 的基因转录调控工具的开发和应用进行综述。介绍了通过与不同转录调节结构域的结合来调节dCas9 或gRNA的活性,概括了dCas9 和gRNA 表达的设计与优化方法;讨论了影响CRISPR/dCas9 体系转录调控效率、特异性和通量的靶向性因素;总结了该工具在酿酒酵母代谢工程中的应用;最后,对该工具的未来研究方向提出了展望。
1 酿酒酵母中CRISPR/dCas9 工具的开发
1.1 针对基因转录调控模式的设计与优化
转录调控因子通常由DNA 结合结构域和转录调控结构域组成。研究人员将dCas9 或gRNA 进行改造,使CRISPR/dCas9 系统与转录抑制因子、激活因子或表观遗传修饰酶等相结合,从而将这些转录调控结构域靶向目的基因的特定位置,实现不同模式的基因调控[20](图1)。
1.1.1 dCas9 融合转录调控结构域 由于空间位阻效应,当gRNA 引导dCas9 靶向目的基因的特定位置时,能起到转录抑制作用,被称为CRISPR 抑制(CRISPR interference,CRISPRi)[13,21]。在细菌中单纯使用dCas9 进行基因表达抑制的效率可高达99.9%[13],而在酿酒酵母中,单纯利用dCas9 通常仅能起到50%-60%的抑制效果[18,21-22],这样的抑制水平与其它真核生物中的报道一致[23-24]。这些实验结果表明,单独使用dCas9 无法在真核细胞中高效抑制转录起始或阻挡RNA 聚合酶的前进。另一方面,当dCas9 与转录抑制结构域融合时,可显著提高其在真核体系中的转录抑制能力[25](图1-A)。Gilbert 等[23]将dCas9 与哺乳动物来源的Mxi1 蛋白的转录抑制结构域[26]相融合,发现对报告基因的表达抑制效果从单纯使用dCas9 时的18 倍提高至53 倍。在解脂耶氏酵母中,dCas9-Mxi1 相比dCas9 把Ku80基因的转录抑制效率从38%提高至87%[27]。为了进一步优化CRISPRi 在酿酒酵母中的效率,Lian 等筛选了多个内源转录抑制因子[28-32],发现在与dCas9 结合时,酿酒酵母TUP1、MIG1、XTC1、UME6 等蛋白的结构域的转录抑制水平高于哺乳细胞Mxi1。他们同时发现将多个内源抑制结构域同时与dCas9 融合时,可以进一步提高转录抑制效率。例如,当靶向酿酒酵母的组成型PTEF1启动子时,dCas9-Mix1、dCas9-TUP1 和dCas9-TUP1-MIG1-UME6对于 荧光报告基因表达的抑制效率分别为65%、74%和82%[19]。综合以上结果表明,与转录抑制结构域的融合能够显著提升dCas9 在酿酒酵母中的转录抑制效率,并且可以利用内源转录因子、融合多重结构域等方式进一步提升抑制效率。
dCas9 与转录激活结构域融合可实现对靶基因表达的上调,被称为CRISPR 激活(CRISPR activation,CRISPRa)[33]。常用的转录激活结构域有VP16、VP64、p65AD 和Rta 等[34-36]( 图1-A)。Farzadfard 等[21]在酿酒酵母中将dCas9 与VP64 融合并靶向最小化PCYC1启动子,使报告基因的荧光强度提高了3 倍左右。在dCas9-VP64 的基础上融合多个转录激活结构域,可以进一步增强基因上调效率[36-38]。如Chavez 等利用融合了3 个转录激活结构域的dCas9-VPR(VP64-p65AD-Rta),使HED1和GAL7的表达分别提高了38 倍和78 倍,而dCas9-VP64仅使二者分别提高 了9倍和14 倍[36]。此外,对于不同的转录激活结构域,融合不同来源的CRISPR 蛋白也会导致激活效率的差异。Lian 等测试了4 种不同来源的内切酶活性缺失Cas 蛋白(dSpCas9,dSaCas9,dSt1Cas9,dLbCpf1) 与3 种转录激活结构域(VP64(V),VP64-p65AD(VP),VP64-p65AD-Rta(VPR))的所有组合,发现不同的dCas 蛋白最优的激活结构域组合是不同的:对于dSpCas9,激活效率最好的是VPR;对于dSt1Cas9,激活效率最好的是V;对于dLbCpf1,激活效率最好的是VP;而对于dSaCas9,不同组合激活效果均比较微弱[19]。以上结果表明,dCas9 与转录激活结构域的融合蛋白可有效上调靶基因转录水平,但不同dCas 蛋白和激活结构域的组合需要分别进行优化。
1.1.2 gRNA 融合适配子 除了将转录调控结构域与dCas9 蛋白融合外,还可以改造gRNA 结构使其同时具有目的基因靶向序列和效应蛋白招募序列,从而利用RNA-蛋白质相互作用招募转录调控结构域[25]。Zalatan 等和Kiani 等将RNA 适配子(RNA aptamer)与gRNA 的3'末端的tracrRNA 融合,另外将转录抑制或激活结构域与RNA 适配子结合蛋白融合,通过RNA 适配子与其结合蛋白的相互作用招募转录调控结构域,实现了靶基因的特异性转录调控(图1-B)[39-40]。利用Scaffold RNA(scRNA)策略与融合VP64 的RNA 结合蛋白,Zalatan 等在酿酒酵母中将一个合成型报告启动子激活了50 倍,而dCas9-VP64 的策略仅实现了2-3 倍的激活[39]。与dCas9 融合多个转录抑制或激活结构域策略类似,作者将多个适配子融合在同一个scRNA 中,从而可招募多个RNA 结合蛋白以增强其转录调控的能力[39]。另外,通过系统性测试,Zalatan 等[39]发现了特异性的3 个RNA 适配子—结合蛋白的配对(MS2:MCP、PP7:PCP 和com:COM,图1-B),为多个结合蛋白与不同调控结构域融合构建正交性的转录调控因子奠定基础。综上,scRNA 技术提供了一种与dCas9 融合蛋白技术相互补的策略,可以对多基因靶标进行多模式调控,在基因互作研究、代谢通路组合式优化等方面有重要的应用价值。
1.2 针对dCas9或gRNA表达的设计与优化
优化dCas9 蛋白的定位、表达水平和诱导表达等性质,可以提高CRISPR/dCas9 系统的转录调控效率。dCas9 需要定位到细胞核中以发挥转录调控的功能,通常情况下,会在dCas9 的氨基端或羧基端融合一段SV40 核定位序列(Nuclear localization sequence,NLS)[21,41-42],也可在两端同时融合一段NLS 序列[43]或在羧基端融合两段NLS 序列[23]。优化dCas9 蛋白表达量的手段包括调节密码子偏好、启动子强度和表达载体拷贝数等。由于不同物种同义密码子偏好性不同,密码子优化是提高重组蛋白异源表达效率的可能途径[44];酿酒酵母中,基于化脓性链球菌野生型、人或酿酒酵母密码子偏好性优化的dCas9 序列均有所应用[21,23,38,42,45]。通常情况下,转录调控因子的表达量越高,对靶基因的抑制或激活的效率也越高[46]。因此,大部分研究使用了比较强的组成型启动子或糖酵解途径启动子来驱动dCas9 的表达。例如,PTEF1、PTDH3、PGPD1和PPDC1等[23,36,42](表1),但是也有研究表明强启动子驱动的dCas9 的表达会降低酿酒酵母的适应度和生长速度[47]。对于dCas9 基因的表达载体,既可将dCas9 表达框整合到酿酒酵母的基因组中[21],也可将其克隆到自主复制质粒上[23,38,42,45](表1)。除了dCas9 的组成型表达外,还可以使用诱导型启动子使其在化学小分子或光诱导条件下表达。例如,通过利用半乳糖和无水四环素诱导合成型启动子驱动dCas9-VP64 的基因转录,Farzadfard 等在酿酒酵母中将最小化PCYC1启动子驱动的荧光报告基因的转录水平提高了70 倍[21,48]。在蛋白稳定性方面,Khakhar 等[45]通过融合降解子序列,实现了植物生长素诱导的SCF(Skp1-Cullin-F-box)泛素连接酶复合物依赖的dCas9 蛋白降解。这些条件性表达或稳定性调控的策略进一步提高了CRISPR/dCas9 系统在转录调控中应用的可控性[49-51]。
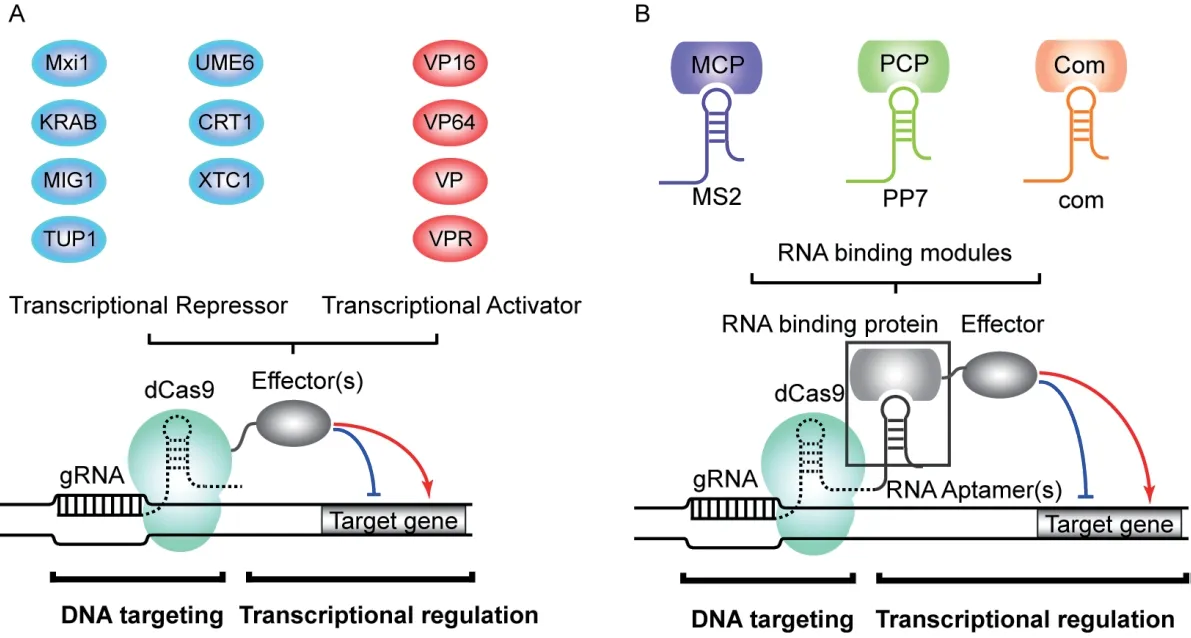
图1 通过融合或招募转录调节结构域调整dCas9 或gRNA 的活性
调节gRNA 的转录水平也是优化CRISPR/dCas9转录调控性能的重要方法。首先,利用不同类型的天然启动子可以用来调节gRNA 的转录水平。与RNA 聚合酶相对应,真核生物启动子可分为I 型(rRNA)、II 型(mRNA) 和III 型(snoRNA、tRNA等)。由于II 型启动子会在gRNA 的5'和3'端添加额外的核苷酸从而干扰其正常功能,因此一般利用III 型启动子驱动gRNA 的表达,如PSNR52和PRPR1等[6,21,23](表1)。然而,由于III 型启动子的转录本长度较短,为了提高gRNA 设计和表达的灵活性,Gao 等[52]通过在gRNA 两端分别融合锤头型核酶(Hammerhead-type ribozymes)和HDV 核酶(Hepatitis delta virus)在酿酒酵母细胞中实现了由II 型PADH1启动子驱动的gRNA 的表达(表1)。这种含有核酸酶的gRNA 称为RGR(Ribozyme-flanked gRNA)。基于该设计,Deaner 等[18]利用II 型的PTEF1启动子将gRNA 的转录水平比III 型的PSNR52启动子提高了3.88 倍,显著提高了dCas9-VPR 介导的转录抑制效率(gRNA 靶点的位置效应,见下文)。而Gander等[53]利用最小化PCYC1启动子驱动RGR 的转录,基于dCas9-Mxi1 建立的非逻辑门显示了最小的泄漏并具有数字响应,为更加复杂的遗传电路研究奠定了基础。除组成型启动子外,还可以改造天然的III型启动子使其具有对环境因素变化响应的能力,从而增强dCas9 调控工具的可控性。例如,研究人员通过在III型PRPR1启动子中整合Tet操纵子结合位点,实现无水四环素对gRNA 的诱导转录,以及2-20 倍的转录调控动态范围[21,38,42,54]。这些不同启动子驱动的多种设计方案为gRNA 的表达量、通量和对gRNA 表达的控制提供了有力的工具和多样的选择。
1.3 针对靶向性能的设计与优化
除了dCas9 和gRNA 元件的表达外,靶向性能也是决定CRISPR/dCas9 基因调控能力的重要因素。具体而言,dCas9/gRNA 复合体能否容易且准确地接近单个或多个目的区域的特定位点,决定了转录调控的有效性、准确性和多重性。以下将从gRNA 的序列特征、靶点位置与可接近性、靶基因数量等3个方面总结了影响CRISPR/dCas9 工具靶向性能的关键因素。
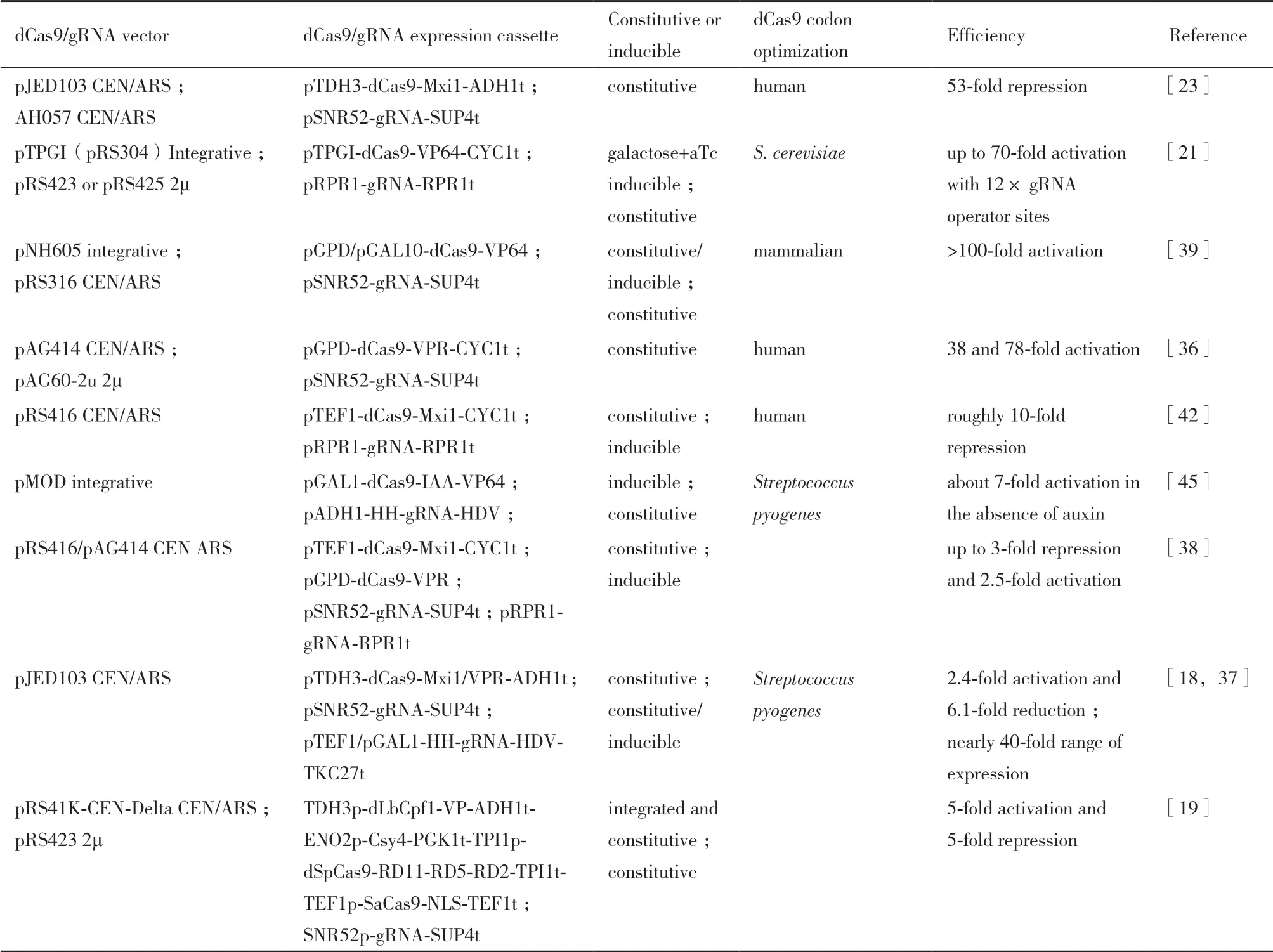
表1 酿酒酵母中dCas9 和gRNA 的表达
1.3.1 gRNA 的长度、 序列特异性和二级结构 gRNA中的向导序列(Guide sequence) 与DNA 靶序列互补配对,其长度和序列特异性对于CRISPR/dCas9 系统的靶向特异性有重要影响(图2-A)。在大肠杆菌的CRISPRi 研究中,Qi 等[13]对向导序列的长度进行了截短或延长,发现20 nt 的原始长度是转录抑制效率最高的,并且随着该区域长度的截短导致转录抑制效率快速下降,最短的有效长度是12 nt;在哺乳动物细胞中,Kiani 等[40]发现使用10-20 nt 的向导序列均能使dCas9-VPR 有效地发挥转录激活的功能。但是,在向导序列长度小于18 nt 时Cas9 介导的基因编辑功能基本缺失,表明转录调控相比基因编辑对向导序列的长度变化有更大的耐受性。对于向导序列的特异性,Qi 等[13]对向导序列单点突变的筛选结果表明,PAM 及距其最近的7 个碱基对转录抑制至关重要。Smith 等[42]在酿酒酵母中的研究结果与此一致,无论是全长还是截短型向导序列,距PAM 最近的10 nt 对dCas9-Mxi1 介导的转录抑制效率影响较大。因此,虽然PAM 远端的核苷酸为向导序列设计提供了一定的灵活性,但截短或错配均会降低CRISPR/dCas9 介导的转录调控的效率。另外,Smith 等[55]利用dCas9-Mxi1 对靶向1 631 个基因的超过18 000 个gRNA 的筛选实验发现,gRNA 的靶向效率还与其二级结构相关(斯皮尔曼相关性系数为-0.11),即gRNA 中向导序列、骨架序列和PRPR1启动子前导序列(Leader sequence)之间的碱基配对会降低gRNA 的靶向效率,因此在设计gRNA 时应尽量避免。
1.3.2 gRNA 靶向的DNA 单链和位点 gRNA 靶点也会对CRISPR/dCas9 的转录调控效率产生影响,这些因素包括不同的DNA 链、基因结构域、靶点与TATA 框(TATA box)或转录起始位点(Transcription start site,TSS)的距离等。在酿酒酵母中,Farzadfard 等[21]将dCas9-VP64 靶向不同DNA 链的相近位置,发现二者具有相似的转录调控水平。高通量的gRNA 文库筛选研究也表明,gRNA 的效率并没有DNA 链的偏好性[33,42]。在作用机制方面,dCas9-gRNA 复合体不仅是作为转录过程的路障,更是共同建立了促进转录起始或终止的新环境[56]。
与DNA 链特异性效应不同,gRNA 靶点的位置效应非常明显(图2-B)。对于dCas9-VP64 或VPR等介导的转录激活,通常将gRNA 靶向TATA 框和TSS 的上游;而对于dCas9 融合蛋白介导的转录抑制,通常将gRNA 靶向TATA 框附近或其下游[21,37]。在酿酒酵母中当gRNA 靶向TSS 上游约50-400 bp 区域时dCas9-VPR 介导的转录激活效率较高,与哺乳动物细胞中dCas9-SunTag:scFV-VP64 的实验结果一致[33,37]。当dCas9-VPR 靶向TATA 框上游50 bp之内时,可能由于抑制了RNA 聚合酶与靶基因的结合,转录激活的效率急剧下降[37]。值得注意的是,当dCas9-VPR 靶向开放阅读框(Open reading frame,ORF)区域时,反而起到了转录抑制的作用[18]。另一方面,Smith 等利用dCas9-Mxi1 测试了靶向20个酿酒酵母基因的约1 000 个gRNA。结果显示,gRNA 靶点位于TSS 上游200 bp 范围时转录抑制效率较高[42],进一步对超过1 500 个基因的筛选将该区域缩小到TSS 上游125 bp 范围[55]。与之相比,Gilbert 等[33]发现哺乳动物细胞中dCas9-KRAB 依赖的最大转录抑制效率的靶点区域为TSS -50 到+300 bp。利用gRNA 靶点位置效应的特性,Deaner 和Alper 综合利用dCas9-VPR 和dCas9-Mxi1 对目标基因表达进行不同程度的下调和上调,从而实现了基因转录的梯度调控(Graded modulation)[37]。例如,针对PGPD1启动子,调控后报告荧光蛋白yECitrine的表达水平是对照组的0.2、0.6、0.7、3、4 和8.5倍,动态范围超过40 倍。然而,Jensen 等[38]在TSS 上游200 bp 范围内设计了靶向12 个酿酒酵母启动子的88 个gRNA,其中一些gRNA 对dCas-VPR或Mxi1 介导的转录激活或抑制是无效的。因此,在选择CRISPR/dCas9 系统的靶点时,除了宿主细胞的不同,还需在合适区域内设计多个gRNA 进行具体的测试。
1.3.3 gRNA 靶点的可接近性 除了gRNA 序列的本体特征和gRNA 靶点所处的基因结构域外,gRNA 靶点的可接近性也是CRISPR/dCas9 系统有效发挥转录调控作用的重要因素,如染色质高级结构和核小体占据等(图2-C)。酿酒酵母等真核生物中染色质以DNA 缠绕组蛋白形成的核小体形式存在并具有复杂和动态的高级结构[57-58]。之前研究表明,靶向同一个启动子临近位置的不同gRNA 具有不同的转录调控效率[21,38,42]。利用酿酒酵母高分辨率的核小体占据谱数据[59-60],Smith 等发现在TSS 上下游400 bp 范围内gRNA 的转录抑制效率与染色质可接近性呈正相关[42]。体外实验表明,核小体的占据抑制了Cas9 蛋白与PAM 的相互作用,当在体系中加入染色质重塑蛋白SNF2h 或RSC 复合物后,该抑制作用被削弱[61-62]。特别的是,在人类细胞系中CRISPR/dCas9 依赖的转录抑制研究表明,有效的gRNA 定位还与开放的染色质环境和转录活跃相关的组蛋白修饰如H3K9ac、H3K27ac、H3K4me2、H3K4me3、H3K79me2 等有关[63-64]。综上,基于CRISPR/dCas9转录调控的效率受到靶点附近染色质微环境的影响,设计gRNA 时应参考核小体占据和DNA 可接近性的高分辨率图谱[59-60,65-66],避免靶向核小体DNA 的核心区域。
1.3.4 gRNA 的数量 与dCas9 融合多个转录调控蛋白结构域或gRNA 融合多个适配子的思路类似,靶向同一目的基因的多条gRNA 能够协同调控靶基因的转录(图3-A)。Farzadfard 等[21]组合使用两条gRNA 时将转录抑制的效率从2 倍提高到了7 倍,Deaner 等[18]同样利用两条gRNA 提高了dCas9-VPR介导的转录抑制水平。此外,还可以通过在启动子内人工合成相同的靶序列,实现多条单一gRNA介导的协同调控。Farzadfard 等[21]利用含有12 个gRNA 结合位点的合成型启动子,将dCas9-VP64 介导的转录激活提高了70 倍,Gilbert 等[23]利用含有7 个Tet 操纵子结合位点的合成型启动子,将dCas9介导的转录抑制提高了115 倍。
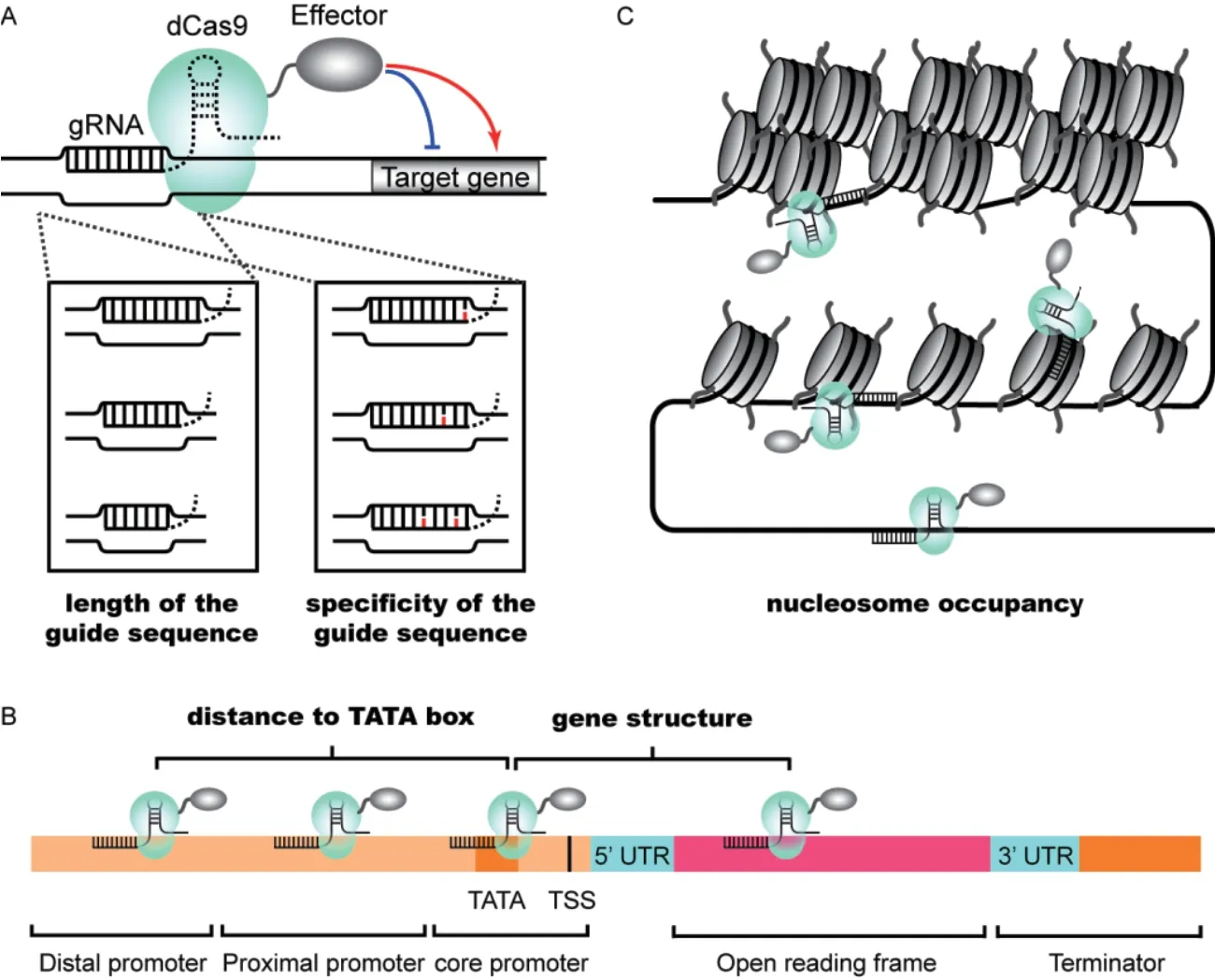
图2 影响CRISPR-dCas9 系统靶向的因素
利用靶向多个目的基因的多条gRNA 或scRNA可实现多重性(Multiplex)的转录调控(图3-B)。Jensen 等[38]利 用dCas9 与scRNA 将Mxi1 和VPR等转录调控结构域靶向类胡萝卜素和三酰甘油生物合成途径的多个基因,显著增加了相关产物的产量。Ferreira 等[54]利用来自绿脓假单胞菌的核糖核酸内切酶Csy4,实现了PRPR1启动子驱动单条转录本中3 个gRNA 的转录和释放,实现了同时对OLE1,HGM1和ACS1等多个基因进行转录调控。此外,基于CRISRP/dCas9 工具中组成元件相互作用的特异性,可以设计彼此正交的gRNA,在同一细胞中对多个目标基因实现不同模式的基因调控。例如,利用不同RNA 适配子与其结合蛋白的特异性识别,Zalatan 等[39]设计彼此正交、与不同转录调控因子结合的scRNA 序列(图3-B),在同一菌株中对紫色杆菌素合成通路的不同分支的基因进行转录抑制和转录激活,通过调控代谢流实现了不同产物的定向合成。利用不同Cas 蛋白识别不同PAM 序列的特点,Lian 等[19]测试了不同来源的dCas 蛋白和多种转录激活或抑制结构域的组合,构建了基于dLbCpf1-VP、dSpCas9-RD1152 和SaCas9 的酿酒酵母三相基因调控策略,称为CRISPR-AID(Transcriptional Activation,transcriptional Interference,and gene Deletion)。作者设计了彼此正交、与不同Cas 蛋白对应的gRNA 序列(图3-C),在同一细胞中可以实现对多个目的基因进行转录抑制、转录激活、基因敲除等不同模式的调控。综上,多重gRNA 的设计与使用能够提高单个靶基因的转录调控能力,实现多个靶基因的单向多重调控,还能够利用正交性设计系统实现靶基因的多重和多模式调控。通过增加可同时操作的基因靶点和调控模式,有助于从多个工程化改造靶点中快速找到最优的调控策略及其组合,即利用组合式优化为复杂基因线路、合成途径和细胞代谢的设计和改造提供有力工具。
2 酿酒酵母中CRISPR/dCas9 工具在代谢工程中的应用
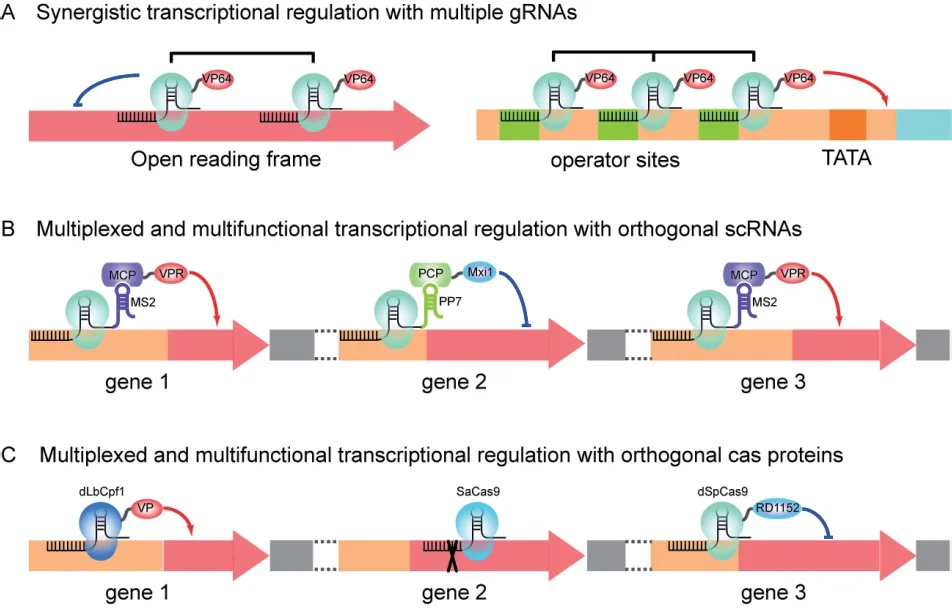
图3 利用多条gRNA 进行协同、多重或多模式转录调控
在酿酒酵母细胞工厂的优化过程中,平衡内源或外源代谢通路的基因表达水平是提高目的产物产量的重要手段。CRISPR/dCas9 转录调控工具可对多重基因的表达进行精细和多模式调控,从而通过代谢工程的理性设计和代谢靶点的高通量筛选等手段优化酿酒酵母的物质和能量流动,提高目的产物的产量和效率等。
首先,利用CRISPR/dCas9 转录调控工具的多重基因靶向和多模式调控的优点,可对酿酒酵母代谢通路多种关键酶的基因表达进行理性设计。例如,上调或下调甲羟戊酸(Mevalonate,MVA)途径的关键基因HMG1、ERG9和ERG20等,能够提高类异戊二烯的产量[67-68]。Jensen 等[38]利用dCas9 和scRNA 将转录调控结构域Mxi1 或VPR 靶向HMG1、HMG2、ERG9、ERG20、PGK1或TDH3等基因的启动子区域,通过测试多种转录调控的组合提高了类胡萝卜素的产量。而Lian 等[19]利用CIRSPR-AID方法对酿酒酵母中甲羟戊酸途径限速酶基因HMG1的表达进行了上调,对处于β-胡萝卜素和甾醇生物合成分支点的必需基因ERG9的表达进行了下调,同时敲除了转录调节因子ROX1基因,使得β-胡萝卜素的产量提高了2.8 倍。另外,Jensen 等[38]将dCas9-VPR 同时靶向OLE1和DGA1基因的启动子区域,使得三酰基甘油的产量提高了2 倍以上。
其次,揭示基因型与表型的关系仍是代谢工程的一大挑战,由于具有易于设计使用和高通量的优点,CRISPR/dCas9 可用于代谢通路靶基因表达强度与表型关系的的高通量筛选和研究。例如,Deaner和Alper 利用dCas9-VPR 或dCas9-Mxi1 以及靶向启动子不同位置的gRNA 文库建立了一种梯度式的基因转录调控工具,实现不同水平的基因表达上调和下调。他们将该方法用于系统性测试代谢通路中不同催化步骤对于调控干扰的敏感性(Systematically Test Enzyme Perturbation Sensitivities,STEPS), 构建了甘油、3-脱氢莽草酸发酵和木糖分解的代谢通量敏感性图谱(Flux sensitivity map)[37]。通过对通量敏感性图谱进行迭代分析,依次鉴定了甘油发酵中GPD1、TPI1等基因编码的酶是合成通路中的限速步骤,并通过工程化改造菌株使甘油的产量从4.89g/L 提高至28g/L。通过类似的方法,又将3-脱氢莽草酸的产量提高至126.4mg/L。Ferreira 等[69]利用dCas9-VPR 和靶向168 个基因的3194 个gRNA 组成的文库,结合基于转录因子FapR 的丙二酰辅酶A生物传感器,利用流平衡分析筛选了促进代谢流向丙二酰辅酶A 的靶基因和gRNA。利用筛选的结果,使3-羟基丙酸的产量得到显著提高。
如上所述,由于具有多重靶基因多模式转录调控和易于高通量等优势,基于CRISPR/dCas9 的基因转录调控工具在代谢靶点筛选、机制研究和代谢通路的工程化改造等方面具有广泛的应用,是基因编辑工具的重要补充。随着该工具的进一步开发和优化,基于CRISPR/dCas9 的转录调控工具定能在微生物细胞工厂及其它领域有更加广泛的应用。
3 结论与展望
综上所述,CRISPR/dCas9 系统在酿酒酵母中提供了一种高效率和可编程的靶向基因转录调控策略。该策略包括如下优点:(1)gRNA 易于设计和使用;(2)可实现多重靶基因转录调控;(3)可通过改变gRNA 靶点的具体位置等实现梯度式基因转录调控;(4)可通过组合正交性的scRNA 或dCas 系统同时实现基因转录的激活和抑制。但是,目前酿酒酵母中CRISPR/dCas9 介导的转录调控的效率普遍仍比大肠杆菌或哺乳动物细胞偏低[13,23,36],相关调控工具还需要进一步开发和优化。
就dCas 元件而言,可以扩展dCas 蛋白的类型、物种来源以及转录调控结构域类型等。除了广泛使用的SpCas9 外,Lian 等[19]测试了其它物种来源的Cas9 蛋白,如NmCas9、St1Cas9、SaCas9,以及两种物种来源的Cpf1,即AsCpf1、LbCpf1,并系统的比较了它们在基因编辑、转录抑制或激活方面的性能。这不仅扩大了PAM 位点的选择范围,还为转录调控的优化提供了借鉴。此外,除了常用的Mxi1、KRAB 和VPR 外,Lian 等测试并发现来源于UME6、MIG1 和TUP1 的结构域组合具有更强的转录抑制作用。Keung 等[70]基于可编程的锌指蛋白测试了213种酿酒酵母染色质调节因子,并对它们的转录调控作用模式进行了系统分析,为多样化转录调控结构域提供了参考。而且,DNA 甲基化、组蛋白修饰等表观遗传学机制同样在基因转录等生物学过程中起着多样化而且重要的作用[71]。利用dCas9 与表观遗传修饰酶全长或活性结构域的融合进行靶向表观遗传修饰的编辑,可对基因表达进行调控以实现特定的生物学效应,在深化理解哺乳动物细胞的生物学过程、开发基因治疗或细胞治疗的工具中有广泛应用[72-74]。酿酒酵母作为单细胞真核生物,有丰富的组蛋白修饰和组蛋白修饰酶,并广泛参与到DNA 转录调控中[75]。然而,虽然在哺乳动物中通过表观遗传编辑进行转录调控已有很多应用案例,但在酿酒酵母中的研究仍十分匮乏,这也为酿酒酵母转录调控的表观遗传编辑工具开发和应用指明了一个重要方向。
就gRNA元件而言,虽 然CHOPCHOP[76]等计算机辅助设计工具被应用于转录调控gRNA 的设计当中,但相关算法以基因编辑为核心,对CRISPR/dCas9 介导转录调控效率的影响因素并未有充分考虑。针对这一局限,Smith 等[42]开发了用于酵母CRISPRi 的gRNA 设计工具(http://lp2.github.io/yeast-crispri/),算法原理涉及基因组位置、染色质可接近性、核小体占据、gRNA 长度与序列和转录因子占据等。未来的设计算法开发还应综合考虑各类转录调节结构域及结合方式、不同dCas 蛋白及PAM 序列、靶点染色质微环境、菌株背景等因素,且需要依据全基因组范围大规模筛选等实验结果进行优化。目前,dCas9 转录激活在酿酒酵母中尚无gRNA 辅助设计工具。另外值得注意的是,通常CRISPR/dCas9 系统的转录激活效率与靶基因的基础表达呈负相关,即靶基因基础表达水平越高,对其转录增强的效率越低[36,38]。因此,还需针对每个靶基因设计特异性的、更为有效的转录调控方法。
