运输胁迫对大菱鲆生理免疫影响的初步探究
2020-03-23马壮田涛吴忠鑫
马壮 田涛 吴忠鑫

摘 要:大菱鲆(Scophthalmus maximus)是我國北方的重要经济鱼种,为研究运输胁迫对大菱鲆生理免疫的影响,并确定大菱鲆的优选运输方案,采用体长为4.65±1cm(幼体组)和14.50±1cm(成体组)2种规格的大菱鲆为研究对象,在实验室模拟运输条件,分析其非特异性免疫、呼吸排泄及死亡率变化,并探究活鱼苗运输的优选方案,结果显示:(1)过氧化氢酶(CAT)、碱性磷酸酶(AKP)、超氧化物歧化酶(SOD)、溶菌酶(LZM)呈现降低趋势并明显低于对照组,随着模拟运输中温度的升高、密度的增加和震动强度的增加而降低,谷草转氨酶(GOT)、谷丙转氨酶(GPT)、丙二醛(MDA)呈现升高趋势并明显高于对照组,随着模拟运输中温度的升高、密度的增加和震动强度的增加而升高;(2)大小2种规格的大菱鲆在运输胁迫下表现出了不同的免疫能力,幼体组中的GOT、GPT、SOD、MDA的表现优于成体组,成体组中的CAT、AKP、LZM优于幼体组;(3)耗氧率和排氨率随着模拟运输中温度的升高、密度的增加和震动强度的增加而升高,幼体组的耗氧率和排氨率的升高程度要低于成体组;(4)实验结束后的幼体组与成体组大菱鲆死亡率均随时间变化先升高后降低,在实验结束时的死亡率极低,而在0~24h死亡率达总死亡率的50%以上,24~48h内的死亡率又开始下降。随着模拟运输中温度的升高、密度的增加和震动强度的增加,2种规格的大菱鲆的总死亡率呈现上升趋势,成体组的总死亡率的升高程度要低于幼体组;(5)通过正交实验极差分析法得出综合考虑成本及运输效果前提下,幼体组与成体组大菱鲆的优选运输方案均为A1B1C2,即温度为15℃、密度为ind·L-1、震动强度为200RPM。
关键词:大菱鲆;非特异性免疫;运输胁迫;温度;密度;震动强度
中图分类号 S981.14文献标识码 A文章编号 1007-7731(2020)04-0074-10
Abstract:Turbot (Scophthalmus maximus) is an important economic fish species in the north of China. In order to study the effect of transportation stress on the physiological immunity of turbot,and to determine the optimal transportation scheme of turbot,the turbot with the body length of 4.65±1 cm (juvenile group) and 14.50±1 cm (adult group) was used as the research object,and the transportation conditions were simulated in the laboratory,and explore the optimal transportation plan,analyze the changes in non-specific immunity,respiratory excretion and mortality,the results show:(1) CAT,AKP,SOD,LZM showed a decreasing trend and were significantly lower than those of the control group. With the increase of temperature,density and vibration intensity during simulated transportation,GOT,GPT,MDA showed an increasing trend It is significantly higher than the control group,and increases with the increase in temperature,density,and vibration intensity during simulated transportation;(2) Two groups of turbot showed different immune abilities under transportation stress. The performance of GOT,GPT,SOD,and MDA in the juvenile group was better than that in the adult group. CAT,AKP,LZM in the adult group Better than juvenile group;(3) The oxygen consumption rate and ammonia excretion rate increase with the increase of temperature,density,and vibration intensity during the simulated transport,and the increase of the oxygen consumption rate and ammonia excretion rate of the juvenile group is lower than Adult group;(4) After the experiment,the mortality of juvenile and adult turbot increased and then decreased with time. The mortality at the end of the experiment was extremely low,and the mortality reached 0-24 hours. More than half,the mortality rate within 24-48 hours began to decline again. With the increase of temperature,density and vibration intensity in simulated transportation,the total mortality rate of turbot with two specifications showed an increasing trend,and the increase of total mortality rate in the adult group was lower than that in the juvenile group;(5) Based on the orthogonal experimental range analysis method,it is concluded that the preferred transportation schemes for juvenile and adult groups of turbot are A1B1C2 under the premise of comprehensive consideration of cost and transportation effect,that is,the temperature is 15 ° C,the density is 40 ind·L-1,the vibration intensity is 200 RPM.
Key words:Scophthalmus maximus;Non-specific immunity;Transport stress;Temperature;Density;Vibration intensity
陆地活鱼运输是现在水产品由滨海城市输送到内地的最主要方式之一[1]。其中运输方法有很多,包括传统的密闭式运输法、无水湿法运输、干法运输、麻醉运输等,而苗种和较小规格活鱼的运输则较多采用低成本高效的密闭式运输法[2],但无论哪种运输方式都会使鱼产生相应程度的应激反应[3]。在此过程中机体的免疫力防御机制会受到抑制,导致鱼类感染各种病原的可能性增加[4],同时在运输中的高密度、温度升高和道路颠簸产生的胁迫作用也会对鱼类的免疫和生存产生影响。运输活鱼的方法和运输中不利条件对鱼类生理上造成的影响已为许多著作所证实[5]。国内外已经对草鱼[6](Ctenopharyngodon idellus)、美洲鲥[7](Alosa sapidissima)、鳜鱼[3](Siniperca chuatsi)、日本黄姑鱼[8](Nibea japonica)等进行了运输胁迫的相关研究,但关于大菱鲆运输胁迫的相关报道相对较少。
大菱鲆(Scophthalmus maximus)又被称为鲽鱼,属于脊索动物门硬骨鱼纲,鲽形目鲆科[9],是目前品质较好的养殖品种之一。鱼体的蛋白质含量非常高[10],味道鲜美,对于美容养颜具有非常好的效果,除此之外可以补肾健脑,提高精气,在我国很受欢迎,有“多宝鱼”的美誉,1992年由黄海水产研究所从英国引进[11]。据不完全统计,我国的大菱鲆总产量已超出其他国家同期水平,养殖面积成倍增加,现已成为我国北方水产经济养殖的重要支柱。为探究最适的大菱鲆的活鱼运输方案,本研究通过室内模拟车辆运输条件,分析鱼体免疫指标、呼吸排泄等变化,结合运输后死亡率,得出最适的运输条件,确定优选运输方案,以期为选择适宜的活鱼运输条件提供参考依据。
1 材料与方法
1.1 实验材料 实验用鱼于2018年4月30日从大连天正实业有限公司采购,平均体质量分别为10.36±5g(幼体组)和50.30±5g(成体组),平均体长分别为4.65±1cm(幼体组)和14.50±1cm(成体组)。实验时挑选体色正常、行动迅速、生命力强的个体用于胁迫实验。实验前在循环养殖水槽中暂养,水温13℃,盐度31‰,24h供氧。每天早晚各投喂1次,投饵量为鱼体重的2%~3%,每天换水1次,换水量为总水量的2/3,采用虹吸的方式清除残饵和粪便并保持池内的清洁。
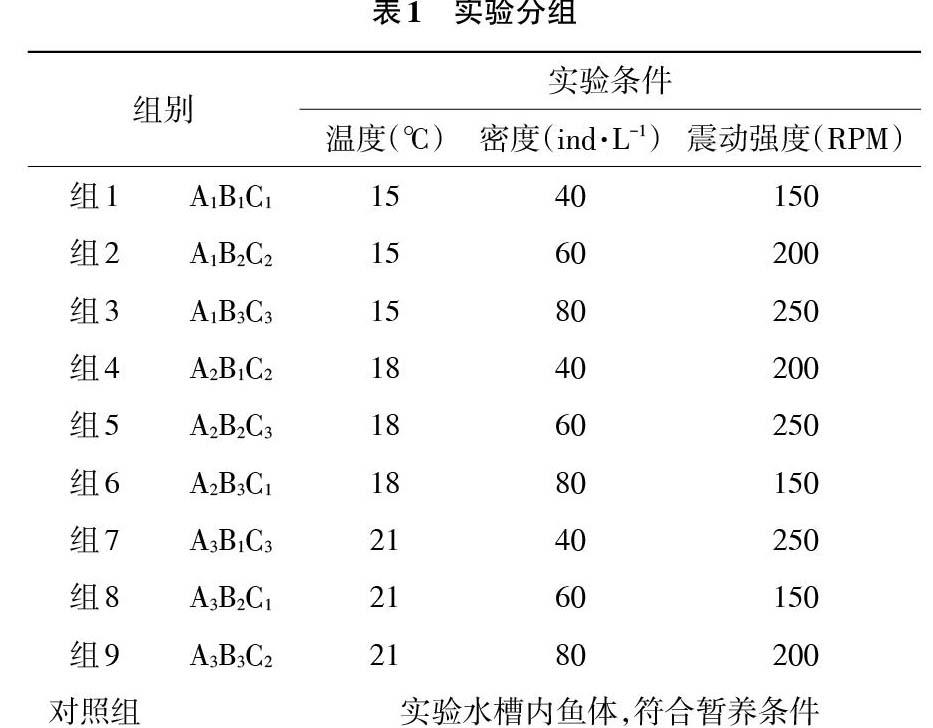
1.2 模拟运输 实验采用活鱼运输专用双层塑料运输袋,每袋加入过滤清洁海水并放入实验鱼,模拟实际运输条件,充纯氧后打包放入泡沫箱中进行运输实验。实验分为2种规格的大菱鲆进行,实验采用正交实验设计,控制温度、密度和震动强度的变化,分9组进行,重复3次试验,分组情况见表1。
1.3 样品采集和处理
1.3.1 样品采集 在运输实验结束之后,将实验用鱼迅速捞起并立即投入MS-222浓度为200mg·L-1的海水中做快速深度麻醉,避免因人为刺激而引起的应激反应。麻醉后立刻取肝脏,进行组织破碎后加入生理盐水配成10%浓度的组织液,离心(3000R,10min)取上清液,最后把采集的样品保存在4℃冰柜中保存,并在24h内测量完毕。
1.3.2 酶指标的测定 实验测定的溶菌酶(LZM),碱性磷酸酶(AKP),谷草转氨酶(GOT),过氧化氢酶(CAT),谷丙转氨酶(GPT),总超氧化物歧化酶(T-SOD),丙二醛(MDA)共计7种酶采用试剂盒测定,试剂盒从南京建成生物科技有限公司采购。
1.3.3 耗氧率与排氨率的测定 在進行每组实验时,分别保存实验前后的水体样品(-20℃冷冻保存,在24h内分别利用碘量法和靛酚蓝法对溶解氧和水中氨氮进行测定),耗氧率和排氨率公式为:
式中:V—水体体积,L;W—个体体重,g;t—试验时间,h。R0—单位体重耗氧量,mg·(g·h)-1;RA—单位体重排氨率,μg·(g·h)-1;DO0、DOt—分别为试验开始和结束时的水体溶氧浓度,mg·L-1;Nt、N0—分别为实验开始和结束是的水体氨氮浓度,μg·g-1。
1.3.4 死亡率 在实验结束后0h、24h、48h分别计数大菱鲆死亡数量,其公式为:
CDR(%)=(N1/N2)×100
式中CDR为死亡率,N1为死亡个体数量,N2为种群总数量。
1.3.5 正交实验极差分析 Rj反映了第j列因素的水平变动时,试验指标的变动幅度。Rj越大,说明该因素对试验指标的影响越大,因此也就越重要。极差Rj为第j列因素的极差,即第j列因素各水平下平均指标值的最大值与最小值之差,于是依据Rj的大小,就可以判断因素的主次。
极差Rj计算公式为:
式中ym为各实验水平的试验指标值,Kjm为第j列因素m水平所对应的试验指标和,[Kjm]为Kjm的平均值。由Kjm的大小可以判断j因素的优水平和各因素的水平组合,即最优组合。
1.4 数据分析 所得的全部数据均利用SPSS 17.0软件对测定结果进行统计分析,采用方差分析和Tukey检验进行显著性分析和多重比较,数据在P<0.05时被认为具有显著性差异。
2 结果与分析
2.1 模拟运输条件对大菱鲆非特异性免疫的影响 由表2、图2-A可以看出,随着模拟运输条件的加剧,2种规格的大菱鲆体内CAT活性显著降低(P<0.05),并且成体组体内AKP酶活性的降低程度要较低于幼体组,两者具有显著差异性(P<0.05)。由表2、图2-B可以看出,随着模拟运输条件的加剧,2种规格的大菱鲆体内AKP活性显著降低(P<0.05),并且成体组体内AKP酶活性的降低程度要较低于幼体组,两者具有显著差异性(P<0.05)。由表2、图2-C、2-D可以看出,随着模拟运输条件的加剧,2种规格大菱鲆体内的GOT和GPT活性呈现正相关,显著高于对照组(P<0.05),而幼体组体内GOT和GPT酶活性的上升程度较低于成体组,两者具有显著差异性(P<0.05)。由表2、图2-E、2-F可以看出,随着模拟运输条件的加剧,两种规格的大菱鲆体内SOD活性显著降低(P<0.05),MDA含量显著升高(P<0.05),而幼体组体内SOD的降低程度和MDA含量的上升程度较低于成体组,两者具有显著差异性(P<0.05)。
由表2可知,这7种酶指标的极差数Rj值,因此可以判断温度、密度和震动强度对大菱鲆非特异性免疫的影响的主次要关系。在CAT活性酶中,幼体组的R1=10.68>R2=2.51>R3=0.54,成体组的R1=11.02>R2=2.24>R3=1.06,因此温度为主要影响因素,密度为次要影响因素,震动强度的影响能力最弱。在AKP活性酶中,幼体组的R1=10.05>R2=3.23>R3=0.53,成体组的R1=7.52>R2=1.42>R3=0.79,因此温度为主要影响因素,密度为次要影响因素,震动强度的影响能力最弱。在GOT活性酶中,幼体组的R1=8.14>R2=1.09>R3=1.09,成体组的R1=18.82>R3=2.04>R2=1.25,因此温度为主要影响因素,密度为幼体组的次要因素,震动强度为成体组的次要影响因素。在GPT活性酶中,幼体组的R1=11.92>R2=2.00>R3=0.58,成体组的R1=13.33>R2=2.11>R3=0.64,因此温度为主要影响因素,密度为次要影响因素,震动强度的影响能力最弱。在SOD活性酶中,幼体组的R1=6.06>R2=1.05>R3=0.51,成体组的R1=5.19>R3=2.19>R2=1.03,因此温度为主要影响因素,密度为幼体组的次要影响因素,震动强度为成体组的次要影响因素。在MDA含量中,幼体组的R1=4.37>R2=1.55>R3=0.52,成体组的R1=9.96>R2=2.15>R3=0.54,因此温度为主要影响因素,密度为次要影响因素,震动强度的影响能力最弱。在LZM活性酶中,幼体组的R1=0.25>R2=0.08>R3=0.05,成体组的R1=0.18>R2=0.06>R3=0.02,因此温度为主要影响因素,密度为次要影响因素,震动强度的影响能力最弱。
2.2 模拟运输条件下大菱鲆的呼吸和代谢 由表3、图3-A可以看出,随着模拟运输条件的加剧,2种规格的大菱鲆的耗氧率显著升高(P<0.05),并且幼体组的耗氧率的升高程度要低于成体组,两者具有显著差异性(P<0.05)。由表3、图3-B以看出,随着模拟运输条件的加剧,两种规格的大菱鲆的排氨率显著升高(P<0.05),并且幼体组的排氨率的升高程度要低于成体组,两者具有显著差异性(P<0.05)。
由表3可知这耗氧率和排氨率的极差数Rj值,因此可以判断温度、密度和震动强度对大菱鲆呼吸代谢的影响的主次要关系。在耗氧率中,幼体组的R1=0.041>R3=0.025>R2=0.010,成体组的R1=0.067>R2=0.014>R3=0.003,因此温度为主要影响因素,震动强度为幼体组的次要影响因素,密度为成体组的次要影响因素。在排氨率中,幼体组的R1=0.219>R2=0.051>R3=0.33,成体组的R1=0.320>R2=0.074>R3=0.031,因此温度为主要影响因素,密度为次要影响因素,震动强度的影响能力最弱。
表3 大菱鲆的耗氧率和排氨率
[分组 耗氧率(mg·(g·h)-1) 排氨率(μg·(g·h)-1) 幼体组 成体组 幼体组 成体组 组1 A1B1C1 0.134b 0.091c 0.894c 0.916e 组2 A1B2C2 0.087d 0.096c 0.774c 0.983e 组3 A1B3C3 0.091d 0.108c 0.831c 1.017d 组4 A2B1C2 0.099d 0.124b 0.891bc 1.123c 组5 A2B2C3 0.114c 0.137b 0.919b 1.142c 组6 A2B3C1 0.130b 0.129b 0.942b 1.153c 组7 A3B1C3 0.131b 0.153a 1.023ab 1.249b 组8 A3B2C1 0.148a 0.168a 1.030a 1.286b 组9 A3B3C2 0.157a 0.174a 1.101a 1.341a Ka1 0.311 0.295 2.499 2.916 Ka2 0.342 0.390 2.752 3.418 Ka3 0.435 0.495 3.154 3.876 Kb1 0.364 0.368 2.808 3.288 Kb2 0.348 0.401 2.722 3.411 Kb3 0.377 0.411 2.875 3.511 Kc1 0.411 0.388 2.866 3.355 Kc2 0.342 0.394 2.766 3.447 Kc3 0.336 0.398 2.773 3.408 R1 0.041 0.067 0.219 0.320 R2 0.010 0.014 0.051 0.074 R3 0.025 0.003 0.033 0.031 ]
图3 大菱鲆的耗氧率和排氨率
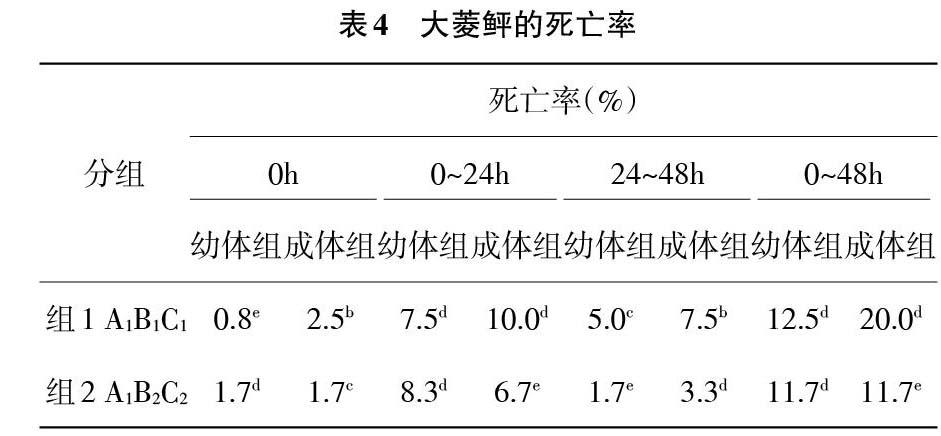
2.3 模拟运输实验后大菱鲆的死亡率 由表4、图4随着模拟运输条件的加剧,2种规格的大菱鲆的总死亡率呈现上升趋势,成体组的总死亡率的升高程度要低于幼体组,两者差异性显著(P<0.05)。0h、0~24h和24~48h大菱鲆的死亡率呈现出先升高后降低的趋势,并且成体组在各个时间段的死亡率均高于幼体组(P<0.05)。
由表4可知死亡率的极差数Rj值,因此可以判断温度、密度和震动强度对大菱鲆死亡现象的影响的主次要关系。在0h,幼体组的R1=0.049>R3=0.013>R2=0.011,成体组的R1=0.028>R2=0.008>R3=0.003,因此温度为主要影响因素,震动强度为幼体组的次要影响因素,密度为成体组的次要影响因素。在0~24h,幼体组的R1=0.158>R2=0.046>R3=0.17,成体组的R1=0.167>R2=0.036>R3=0.031,因此温度为主要影响因素,密度为次要影响因素,震动强度的影响能力最弱。在24~48h,幼体组的R1=0.068>R3=0.31>R2=0.024,成体组的R1=0.044>R2=0.033>R3=0.022,因此温度为主要影响因素,密度为成体组的次要影响因素,震动强度为幼体组的次要影响因素。在0~48h,幼体组的R1=0.275>R3=0.053>R2=0.046,成体组的R1=0.239>R2=0.067>R3=0.010,因此温度为主要影响因素,密度为成体组的次要影响因素,震动强度为幼体组的次要影响因素。
2.4 大菱鲆幼体组和成体组的优选运输方案
2.4.1 幼体组的优选运输方案 首先根据各指标下的平均数据和K1、K2、K3,初步确定各因素的最优水平组合(见表5),由于8个指标单独分析出来的最优条件并不一致,所以必须根据因素对3个指标影响的主次顺序,综合考虑,确定出最优条件。
温度在各指标中均为主要影响因素,并且其在各指标中表现一致,均为15℃最优;密度在CAT、AKP、GOT、GPT、SOD、MDA、LZM中的影响都排在第2位为次要因素,而对0~48h死亡率的影响则排在第3位,属第3影响因素。根据表5可以看出在CAT、AKP、GOT、GPT、MDA、LZM中取B1为好,在SOD中取B2为好。当取B1为40ind·L-1时,CAT活性比取B2时提高10.075%,AKP活性比取B2时升高10.421%,GOT活性比取B2时降低1.476%,GPT活性比取B2时降低7.833%,MDA含量比取B2时降低8.201%,LZM活性比取B2时降低22.429%,而在取B2为60ind·L-1时,SOD活性比B1时升高0.578%,因此应该选择优势水平更大的B1水平。而震动强度则正好相反,其在0~48h死亡率中的影响都排在第2位,为次要因素,而对CAT、AKP、GOT、GPT、SOD、MDA、LZM的影响则排在第3位,属第三影响因素。根据表5可以看出在,在0~48h死亡率中取C2为好,因此应该选择C2水平。
综上所述,幼体组大菱鲆的优选运输方案为A1B1C2,即温度为15℃、密度为40ind·L-1、震动强度为200RPM。
2.4.2 成体组的优选运输方案 首先根据各指标下的平均数据和K1、K2、K3,初步确定各因素的最优水平组合(见表6),由于8个指标单独分析出来的最优条件并不一致,所以必须根据因素对3个指标影响的主次顺序,综合考虑,确定出最优条件。
温度在各指标中均为主要影响因素,并且其在各指标中表现一致,均为15℃最优;密度在CAT、AKP、GPT、MDA、LZM和0~48h总死亡率中的影响都排在第2位为次要因素,而对GOT和SOD的影响则排在第3位,属第3影响因素。根据表6可以看出在CAT、AKP、MDA、LZM中取B1为好,在GPT中取B2为好,而在0~48h总死亡率取B3为好。当取B1为40ind·L-1时,CAT活性比取B2时提高12.729%,比取B3时升高16.019%,AKP活性比取B2时升高6.488%,比取B3时升高11.490%,MDA含量比取B2时降低6.987%,比取B3时降低16.483%,LZM活性比取B2时降低9.302%,比取B3时降低19.767%;而在取B2为60ind·L-1时,GPT活性比B1时升高0.578%,比取B3时升高10.149%;在取B3为80ind/L时,0~48h总死亡率比B1时降低27.586%,比取B2时降低1.149%。因此应该选择优势水平更大的B1水平。而震动强度则正好相反,其在GOP和SOD中的影响都排在第2位,为次要因素,而对CAT、AKP、GPT、MDA、LZM和0~48h总死亡率的影响则排在第3位,属第3影响因素。根据表6可以看出在,在GOT中取C3为好,在SOD中取C2为好。在取C2为200RPM时,SOD活性比C3升高12.502%,在取C3为250RPM时,GOT活性比C2升高5.372%,因此应该选择优势水平更大C2水平。
综上所述,成体组大菱鲆的优选运输方案同样为A1B1C2,即温度为15℃、密度为40ind·L-1、震动强度为200RPM.
3 讨论
3.1 模拟运输条件对大菱鲆非特异性免疫的影响 一般而言,温度升高能够促进鱼类的生长发育,但当温度超过一定限度时,会影响其生长,甚至会引发死亡[12]。温度对鱼类生长的影响具有两面性,一方面温度升高导致鱼体的能量代谢加快,消耗的能量增加;另一方面,温度升高可以促进鱼体摄入能量转化为自身的净能量[13]。酶在动物体内起到促进新陈代谢的作用,也是在动物体内的物质代谢中起到关键作用,血清酶一般存在于动物的各种组织器官中,相应组织器官的代谢水平和功能状态影响着酶活性的高低,各组织器官的机能水平也影响着机体的调节和适应能力[14]。Cheng等[15]研究发现斜带石斑鱼(Epinepheluscoioides)在高于最适饲养温度时免疫力明显受到抑制。
而高密度养殖会对水产业的产量和成本提供有利的帮助,但是当密度的逐渐增加时,养殖种群内个体对资源、空间的竞争加剧,产生高压环境,导致动物生理和免疫功能等方面产生不利的变化,最终可能会降低动物的生存能力[16-17]。对拥挤胁迫的应激反應在生理上的表现为血液中白细胞和红细胞数量发生变化,肝脏和脾脏指数明显降低,免疫功能受到抑制[18]。强俊等[19]发现短期高密度胁迫可使尼罗罗非鱼(Oreochromis niloticus)血清葡萄糖与甘油三酯提高,并诱发肝损伤。
运输胁迫会使鱼体内的多项生理生化指标发生变化。震动强度则是由于运输过程中车辆行驶带来的颠簸,是活鱼运输中不可避免的影响因素之一。车辆的颠簸经过传导,最终致使运输鱼类的水体也产生震荡,对其中的鱼类产生影响。大量的研究表明运输胁迫会使鱼类的生理指标发生变化。Brian等发现运输后的尖吻鲈(Lates calcarifer)的红细胞出现变形膨胀,血液pH值降低且影响血液输氧能力[20]。在运输条件下,牙鲆的肌肉氨基酸组成发生变化,体内ATP含量下降,乳酸含量升高[21];头石脂鲤(Brycon cephalus)幼鱼血糖含量升高[22];大西洋鲑鱼(Salmo salar)血浆肾上腺素、血液乳酸和血糖合量都有明显变化[23];红鳟鱼(Salmo gairdneri)血液肾上腺素浓度增加[24]。
过氧化氢酶(CAT)是一种生物防御体系的关键酶,在机体内稳定的起到分解过氧化氢的作用,使过氧化氢分解成水和氧气,减少机体的-OH的积累,使细胞免于H2O2的破坏[25]。而实验组大菱鲆的CAT活性均出现下降趋势,表明大菱鲆体内清除超氧自由基、H2O2和过氧化物以及阻止或减少羟基自由基形成等方面的能力大幅度降低[26],这与徐冬冬等[13]将褐牙鲆(Paralichthys olivaceus)进行不同温度的高温实验结果相似。
碱性磷酸酶(AKP)是一种磷酸单酯酶,可催化磷酸单酯及磷酸基团的转移反应,并直接参与磷的代谢,与RNA、DNA、脂质和蛋白质的代谢有关[27],它也是巨噬细胞溶酶体中的非特异性磷酸水解酶,在血液中能防御外界微生物入侵[28]。而且AKP是一种特殊的膜结合蛋白,能够控制鱼体的代谢,还用于维持生物体内钙磷比相对稳定,并且能参与蛋白质和脂类的代谢[29]。AKP活力受生物体生长阶段、摄食状况、环境等因素影响而发生相应变化[30-31]。实验组2种规格的大菱鲆体内AKP活性均呈现下降趋势,导致鱼体相关代谢的紊乱,减弱鱼体对防御外界微生物入侵能力。
谷草转氨酶(GOT)和谷丙转氨酶(GPT)是反应鱼体肝脏健康情况的2种酶,活性高低和肝脏健康直接相关。2种转氨酶主要存在于细胞线粒体,尤以肝组织中最为丰富,可用于分析肝功能是否健康[32]。正常情况下,在肝组织中转氨酶活性值相对较小[33]。当GOT和GPT浓度上升或活性突然持续性增强时,表明组织中毒发生病变或者受损伤的组织范围较大[34]。实验组2种规格大菱鲆体内的GOT和GPT活性显著上升,说明运输胁迫对大菱鲆的肝脏造成损伤,并且这种损伤还在不断加剧。
超氧化物歧化酶(SOD)与丙二醛(MDA)一起反应了鱼类机体抗氧化能力的情况,SOD具有清除阴离子自由基的作用,是机体中免疫系统的重要组成部分[35],还能促进免疫细胞的吞噬能力,控制细胞膜脂质过氧化并保护机体正常的生理功能。SOD活性的高低反应了机体清除氧自由基的能力的高低,也表明鱼体对炎症的抵抗能力的强弱[35];MDA丙二醛是膜脂过氧化的产物之一,具有很强的毒性,因此MDA含量的高低与细胞受迫程度有很大的关系,MDA主要损伤生物膜结构,改变膜的通透性,影响生化反应,其含量侧面反映机体细胞受损程度[36]。试验组2种规格的大菱鲆体内SOD活性显著下降,MDA含量显著升高,其机体抗氧化能力大幅度下降,体内MDA迅速堆积。
溶菌酶(LZM),由巨型吞噬细胞分泌,并且广泛存在于鱼体血液及一些淋巴细胞中,具有抗菌、消炎、抗病毒等作用,是体液免疫的第一道防线[37-38]。实验组两种规格的大菱鲆体内LZM活性呈现下降,说明大菱鲆在运输条件下对细菌和病毒的清除能力受到较强的抑制,免疫防线受损,更容易受到感染,但由于运输水体封闭,水质较差,易滋生病菌,大菱鲆很有可能已经处于感染中。
3.2 模拟运输条件下大菱鲆的呼吸和代谢 在养殖过程中,大菱鲆的耗氧、氨氮排泄是要着重考虑的要素,而且耗氧率、氨氮排泄率也是反映大菱鲆生理状况的重要指标[39]。这些生理指标受温度、体重等的影响,国内外学者对成鱼做了大量的工作[40],并且普遍认为温度高低和耗氧率、氨氮排泄率成正比,这是变温动物的特性[41]。实验组2种规格大菱鲆耗氧率和排氨率的升高可以表明大菱鲆进入了应激反应状态,其机体正在抵抗运输胁迫带来的损伤。并且随水温的升高,水体中的饱和溶解氧含量降低,鱼体要获得等量的氧气其鳃部就要过滤更多的高温水[42],即在高温时鱼用于呼吸的能量将增加。过高的密度会导致种鱼类对空间竞争,运输带来的震动强度也会使鱼类处于运动状态,这些都是促使大菱鲆呼吸和代谢上升的原因。
3.3 模拟运输实验后大菱鲆的死亡率 在模拟运输试验后的0~48h内,大菱鲆出现了死亡现象。实验结束后的大菱鲆死亡率随着时间的增加先升高后降低,而大菱鲆在0h的死亡率极低,在0~24h里大菱鲆的死亡率达总死亡率的一半以上,之后在24~48h内的死亡率又开始下降。由此可见,运输胁迫对大菱鲆的生理免疫方面造成了较大的损伤,这种损伤会在结束后的48h内表现出来,并且大部分为不可逆损伤,直至引发了鱼体的死亡。
3.4 菱鲆幼体组和成体组的优选运输方案 由正交实验极差分析中综合平衡法分析得出:幼体组大菱鲆的最优运输方案为A1B1C2,即温度为15℃、密度为40ind·L-1、震动强度为200RPM;成体组大菱鲆的最优运输方案同样为A1B1C2,即温度为15℃、密度为40ind·L-1、震动强度为200RPM。虽然2种规格组在不同指标下的各因素的最优水平有差异,但根据分析方法分析得出的最优方案却是一致的,这表明温度、密度和震动强度3个因素对大菱鲆生理免疫方面的影响与这2种大菱鲆规格并没有较大关联性。此次正交实验对CAT、AKP、GOT、GPT、SOD、MDA、LZM和總死亡率共8种指标进行分析,未对耗氧率、排氨率和各时间段的独立死亡率进行分析,因为耗氧率和排氨率的变化不能单独的作为分析鱼体生理性能的参考,多种因素可以引起它们的变化,不能一概而论,需要具体分析,而各时间段的独立死亡率由总死亡率代替,为了避免死亡率的重复分析,故而未对其进行分析。
综上所述,运输胁迫对大菱鲆的生理免疫方面造成了影响,随着模拟运输条件的加剧,2种规格的大菱鲆体内这7种非特异性免疫酶呈现出上升和下降,均反应其免疫水平存在不同程度的下降。根据实验数据可以看出7种非特异性免疫酶在温度变化带来的损伤大于密度和震动强度变化带来的损伤。2种规格的大菱鲆在运输胁迫下表现出了不同的免疫能力,幼体组大菱鲆中的GOT、GPT、SOD、MDA的表现优于成体组的大菱鲆,成体组大菱鲆中的CAT、AKP、LZM和总死亡率的表现优于幼体组的大菱鲆。同时耗氧率和排氨率的升高也表明大菱鲆处于运输胁迫的应激反应中。通过正交实验分析,幼体组与成体组大菱鲆的最优运输方案一致,均为A1B1C2,即温度为15℃、密度为40ind·L-1、震动强度为200RPM。
本实验通过对大菱鲆的生理免疫、呼吸排泄等方面受到运输胁迫的影响进行了探究,探明了运输条件中各因子对大菱鲆的影响程度,并且确定了2种规格大菱鲆短途运输的最优方案。这为深入探究运输胁迫对大菱鲆的影响提供了基础数据,也为大菱鲆短途活鱼运输提供了参考依据。
参考文献
[1]米红波.活鱼运输箱内水质的控制及降低运输胁迫的方法[A].中国食品科学技术学会.中国食品科学技术学会第十一届年会论文摘要集[C].中国食品科学技术学会:中国食品科学技术学会,2014:2.
[2]管维良,刘天天,梁中永,等.活鱼运输的研究进展[J].轻工科技,2016,32(06):3-5.
[3]程守坤.鳜鱼活体运输技术基础研究[D].上海:上海海洋大学,2015.
[4]ROBERTSON OH,HANE S,WEXLER BC,et al. THE EFFECT OF HYDROCORTISONE ON IMMATURE RAINBOW TROUT (SALMO GAIRDNERII)[J]. General and Comparative Endocrinology,1963,3(4):422-436.
[5]陈仙标.模拟运输期间影响遮目鱼卵成活率和孵化率的主要因素[J].渔业信息与战略,1989(11):24-27.
[6]王文博,李爱华,汪建国,等.拥挤胁迫对草鱼非特异性免疫功能的影响[J].水产学报,2004,28(2):139-144.
[7]张勇.空气暴露及运输胁迫对美洲鲥亲鱼生理生化指标及hsp70基因表达的影响[D].南京:南京农业大学,2016.
[8]孙鹏,柴学军,尹飞,等.运输胁迫下日本黄姑鱼肝脏抗氧化系统的响应[J].海洋渔业,2014,36(5):469.
[9]Ruyet P L. Turbot (Scophthalmus maximus) grow out in Europe: practices,results and prospects[J]. Turkish Journal of Fisheries and Aquatic Sciences,2002,2:29-39.
[10]雷霁霖.鲆鲽类养殖新形势和发展动向[J].科学养鱼,2005(01):38-39.
[11]雷霁霖,门强.大菱鲆人工繁殖与养殖技术讲座(Ⅱ)[J].齐鲁渔业,2002(10):47-48.
[12]陈全震,曾江寧,高爱根,等.鱼类热忍耐温度研究进展[J].水产学报,2004,28(5):562-567.
[13]徐冬冬,楼宝,詹炜,等. 高温胁迫对褐牙鲆生长及肝脏抗氧化酶活性的影响[J].水产学报,2010,34(7):1099-1105.
[14]王美垚.急性低温胁迫及恢复对吉富罗非鱼血清生化、免疫以及应激蛋白HSP70基因表达的影响[D].南京:南京农业大学,2009.
[15]Cheng AC,Cheng SA,Chen YY,et al. Effects of temperature change on the innate cellular and humoral immune responses of orange-spotted grouper Epinephelus coioides and its susceptibility to Vibrio alginolyticus[J]. Fish and Shellfish Immunology,2009,26(5):768-772.
[16]Pickering A D,Pottinger T G. Stress responses and disease resistance in salmonid fish: Effects of chronic elevation of plasma cortisol.[J]. Fish Physiology & Biochemistry,1989,7(1-6):253-258.
[17]Lynne S. Procarione,Terence P. Barry,Jeffrey A. Malison. Effects of High Rearing Densities and Loading Rates on the Growth and Stress Responses of Juvenile Rainbow Trout[J]. North American Journal of Aquaculture,1999,61(2):91-96.
[18]陈欣然,牛翠娟,蒲丽君. 水产动物密度胁迫研究进展[J].中国水产科学,2007,14(s1):138-146.
[19]强俊,杨弘,何杰,等.3种品系尼罗罗非鱼生长及高密度胁迫后生理响应变化的比较[J].中国水产科学,2014,21(01):142-152.
[20]Thomas P M ,Pankhurst N W ,Bremner H A . The effect of stress and exercise on post﹎ortem biochemistry of Atlantic salmon and rainbow trout[J]. Journal of Fish Biology,2005,54(6):1177-1196.
[21]Cataldi E ,Marco P D ,Mandich A ,et al. Serum parameters of Adriatic Sturgeon Acipenser naccarii (Pisces: Acipenseriformes): effects of temperature and stress[J]. Comparative Biochemistry and Physiology - Part A Molecular & Integrative Physiology,1998,121(4):351-354.
[22]Cláudia Maria Domingues Alvarenga,Volpato G L . Agonistic profile and metabolism in alevins of the Nile tilapia[J]. Physiology & Behavior,1995,57(1):75-80.
[23]Cláudia Maria Domingues Alvarenga,Volpato G L . Agonistic profile and metabolism in alevins of the Nile tilapia[J]. Physiology & Behavior,1995,57(1):75-80.
[24]Ishioka H.Live fish transportation,series of fisheries book[M].Tokyo:Kosesha Kosekaku,1982.
[25]Del Maestro R F. An approach to free radicals in medicine and biology.[J]. Acta Physiologica Scndinavica Supplementum,1980,492(492):153-68.
[26]荣志江.过氧化氢酶的电化学研究[D].太原:山西大学,2006.
[27]Chen Q X,Zhang W,Zheng W Z,et al. Kinetics of inhibition of alkaline phosphatase from green crab (Scylla serrata )by N-bromosuccinimide[J]. Journal of Protein Chemistry,1996,15(4):345-350.
[28]彭小玉,沈明浩.蒲公英水提物对鲫鱼生长、鱼体成分、消化酶和免疫系统的影响[J].中国畜牧兽医,2010,37(5):13-17.
[29]孟晓林,冷向军,李小勤,等.杜仲对草鱼鱼种生长和血清非特异性免疫指标的影响[J].上海海洋大学学报,2007,16(4):329-333.
[30]Mingchun Ren,Bo Liu,Habte-Michael Habte-Tsion,Xianping Ge,Jun Xie,Qunlan Zhou,Hualiang Liang,Zhenxin Zhao,Liangkun Pan. Dietary phenylalanine requirement and tyrosine replacement value for phenylalanine of juvenile blunt snout bream,Megalobrama amblycephala[J]. Aquaculture,2015,442.
[31]Qi-hui Yang,Bei-ping Tan,Xiao-hui Dong,Shu-yan Chi,Hong-yu Liu. Effects of different levels of Yucca schidigera extract on the growth and nonspecific immunity of Pacific white shrimp(Litopenaeus vannamei) and on culture water quality[J]. Aquaculture,2015,439.
[32]王香麗,麦康森,徐玮,等.蛋氨酸对瓦氏黄颡鱼幼鱼肝脏及血浆中谷草转氨酶和谷丙转氨酶活力的影响[J].中国海洋大学学报:自然科学版,2015,46(9):49-53.
[33]N. ?ner,E. ?zcan Oru?,M. Canli,et al. Effects of Cypermethrin on Antioxidant Enzyme Activities and Lipid Peroxidation in Liver and Kidney of the Freshwater Fish,Oreochromis niloticus,and Cyprinus carpio,(L.)[J]. Bulletin of Environmental Contamination & Toxicology,2001,67(5):657-664.
[34]郑永华,蒲富永.汞对鲤鲫鱼组织转氨酶活性的影响[J].西南农业大学学报,1997(01):43-47.
[35]Callewaert L,Michiels CW. Lysozymes in the animal kingdom.[J]. Journal of Biosciences,2010,35(1):127-160.
[36]顾军,龚锦涵,殷岳保,等.高压氧间断暴露及氧惊厥对大鼠抗氧化酶活力及脂质过氧化物含量的影响[J].中华航海医学杂志,1995(04):31-34.
[37]狄瑜,徐奕,李姗敏,等.捕捉胁迫对暗纹东方鲀肝脏非特异性免疫指标的影响[J].浙江海洋学院学报(自然科学版),2015,34(4):314-317.
[38]苏鹏,潘金露,韩雨哲,等.壳寡糖对红鳍东方鲀血液指标和非特异性免疫指标的影响[J].大连海洋大学学报,2016,31(1):37-43.
[39]刘鹏,宋协法,葛长字.温度、体重和昼夜节律对大菱鲆幼鱼代谢的影响[J].河北渔业,2009(05):1-4+27.
[40]朱松泉.中国条鳅志[M].南京:江苏科学技术出版社,1989.
[41]林肇信,刘天齐,刘逸农.环境保护概论修订版[M].北京:高等教育出版社,1983.
[42]成庆泰,郑葆珊.中国鱼类系统检索(上,下)[M].北京:科学出版社,1987.
(责编:王慧晴)
