麦角硫因机体内的分布与代谢和其在疾病中的作用研究进展
2020-01-08潘虹余郭丽琼林俊芳
潘虹余,郭丽琼,2,林俊芳,2,
(1.华南农业大学食品学院,广东 广州 510642;2.广东省微生态制剂工程技术研究中心,广东 广州 510640)
1909年,有学者在研究麦角真菌Claviceps purpurea时分离出一种独特的白色结晶含硫化合物,后来被确定为2-硫基-L-组氨酸三甲基内盐[1],随后其被命名为麦角硫因(ergothioneine,EGT)。EGT可由非酵母真菌、放线菌目和某些蓝细菌合成[1]。动物机体自身不能合成EGT,只能从食物中摄取并积累在组织和细胞中。EGT具有很强的抗氧化性,在机体内扮演着重要的角色。随着人们对抗氧化剂研究的不断深入,EGT作为一种天然的生物活性物质受到人们广泛的关注,国外已有相关报道,而国内对其研究还处于探索阶段[2]。目前为止,鲜有研究探讨EGT在人类受试者中的药代动力学[2]。
1 EGT的结构、性质和生物合成
EGT是一种无味、无色化合物,分子式为C9H16N3O2S,25 ℃时水溶解度极限值为0.9 mol/L[3],分子结构中含有咪唑-2-硫酮基团。EGT在溶解状态下存在硫醇和硫酮两种结构的互变异构体(图1)。在生理pH值条件下EGT主要以硫酮形式存在于水溶液中,因此与其他天然存在的硫醇(例如谷胱甘肽、N-乙酰半胱氨酸)相比,EGT是非常稳定的抗氧化剂,在生理pH值下不易发生自氧化。在pH值为7时,EGT的标准氧化还原电势为-60 mV,其他天然存在的硫醇氧化还原电势在-200~-320 mV之间[4]。硫酮硫醇互变异构体和高氧化还原电位是EGT具有更高稳定性的原因,这种特性可防止EGT在生理pH值下像其他硫醇(如谷胱甘肽(glutathione,GSH))一样进行自氧化[4]。
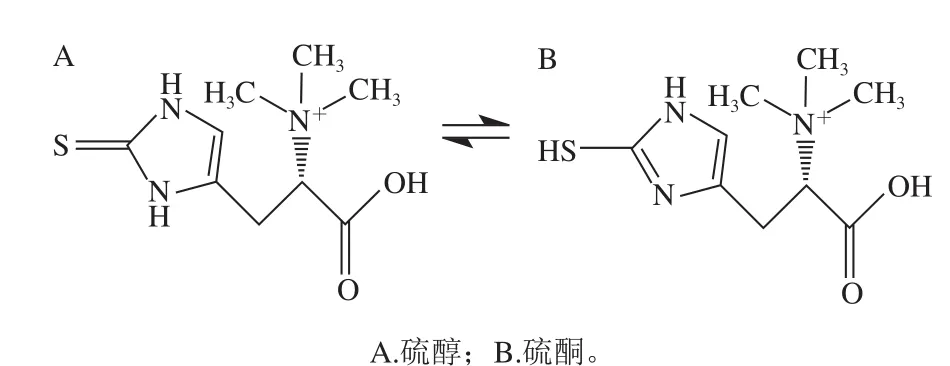
图1 EGT的硫醇和硫酮的互变异构体Fig. 1 Structures of thione-thiol tautomers of EGT
Seebeck首次探讨了合成EGT的基因簇(egt A/B/C/D/E)在分枝杆菌属中的应用[10],并且在许多真核生物和细菌中发现了EGT合成的关键基因,表明能够产生EGT的微生物不仅限于上文所述[10-11]。Seebeck[10]推测这5 个基因簇(egt A/B/C/D/E)主要负责分枝杆菌中EGT的生物合成(图2)。EGT的生物合成从L-组氨酸开始,通过中间体前体组氨酸三甲基内盐,分别从半胱氨酸和蛋氨酸获取硫和甲基以形成最终产物EGT[12]。EGT生物合成中存在两个主要反应,即氧化C—S键的形成和C—S键的裂解反应,使硫原子从半胱氨酸或c-谷氨酰半胱氨酸完全转移至组氨酸的咪唑侧链[13]。2018年,Osawa等[14]在大肠杆菌中成功表达了耻垢分枝杆菌中的EGT合成酶基因egt A、egt B、egt C、egt D、egt E,使重组菌的EGT产量达到了24 mg/L。
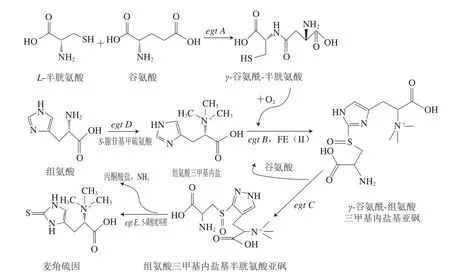
图2 EGT在分枝杆菌中的生物合成路径[10]Fig. 2 Biosynthetic pathway of EGT in mycobacteria[10]
2 EGT在体内的摄取、分布和代谢
2.1 EGT的摄取和分布
尽管EGT存在于许多细胞中,但其本身不能跨越细胞质膜,需要特定的载体将其运输至细胞内[15]。2005年,Gründemann等[16]在动物组织中发现了一种Na+依赖型EGT转运蛋白——有机阳离子转运蛋白1型(organic cation transporter novel type-1,OCTN1),并使用液相色谱-质谱差异着色法确定了EGT是OCTN1的关键底物。OCTN1的特异性表现在除了EGT外对其他有机阳离子摄取和转运率均很低,包括与EGT结构相似的化合物,例如甲巯咪唑和组氨酸三甲基内盐[17-18]。OCTN1对EGT的转运米氏常数(Km)很低(约为21 mmol/L),说明EGT是其特殊的结合底物[17-18]。
特定转运蛋白的存在表明动物(包括人类)可以从饮食中摄取EGT并通过在组织中积累而获益。EGT有自己的选择性转运蛋白用于肠道的摄取和输送到各组织[19]。早期研究揭示了EGT在组织中的积累具有性别差异[20](可能与睾酮水平有关)和年龄差异,EGT在体内组织中的水平具有与年龄增长相关的下降趋势[21-22]。尽管EGT在人体组织中分布较广泛,但它目前不被认为是必需的营养成分,因为没有研究发现其缺乏会引起不良反应。最近研究发现EGT的基线水平在肝脏和血细胞中最高,比脑和眼睛中高约10~20 倍,表明这些组织可能是体内EGT积累的主要部位[23]。对体内EGT分布的研究表明,EGT优先积聚在易受高水平氧化应激和炎症影响的器官、细胞和分泌物中,如肝脏、肾脏、红细胞[2,23]、眼晶状体和精液[15],这可能是一种适应性的细胞保护机制[17,24]。
2.2 EGT的代谢
EGT的代谢转换率相对缓慢,在禁食1 周的大鼠中观察到其在血液和肝脏中的水平没有明显下降[25],EGT的这种高积累和缓慢代谢的特征归因于肾脏重吸收和EGT的低尿排泄[2,26]。人体研究显示,尿液中EGT的排泄量很小,表明EGT在口服后主要在体内保留[2]。
Servillo等[27]研究了EGT的可能氧化产物,并提出组氨酸三甲基内盐和EGT-磺酸是EGT的稳定氧化产物。Cheah等[2]测定了组织中3 种与EGT相关代谢物(组氨酸三甲基内盐、EGT-磺酸、S-甲基-EGT)的分布水平,发现仅能检测到组氨酸三甲基内盐,并且其含量与各组织中的EGT浓度显著相关[23]。这可能表明组氨酸三甲基内盐确实是体内EGT的代谢产物。研究发现EGT的积累并不总是与上述氧化产物的水平相关,例如肝脏中EGT含量最高,但检测不出EGT-磺酸,而肺中EGT的含量处于中等水平,却检测出较高水平的EGT-磺酸[2]。
3 EGT的抗氧化特性
EGT的强抗氧化性可能与以下4 种分子活动有关:直接清除活性氧(reactive oxygen species,ROS)[28];有效螯合各种二价金属阳离子[29];激活抗氧化酶(如谷胱甘肽过氧化物酶和符胱什肽过氧化物酶),并抑制超级氧化激酶,如NADPH-细胞色素c还原酶;抑制各种血红素蛋白(如血红素和肌红蛋白)的氧化作用。
EGT与二价金属阳离子例如Cu2+、Hg2+、Co2+、Fe2+形成无氧化还原活性的络合物[30],其中最稳定的是铜,其具有最高的络合物形成常数[29]。这些金属离子在体内的结合可能有助于防止它们参与ROS的产生。
以前如果想买私募基金或者信托产品,只要满足100万元到300万元人民币不等的资金门槛就行,所谓的合格投资者审查基本上是一个形式。而资管新规实施后,无论是银行、券商还是信托、私募基金,发行的资管产品将被严格分为公募和私募两类。公募产品谁都能买,而私募产品仅供合格投资者。这里说的合格投资者不但必须具有2年以上投资经历,并且需要家庭金融资产至少在300万元到500万元以上,或者是投资者本人三年的年均收入在40万元以上。要求显然比以前提高不少。换句话说,目前的很多高净值客户以后只够格购买公募产品了。想买定制化、投资非标或者非上市股权的私募产品,需满足合格投资者的要求。
EGT具有调节炎症[31]、防止紫外线辐射诱导的损伤[32-33]以及其他保护细胞活动的功能[34]。与其他硫醇相比,EGT还显示出更高的速率(速率常数为2.3×107L/(mol·s)从而使单线态氧失活[35]。相比之下,EGT与超氧化物和H2O2不能很快地进行反应,与超氧化物反应的速率常数通常低于103L/(mol·s)[36]。EGT对由亚硝基铁氰化钠(硝普钠)产生的NO·诱导的细胞死亡没有影响,表明它与NO·的反应很缓慢或没有反应[37]。
研究表明,EGT可能作为适应性抗氧化剂在体内发挥抗氧化作用。EGT在健康动物中的抗氧化作用有限,但是在由组织损伤引起的自由基活性物质水平升高、存在ROS/活性氮(reactive nitrogen species,RNS)产生的毒素或涉及氧化应激引起的疾病程度加重时,EGT会发挥强有力的抗氧化作用[15-16]。通过检测血浆和尿液中的氧化损伤标准生物指标水平,发现EGT的摄入可减少由不同物质引起的氧化损伤,包括尿囊素(尿酸盐氧化)、8-羟基脱氧鸟苷(DNA损伤)、8-iso-PGF2a(脂质过氧化)、蛋白质羰基化和C-反应蛋白(蛋白质氧化),但减少程度不明显[2]。EGT的这种特性可能具有重要意义,即在不干扰ROS/RNS在健康组织中重要作用的情况下发挥抗氧化作用[38-40];当氧化损伤变得严重时,EGT则在体内组织中大量积累并发挥抗氧化作用[41]。
研究发现某些在体外具有强大抗氧化活性的物质在体内不发挥作用,例如许多类黄酮[42]。EGT在细菌、真菌[5-6,14,43]、秀丽隐杆线虫和动物体内也具有抗氧化作用。例如,缺乏EGT转运蛋白的秀丽隐杆线虫表现出较高的蛋白质羰基化水平(蛋白质氧化损伤的生物指标)[42,44];在缺乏EGT的斑马鱼中,8-羟基-2-多氧鸟苷和8-二氢鸟嘌呤(一种核酸氧化损伤的生物标志物)的水平增加[42,45]。
4 EGT的药代动力学
4.1 皮肤保护功能
EGT在皮肤细胞和离体皮肤的表皮细胞中呈现时间和浓度依赖性积累[46]。与具有较高氧化自由基吸收能力但生物利用度低和易被快速清除的抗氧化剂(如白藜芦醇)相比,EGT具有较好的生物利用度[47]。EGT不仅可以防止氧化损伤[32],还可对经紫外线照射的细胞进行DNA修复,防止单链DNA断裂[48],抑制细胞凋亡的发生,并使细胞活力增加[46,48]。细胞吸收、积累和利用L-EGT的能力使EGT及其受体/转运蛋白OCTN1成为皮肤抗氧化防御系统的重要组成部分[46]。
4.2 抗抑郁症
最近的研究发现,口服摄入的EGT可通过血脑屏障转运到大脑中,并在小鼠体内发挥类似抗抑郁药的作用。对小鼠进行强迫游泳实验(forced swimming test,FST)和悬尾实验(tail suspension test,TST),发现与正常饮食对照组小鼠相比,EGT显著缩短了处理组小鼠在FST和TST中的不动时间[49]。口服摄入EGT后,脑内EGT浓度呈剂量依赖性增加[49]。EGT有两种作用:抑制神经干细胞的增殖和促进神经元分化[49-50]。抗抑郁药物如氟西汀、丙咪嗪和瑞波西汀可促进神经新生,表明神经新生和抗抑郁作用之间可能存在关联[49,51]。EGT可能是通过促进神经元的分化来发挥其在体内的抗抑郁作用,但这一点需要进一步的研究来证实[49]。
4.3 眼部疾病
研究表明,EGT可以保护角膜内皮细胞免受氧化应激和未折叠蛋白反应[52]。最近发现EGT在人类泪液和房水样本中具有较高的积累水平(分别为约0.35 μmol/L和约28 μmol/L),并且证明EGT很容易在小鼠眼内积累[23]。早期研究发现在患有白内障的生物机体中,晶状体和角膜中的EGT水平显著降低;随着白内障程度的增加,EGT水平持续下降[53]。另一项研究发现用EGT治疗可以有效地保护小鸡免受糖皮质激素诱导的白内障损伤[54]。
4.4 心血管疾病
已知心肌缺血-再灌注损伤(ischemia-reperfusion injury,IR)可产生ROS/RNS,而ROS/RNS过度形成会加剧IR,导致氧化肌细胞损伤[55-56]。EGT在IR期间可保护心脏和肝脏组织免受损伤[57-58]。EGT保护心脏的机制之一可能是防止ROS/RNS氧化肌红蛋白,形成具有细胞毒性的铁蛋白肌红蛋白;EGT还能降低组织丙二醛水平并调节热休克蛋白70和促炎细胞因子(肿瘤坏死因子-α(tumor necrosis factor-α,TNF-α)和白细胞介素-1β(interleukin-1β,IL-1β))[58]。此外,EGT作为铁和铜的螯合物可通过减少IR期间自由基的产生来保护机体组织,这可能是EGT的另一种保护机制[59]。在小鼠体内,EGT容易在血液中累积并进入心脏,因此它可以发挥保护心脏的作用[2,23]。
4.5 肾脏疾病与糖尿病
最近一项研究发现,敲除编码OCTN1基因的慢性肾病(chronic kidney disease,CKD)小鼠模型出现肾纤维化恶化的症状,同时CKD患者的血液中EGT水平较低[60]。CKD会增加氧化损伤[61],EGT可能通过在肾脏中的积累来减少这种损害。
糖尿病和糖尿病并发症与氧化和糖化应激增加密切相关,部分原因是高血糖的促氧化作用[62]。EGT能够保护内皮细胞和大鼠嗜铬细胞瘤(PC12)细胞免受高葡萄糖水平引起的损伤[63-64];对患糖尿病的妊娠大鼠补充EGT后减少了畸形胚胎的发生率[65],这归因于EGT对高血糖依赖性氧化应激的调节[65]。
4.6 慢性炎症疾病
EGT在炎症中的作用颇有争议。研究发现,轻度类风湿性关节炎(rheumatoid arthritis,RA)患者体内的红细胞和单核细胞EGT水平显著升高,并且其与CD14+细胞中OCTN1 mRNA的表达密切相关[66]。在由胶原诱导的关节炎小鼠发炎关节的血液和免疫组织中也发现了高度表达的OCTN1[67]。与RA类似,对克罗恩病(Crohn's disease,CD)患者发炎回肠黏膜的分析中发现EGT水平显著升高;相应地,与非CD受试者相比,CD患者的OCTN1 mRNA表达也有增加[68]。
在培养的红细胞中,对OCTN1基因的沉默抑制了EGT摄取,并促进了细胞凋亡的发生[69]。在这些慢性炎症状态中,可能是由于EGT的抗细胞凋亡特性可介导免疫细胞的存活并刺激炎症的发生,进而导致EGT在体内组织中水平的增加[68]。一些研究表明,EGT水平升高的另一个原因可能是由于细胞因子介导的OCTN1表达上调。用TNF-α孵育Caco-2细胞,结果显示通过TNF-α受体-1的激活增加了OCTN1表达,因此EGT的增加可能是通过上述提到的延长免疫细胞存活来进一步促进炎症反应[68]。类似的,与RA相关的炎性细胞因子例如IL-1β和转录因子RUNX1能够上调OCTN1的表达。然而,这种反应是否与发病机制相关或作为细胞防御的反作用反应,以及EGT的分子作用仍未确定。
4.7 神经退行性疾病
EGT在大脑中的功能尚不确定,但在脑和各种神经细胞中能够观察到OCTN1表达[70]。EGT渗透血脑屏障的能力表明EGT能在中枢神经系统和神经元中发挥功能性作用。EGT可通过激活p38 MAPK细胞内抗氧化途径以保护PC12细胞免受H2O2诱导的细胞损伤[71],还可抑制由β-淀粉样蛋白和顺铂诱导的神经元细胞凋亡[55]。研究表明,轻度认知受损受试者和帕金森病患者的血液中EGT水平显著低于同年龄的健康个体[72-73]。这可通过多种机制引起,例如饮食的改变和/或OCTN1活性的变化。最近有学者提出蘑菇摄入量的增加与痴呆发病率降低之间存在相关性[74-75]。然而蘑菇含有多种可能具有生物活性的化合物,不能认为是EGT发生作用。因此,仍然需要更多研究来讨论EGT是否对神经退行性疾病产生有益作用。
5 EGT的安全性
迄今为止对EGT的毒理学研究未发现其有副作用[76]。然而,一些人认为在慢性炎症中,EGT的积累会加重炎症反应[68]。为证明发炎和受损组织是否会自发增加EGT水平使其自身受到进一步伤害,推测细胞可通过EGT的积累来进行自我保护,有助于保护组织免受氧化损伤的进一步伤害。因此,可以推断在一些人类疾病中观察到的EGT在组织中的积累不是病理性的,而是细胞保护性的[77-80]。例如一项代谢组学研究确定了压力超负荷和梗塞的小鼠心脏中EGT水平显著增加[81],这可能反映了机体组织能通过积累EGT来保护自身免受进一步的伤害。
2017年,欧洲食品安全局发布报告提出EGT作为膳食补充剂和强化食品摄入不会加剧RA、CD和糖尿病的易感性和病症发展;同时,其下属的营养产品、营养及致敏科学组提出EGT的婴儿安全每日限量为2.82 mg/kg mb,儿童为3.39 mg/kg mb,成人(包括孕妇和哺乳期妇女)为1.31 mg/kg mb[82]。因此新型合成L-EGT在安全食用水平下对成人(孕妇)、儿童和婴儿均是安全的。
6 结 语
虽然近年来对EGT的研究不断取得进展,并有大量研究证明EGT在体外具有多种抗氧化和细胞保护作用;但是EGT的真正生理作用尚未完全阐明,这些细胞保护作用背后的分子机制仍然未知。同时,由于体外研究中采用的条件与体内不同,体外研究结果不能完全应用于体内。因此,仍需要大量的体内研究来验证EGT的治疗潜力。
此外,不仅某些真菌,一些致病微生物、病原体也会产生EGT并利用它来保护自身免受宿主防御(包括ROS/RNS)的伤害[43,67],例如结核分枝杆菌(人结核病的致病因子),而其EGT合成能力的丧失也会降低它们的毒力。因此,EGT生物合成途径的抑制剂(应该是一种可行的治疗方法,因为这种途径不会在人体中发生)可能是攻击致病性合成EGT生物的新型药物潜在来源。
由于EGT在生物体内的作用机制及与疾病的关系还不清楚,限制了其生产和应用。因此需要对EGT作为免疫调节剂和某些疾病的治疗剂开展更多的研究,探讨其在体内的作用机制,以推进EGT在疾病治疗上的应用。
