5种绿化植物对SO2的吸收能力与生理响应
2019-11-28李其营
摘要:采用人工模拟熏气法,研究玉兰、紫荆、蔷薇、香樟、夹竹桃5种绿化植物对SO2气体吸收净化能力及其生理变化特征。结果显示,在不同SO2质量浓度(0.25、0.50 mg/m3)环境下,玉兰和香樟叶片硫含量以及对SO2吸收净化量最高,叶片硫含量及SO2吸收净化量均表现为玉兰和香樟显著高于紫荆、蔷薇和夹竹桃(P<0.05),紫荆和夹竹桃较低,不同绿化植物叶片硫含量以及对SO2吸收净化量随SO2浓度的增加而增加。玉兰和香樟叶片类胡萝卜素、叶绿素、可溶性蛋白和可溶性糖含量最高,叶片类胡萝卜素含量、叶绿素含量、细胞膜渗透率、可溶性蛋白含量和可溶性糖含量均表现为玉兰>香樟>蔷薇>紫荆>夹竹桃。紫荆和夹竹桃叶片游离脯氨酸含量、细胞膜渗透率、丙二醛含量最高,叶片游离脯氨酸、丙二醛含量均表现为玉兰<香樟<蔷薇<紫荆<夹竹桃。不同绿化植物叶片净化量与叶片生理特性具有显著的相关性,叶片对SO2吸收净化量与类胡萝卜素含量、叶绿素含量、细胞膜渗透率、可溶性蛋白含量和可溶性糖含量呈显著或极显著的正相关,与游离脯氨酸、丙二醛含量呈显著或极显著的负相关,以玉兰和香樟的相关系数绝对值最大。由此可知,玉兰和香樟对SO2吸收净化能力较强。其研究结果为我国城市功能型植物选择和生态景观林带建设提供科学依据。
关键词:绿化植物;SO2;吸收能力;生理特性;响应
中图分类号: X173文献标志码: A
文章编号:1002-1302(2019)18-0182-05
收稿日期:2018-05-29
作者简介:李其营(1981—),男,河南南阳人,硕士,讲师,研究方向为园林植物生态学。E-mail:Qiying_lee81@163.com。
在现代城市中,大气中SO2是主要的污染物之一,不仅会污染到植物,而且还会影响到人们的健康。SO2污染在超标的情况下会形成酸雨,酸雨的来源主要是日常生活中燃烧的煤与石油以及工业硫矿石的燃烧[1-3]。建设生态文明城市,绿化部门需要多种植绿色植物,因为植物对空气有净化作用,可以将大气层中的粉尘、颗粒物进行过滤,具有吸附功能,能够改善被污染的环境,增大空气中的湿度、减少阳光辐射、调节气候等[4]。在建立生态文明城市的过程中,广大群体应该意识到种植植物与环境之间具有促进关系,但是从当今社会发展趋势来看,很多农村都盖集资楼,占用农民的田地,绿色面积不断地减少,空气污染变得更为严重[5-6]。通过以上阐述的绿色植物作用来看,除了具有经济产出的功能外,还具有绿化、美化、净化等生态效益。绿色植物的作用与自然界中的植物一样,都具有较强的光合作用,以吸收CO2,释放O2,吸附环境中的有毒物质,对城市的绿化和空气的净化具有重要作用。
从当前国内发展的趋势来看,冬季雾霾天气增多,广大群体逐渐对大气污染进行关注,由于大型、中型、小型城市交通网,信息网与社会经济联系较为密切,多种污染结合,突出了大气污染环境的问题,并且生物的有机生命已经受到了污染源放出的大气污染物及其生产物的影响,全球将近1/2以上的人都生存在这样的环境下,广大群体的身体健康受损害,这也成为阻碍社会经济迅速发展的因素[7]。
通过搜集关于城市绿化植物净化空气方面的文献资料,了解到国内外关于城市绿化植物净化大气层中的SO2主要是分析绿化植物吸收大气中SO2的效果,采用野外污染与清洁干净的绿化植物叶片中的硫进行测验,但采用人工模拟熏气试验开展的绿色植物吸收SO2[8-9]的研究还较少。江西省作为国内的大型省份之一,社会经济发展较快,各种车型都不断增加,城市中污染最为突出的问题就是大气方面的污染,排放的SO2不断增加,空气变得非常浑浊,严重影响广大群众身体健康与建设绿色文明城市。对大气污染条件了解下,才能发现城市中植物生理特点的变化,这对分析城市大气污染的生物效应具有重要的意义,为研究大气污染对植物的伤害与植物的抗性机理提供相应的参考。因此,本研究针对人工熏气法对江西省南昌市绿色植物吸收SO2净化能力展开定量的分析,对城市绿化过程中选择树种提供相应的参考。
1 材料与方法
1.1 人工气候室
封闭式自动检测熏气设备,可以模拟自然界中温度、湿度、光照、SO2浓度,并能够对其进行恒温控制,称之为人工气候室[10]。在此系统中,可以设置和修订温度、湿度、光照、SO2等相关参数。对于SO2的储存主要是采用钢瓶,各自的管道经过阀门减压后才能运输到气候箱内,监测人员还可以在管路中安装精度SO2流量计与电子流量调节阀,顺着气体喷嘴流入到气候室[11-12]。通常情况下,气候室主要是采用红外线SO2传感器对室内的值进行测算,计算机上所显示出的数据与试验开始前SO2浓度进行对比,利用PID对电子流量节阀进行控制,使得室内气候SO2浓度更为准确。在人工气候室底侧栅板传送风,上侧栏板回风,这样才能使得人工气候空气进行循环,确保混合气体之间的均衡性,人工气候室的尺寸通常是2.0 m×1.5 m×1.5 m。
1.2 树种选择
本研究选取南昌市园林绿化植物玉兰、紫荆、蔷薇、香樟、夹竹桃作为试验对象(表1),选择1~2年盆栽实生苗,生产情况、大小高矮基本相同,每种树苗栽植5盆,将其放置到瓷盘中适当浇水,这样才能在人工气候室进行人工熏氣试验,不同的SO2浓度下所吸附的污染能力也是不同的。分别从东、西、南、北采集植物叶片,分别剪取上、中、下部分的茎,当所剪取完整的叶片放置到密闭比较好的锥形瓶子内,应用现代数字图像对洗干净的叶片测定面积(包含叶柄),比叶重=叶片干质量/叶面积。
1.3 研究方法
笔者在2016年11月24到2017年12月24日,利用人工气候室模拟大气环境条件,SO2的质量浓度设置为(0.25±0004)、(0.50±0.005) mg/m3,熏气30 d,对绿色植物吸附大气污染物中的SO2净化效果进行分析[13]。当白天温度维持在30 ℃时,夜间为20 ℃时,其湿度达到了65%~85%,白天的光照度应该控制在3 600 lx,夜间的约为300 lx,在相应的自然条件下应该对植物的生产范围进行控制。植物在生长过程中需要硫元素,但是这个不能作为净化大气硫的标准,此外,当植物进行大气净化时,植物还可以吸收到更多的硫[14-15]。本试验过程中需要注意的是可以在树木多点进行采样,用离子水清洗叶片、擦干,将此放入到烘箱内烘干,取出将其打磨,放到干净的广口试剂瓶内,储备多个干燥的试剂瓶备用[16-17]。植物叶片中硫质量分数通常采用硫酸钡比浊法。叶片硫吸附质量分数(mg/g)=熏气后硫的质量分数-熏气前硫的质量分数[12]。
1.4 叶片生理指标的测定
浓度不同的SO2气体熏气后,从中取出数量相等的叶片在蒸馏水中浸泡,使用电导仪器分为测算出煮沸前、后电导率,计算出相互对应的电导(%)。
将新鲜植物的叶片进行磨合后,以80%丙酮溶液浸泡分析测定叶绿素含量;茚三酮比色法测定游离脯氨酸含量;硫代巴比妥酸法测定丙二醛含量;考马斯亮蓝-G250染色法测定可溶性蛋白含量;蒽酮比色法测定可溶性糖含量;植物全硫含量的室内测定采用硫酸钡比浊法[12]。
1.5 数据处理与分析
利用Excel 2010将相关数据进行整理,方差与统计学的检验采用SPSS 18.0,LSD多重比较(显著水平设置α=0.05、0.01),单因素方差分析(One-way ANOVA)比较其差异显著性。
2 结果与分析
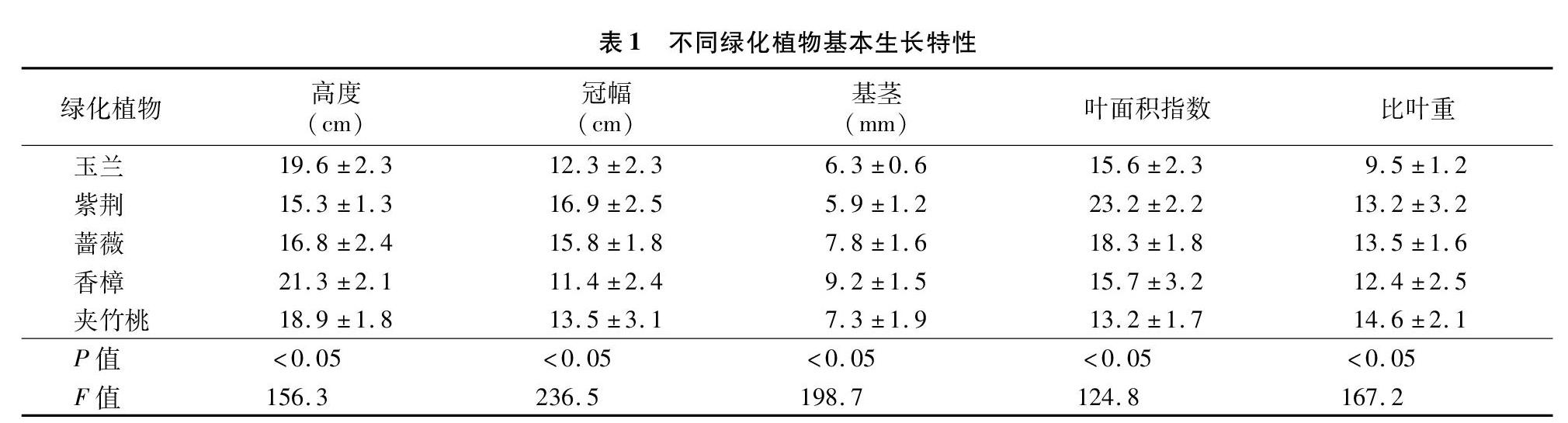
2.1 不同SO2浓度下植物叶片硫含量
由图1可知,在SO2质量浓度为0.25 mg/m3环境下,玉兰和香樟叶片硫含量较高,分别为4.32、4.01 mg/g,显著高于紫荆、蔷薇和夹竹桃(P<0.05),其中玉兰和香樟、紫荆和夹竹桃差异不显著,叶片硫含量大小依次为玉兰>香樟>蔷薇>夹竹桃>紫荆。在SO2质量浓度为0.50 mg/m3环境下,玉兰和香樟叶片硫含量较高,分别为5.36、5.13 mg/g,显著高于紫荆、蔷薇和夹竹桃(P<0.05),其中玉兰和香樟、紫荆和夹竹桃差异不显著,叶片硫含量大小依次为玉兰>香樟>蔷薇>紫荆>夹竹桃。
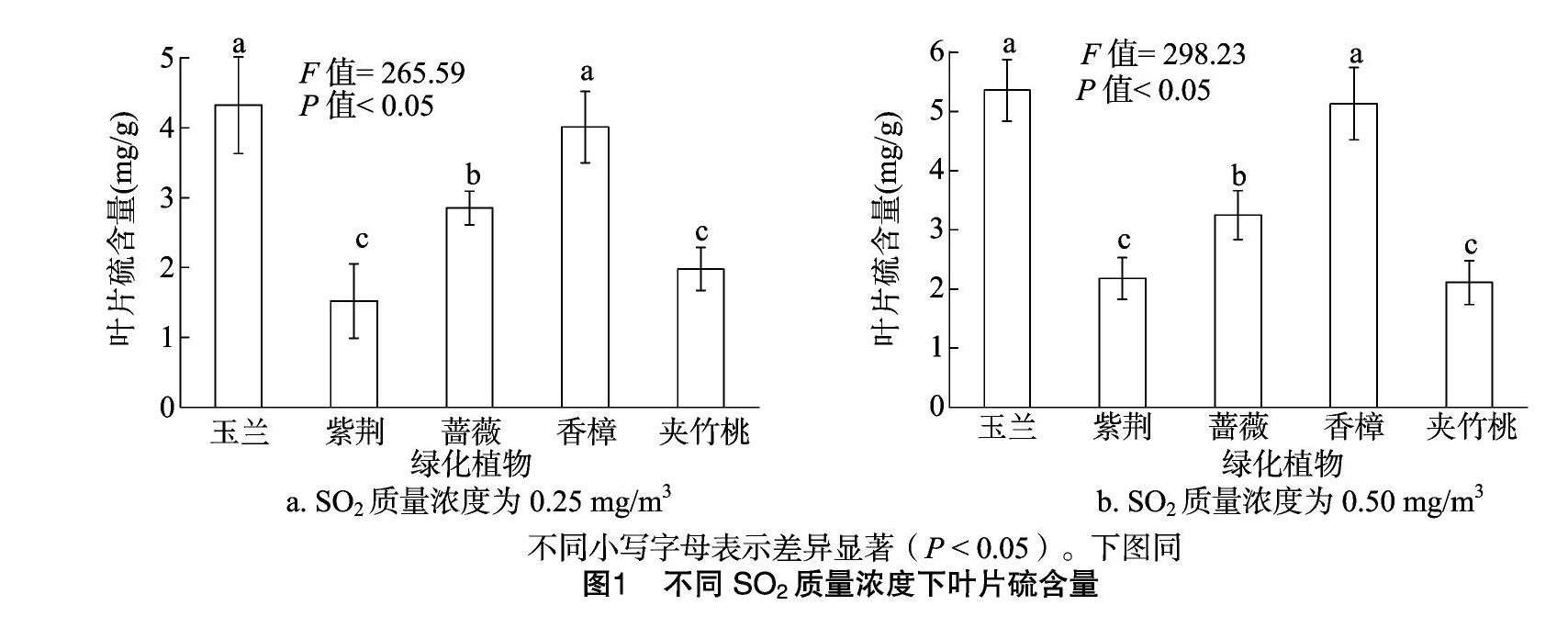
2.2 不同SO2质量浓度下植物吸收净化能力
由图2可知, 在SO2质量浓度为0.25 mg/m3环境下,玉兰和香樟叶片硫吸收量较高,分别为3.25、3.12 mg/g,显著高于紫荆、蔷薇和夹竹桃(P<0.05),其中玉兰和香樟、紫荆和夹竹桃叶片硫含量差异不显著,叶片硫吸收量大小依次为玉兰>香樟>蔷薇>紫荆>夹竹桃。在SO2质量浓度为050 mg/m3环境下,玉兰和香樟叶片硫吸收量较高,分别为4.01、3.84 mg/g,显著高于紫荆、蔷薇和夹竹桃(P<0.05),其中玉兰和香樟、紫荆和夹竹桃差异不显著,叶片硫含量大小依次为玉兰>香樟>蔷薇>紫荆>夹竹桃。
2.3 不同SO2质量浓度下植物叶片光合色素含量
光合色素在植物光合作用的原初光反应过程中起着关键作用,其含量的变化往往与叶片的生理活性、植物对环境的适应性和抗逆性有关。本研究中不同绿化植物叶片叶绿素含量和类胡萝卜素含量也显示出一定的差异(表2)。在SO2质量浓度为0.25 mg/m3环境下,玉兰和香樟叶片类胡萝卜素含量较高,分别为9.23、8.26 g/cm2,显著高于紫荆、蔷薇和夹竹桃(P<0.05),其中玉兰和香樟、紫荆和夹竹桃差异不显著,叶片类胡萝卜素含量大小依次为玉兰>香樟> 蔷薇>紫荆>夹竹桃。在SO2质量浓度为0.50 mg/m3环境下,玉兰和香樟叶片类胡萝卜素含量较高,分别为8.16、7.16 g/cm2,显著高于紫荆、蔷薇和夹竹桃(P<0.05),其中玉兰和香樟、紫荆和夹竹桃差异不显著,叶片类胡萝卜素含量大小依次为玉兰>香樟>蔷薇>紫荆>夹竹桃。在SO2质量浓度为0.25 mg/m3 环境下,叶片叶绿素a和b含量呈一致的变化规律,大小依次为玉兰>香樟>蔷薇>紫荆>夹竹桃;在SO2质量浓度为0.50 mg/m3环境下,叶片叶绿素a和b含量呈一致的变化规律,大小依次为玉兰>香樟>蔷薇>紫荆>夹竹桃。
2.4 不同SO2質量浓度下植物叶片细胞膜渗透率
细胞膜渗透率是反映膜系统稳定性的一个重要指标。由图3可知,不同SO2质量浓度下绿化植物膜系统出现明显的损伤。在SO2质量浓度为0.25 mg/m3环境下,紫荆和夹竹桃叶片细胞膜渗透率较高,分别为24.3%、22.1%,显著高于玉兰、蔷薇和香樟(P<0.05),其中紫荆和夹竹桃、玉兰和香樟差异不显著;在SO2质量浓度为0.50 mg/m3环境下,紫荆和夹竹桃叶片细胞膜渗透率较高,分别为35.6%、32.1%,显著高于玉兰、蔷薇和香樟(P<0.05),其中紫荆和夹竹桃、玉兰和香樟差异不显著。
2.5 不同SO2质量浓度下植物叶片生理指标
由表3可知,不同SO2质量浓度下绿化植物膜系统出现明显的损伤。在SO2质量浓度为0.25 mg/m3环境下,玉兰和香樟叶片可溶性蛋白含量较高,分别为123.26、115.89 μg/g,显著高于紫荆、蔷薇和夹竹桃(P<0.05),其中玉兰和香樟差异不显著,紫荆、蔷薇和夹竹桃差异不显著,叶片可溶性蛋白含量由大到小依次为玉兰>香樟>蔷薇>紫荆> 夹竹桃。可溶性糖含量由大到小依次为玉兰>香樟>蔷薇>紫荆>夹竹桃,不同植物可溶性糖含量差异均不显著。游离脯氨酸和丙二醛含量呈一致的变化趋势,均表现为夹竹桃>紫荆>蔷薇>香樟>玉兰,其中香樟和玉兰差异不显著。
在SO2质量浓度为0.50 mg/m3环境下,玉兰和香樟叶片可溶性蛋白含量较高,分别为116.15、103.27 μg/g,显著高于紫荆、蔷薇和夹竹桃(P<0.05),其中玉兰和香樟差异不显著,叶片可溶性蛋白含量由大到小依次为玉兰>香樟>蔷薇>紫荆>夹竹桃。可溶性糖含量由大到小依次为玉兰>香樟>蔷薇>紫荆>夹竹桃,不同植物可溶性糖含量差异不显著。游离脯氨酸和丙二醛含量呈一致的变化趋势, 均表现为夹竹桃>紫荆>蔷薇>香樟>玉兰,其中香樟和玉兰差异不显著。
2.6 綠化植物净化量与叶片生理特性相关性
由表4可知,不同绿化植物叶片净化量与叶片生理特性具有显著的相关性,玉兰叶片净化量与类胡萝卜素、叶绿素a和b含量、细胞膜渗透率、可溶性蛋白含量、可溶性糖含量呈极显著正相关(P<0.01),与游离脯氨酸、丙二醛含量呈极显著负相关(P<0.01)。紫荆叶片净化量与类胡萝卜素、叶绿素a含量呈显著正相关(P<0.05),与游离脯氨酸、丙二醛含量呈显著负相关(P<0.05)。蔷薇叶片净化量与类胡萝卜素含量呈极显著正相关(P<0.01),与叶绿素a和b、可溶性糖含量呈显著正相关(P<0.05),与游离脯氨酸、丙二醛含量呈显著或极显著负相关。香樟叶片净化量与类胡萝卜素含量、叶绿素a和b含量、细胞膜渗透率、可溶性糖含量呈极显著正相关(P<0.01),与可溶性蛋白含量呈显著正相关(P<0.05),与游离脯氨酸、丙二醛含量呈极显著负相关(P<0.01)。夹竹桃叶片净化量与叶绿素a含量呈显著正相关(P<0.05),与游离脯氨酸和丙二醛含量呈显著负相关(P<0.05)。
3 讨论与结论
大气层中存在有毒的物质,绿化植物对SO2具有一定的吸收能力[17-18],在低质量浓度的SO2中绿化植物可以表现出一种受害的情况,从而可以利用对SO2的感应度检测大气污染,起到了一种警示作用;此外,绿化植物还可以吸附其他大量的有害物质,起到净化环境的目的。在日常环境中SO2主要来自煤与石油燃烧,在现代城市中汽车燃料燃烧过程中也会产生SO2。按照植物含硫量的标准,部分研究人员已经将植物硫累积量作为大气SO2的指示剂[19-22],从而大气环境中SO2的污染程度通过绿化植物含硫量可测得。目前,国内关于交通污染对城市绿化植物影响研究比较少。通过相应的调查了解到,部分植物在经过熏气试验后选择出来抗性的品种再生产过程比较慢。如抗SO2很强的树种玉兰与香樟,当种植在交通比较繁华的地段时容易出现枯死的情况,导致出现这种情况的原因是在熏气试验中得出的植物抗性强弱的结论是根据气体受到污染状况下得出,并且这些植物是幼株,但是在交通环境下的植物都受多种污染物的复合污染,如乙烯、NOX、CO等[23-25]。除此以外,在对熏气试验进行时,其中的气温、湿度、风向都保持稳定的状态,但是交通环境中的气温、湿度、风力都随时发生变化,从而仅仅是选择那些抗污性绿色植物是远远不够的。本研究从生态检测着手,在对生态有所了解的基础上选择栽种面积广、数量大的绿色植物作为材料,通过污染物整体的含量对城市中绿色植物进行分析,这样才能继续对植物进行筛选,从中选择抗污性较强的植物为其他区域提供依据。
本研究中通过人工熏气箱实际情况了解到,植物受到SO2的伤害程度与品种有着直接的联系。经过相应的试验研究发现,在植物的叶片中SO2可以通过气孔进入到叶肉组织,发生相应的变化产生一些新的物质,间接或直接地对绿色植物产生伤害。SO2在植物的细胞中可以释放出H+、HSO3-和SO32-,SO32-在光下叶绿体产生的O2启动下,可氧化成SO42-,同时产生大量的活性氧,从而对细胞产生直接或间接的毒害作用,造成植物叶片受害,其生理特性功能降低。绿色植物对于SO2吸收量被称为相对吸收量,这充分证明植物对大气中SO2的净化作用,通常吸附属于物理性的过程,与植物表面的结构如叶片形态、粗糙程度、叶片角度与分泌物都有直接的关系[26-27]。植物对污染物的吸附通常是发生在地上部分的表面及其叶片的气孔,植物的吸附净化作用与叶片结构也存在相应的关系,叶片有蜡质、革质或叶面密生绒毛的植物污染气体不能畅通地进入叶内,对植物吸收净化效果产生较大的影响。本研究表明,在不同质量浓度SO2处理下,香樟与玉兰对SO2吸收净化能力处于弱势,产生这些情况是由于香樟与玉兰的叶片为革质,叶内SO2气体不能畅通地进入,这对叶片吸收净化效果的影响较大。
叶绿素是植物光合作用的基础物质,在光合条件下将其转化成能量;代谢的酶都包含在可溶性蛋白与可溶性糖中,产生的含量与植物自身的代谢能力有关。本研究表明,当植物的叶片受到大气污染后,叶片中的叶绿素含量也会受到影响,会不断地减少,不同的树种可溶性蛋白、可溶性糖、叶绿素含量也是不同的,如樟树与紫薇叶绿素含量偏高,白兰与木棉相对偏低,进一步的比较发现,玉兰与香樟都可以很好地光合作用,为光合补偿物质转化中需要的能量,其原因是SO2处理下植物叶片可溶性糖含量上升。当SO2质量浓度较低时,植物积累了很多的可溶性糖转化为其他物质,对污染进行抵抗,各种叶绿素受到影响,造成叶片中的叶绿素有所下降,但缺乏生物学与生理学方面的解释。当在逆境环境下绿色植物受到损伤后,通常会发生膜脂过氧化,膜脂过氧化作用中最为重要的产物之一就是丙二醛,其含量与质膜透性都有直接关系[28-30]。在正常条件,植物中活性氧的产生与清洗作用处于平衡状态,当处于逆境环境时,植物清除污染的能力也会受到影响,不利于体内活性氧产生,长期积累的活性氧会发生膜脂过氧化,植物中丙二醛积累得越多,植物受到的危害就越大,所处的环境越恶劣。通过该次不同绿化树种叶片丙二醛含量呈现相反趋势情况,充分证明了SO2处理下,植物体内的丙二醛会不断地增加,对植物造成了一定的损害。
参考文献:
[1]刘艳菊,丁 辉. 植物对大气污染的反应与城市绿化[J]. 植物学通报,2001,18(5):577-586,576.
[2]陆焕海,曹仲伟,孙羿灵,等. 大气污染经雨水对植物花粉活性影响的初步分析[J]. 生物学通报,2016,51(7):44-46,64.
[3]田育红,黄薇霖,刘怡青,等. 城市空气污染的植物监测研究进展[J]. 北京师范大学学报(自然科学版),2017,53(2):242-246.
[4]刘 波,赵建华,刘曦子,等. 4个树种叶片的重金属含量特征及其指示大气污染的研究[J]. 浙江林业科技,2016,36(5):1-7.
[5]杨晓晓,杨 丹,方欢欢,等. 3种地被植物对二氧化硫胁迫的生理响应[J]. 西北植物学报,2016,36(2):361-369.
[6]杨瑞卿,杨学民,申 晨. 21种园林植物对大气重金属污染物的吸收能力比较[J]. 江苏农业科学,2016,44(8):515-518.
[7]谢丽宏,黄芳芳,甘先华,等. 城市森林净化大气颗粒物污染作用研究进展[J]. 林业与环境科学,2017,33(3):96-103.
[8]陈伟光,黄芳芳,温小莹,等. 大气SO2和NO2污染及植物的抗性和净化能力研究进展[J]. 林业与环境科学,2017,33(4):123-129.
[9]王会京,谢宇光,王月惠. 河北省大气污染对不同绿化植物生理生态特性的影响[J]. 水土保持研究,2017,24(3):170-176,182.
[10]颉洪涛,顾沈华,刘丽月,等. 浙北52种景观树种对大气硫、氟污染物吸收富集能力研究[J]. 热带亚热带植物学报,2017,25(5):456-464.
[11]张鹏骞,朱明淏,刘艳菊,等. 北京路边9种植物叶片表面微结构及其滞尘潜力研究[J]. 生态环境学报,2017,26(12):2126-2133.
[12]毛东雷,蔡富艳,雷加强,等. 新疆策勒不同下垫面大气降尘时空分布特征[J]. 干旱区研究,2017,34(6):1222-1229.
[13]张家洋. 新乡市2种常见绿化植物叶片及叶面降尘的硫含量研究[J]. 西南林业大学学报,2016,36(4):167-170.
[14]張 璘,张祥志,秦 玮,等. G20峰会期间宜兴市大气VOCs特征及来源分析[J]. 环境科学,2017,38(7):2718-2727.
[15]毛永成,刘 璐,王小德. 干旱胁迫对3种槭树科植物生理特性的影响[J]. 浙江农林大学学报,2016,33(1):60-64.
[16]郭俊刚. 粉尘污染对不同园林植物生理特性的影响[J]. 江苏农业科学,2016,44(1):231-235.
[17]郑欣颖,张 潮,薛 立,等. 盐胁迫对4种园林植物生理特性的影响[J]. 河北农业大学学报,2017,40(6):44-50.
[18]庞发虎,杜瑞卿,周 索,等. 植物生理特征对南阳市空气中SO2浓度的监测分析[J]. 湖北农业科学,2016(5):1142-1146.
[19]何静雯,明 萌,卢 丹,等. 弱光胁迫对植物生理特性影响的研究进展[J]. 中国农学通报,2018,34(6):123-130.
[20]史冬冬,招礼军,朱栗琼,等. 自然低温对4种榕属植物生理特性的影响及抗寒性评价[J]. 广西林业科学,2016,45(2):170-174.
[21]胡羡聪,张德强,孔国辉,等. 大气SO2、氟化物对植物生理生态指标的影响[J]. 热带亚热带植物学报,2003,11(4):372-378.
[22]林双冀,孙 明. 盐胁迫下芙蓉菊与4种菊属植物生理响应特征及其耐盐机理分析[J]. 西北植物学报,2017,37(6):1137-1144.
[23]韩 枫,邓红章,李春荣,等. 土壤高浓度二氧化碳对植物生理生化特性的影响[J]. 应用化工,2017,46(3):422-425.
[24]李 洁,徐军桂,林 程,等. 引发对低温胁迫下不同类型玉米种子萌发及幼苗生理特性的影响[J]. 植物生理学报,2016,52(2):157-166.
[25]欧宇丹,邵 玲,周 澄,等. 不同叶色型苋菜叶片光合生理特性对弱光胁迫及恢复的响应[J]. 植物生理学报,2016,52(10):1527-1536.
[26]卞凤娥,孙永江,牛彦杰,等. 高温胁迫下根施褪黑素对葡萄叶片叶绿素荧光特性的影响[J]. 植物生理学报,2017,53(2):257-263.
[27]查 倩,奚晓军,蒋爱丽,等. 高温胁迫对葡萄幼树叶绿素荧光特性和抗氧化酶活性的影响[J]. 植物生理学报,2016,52(4):525-532.
[28]李美琦,姜在民,赵 涵,等. 加杨水力学与生理特性对不同土壤水分条件响应研究[J]. 植物生理学报,2017,53(4):632-640.
[29]殷庆霏,郭建斌,倪肖卫,等. 不同堆肥对南方屋顶绿化植物生长特性的影响[J]. 环境工程学报,2017,11(11):6205-6213.
[30]王雪莉. 兰州地区植物PAHs超累积特性和生物炭对土壤PAHs污染修复初探及对策[D]. 兰州:兰州大学,2016.
