生物质炭和微生物菌剂配施对设施土壤理化特性及黄瓜产量和品质的影响
2019-11-28张功臣赵征宇王波李磊秦玉红王珍青张守才
张功臣 赵征宇 王波 李磊 秦玉红 王珍青 张守才

摘要:通过温室盆栽试验研究单施小麦秸秆炭(T1)、单施微生物菌剂“宁盾”(T2)、小麦秸秆炭和微生物菌剂配施(T3)对连作土壤理化性状及黄瓜产量和品质的影响,以不进行任何处理的黄瓜连作土壤作对照(T0)。研究结果表明,生物质炭和微生物菌剂配施条件下,土壤速效氮含量、速效钾含量、全氮含量、全钾含量与对照相比显著增加,而各处理间速效磷含量、全磷含量差异不显著;在T2、T3处理条件下,土壤蔗糖酶活性显著高于对照,而土壤脲酶活性显著低于对照。与对照相比,T3处理条件下黄瓜根际土壤中可培养细菌数量显著增加,真菌数量显著降低。小麦秸秆炭和“宁盾”配施可显著提高黄瓜的产量和品质,春季、秋季两季黄瓜的总产量分别为114.41、85.80 t/hm2,比对照分别增加15.89%、12.36%;黄瓜果实的维生素C含量与对照相比增加21.22%,而硝酸盐含量与对照相比降低37.63%。因此,生物质炭和微生物菌剂配施可增加连作土壤养分含量,改善土壤酶活性和微生物菌群结构,提高黄瓜产量和品质。
关键词:连作土壤;黄瓜;微生物菌剂;协同作用;生物质炭
中图分类号: S642.206文献标志码: A
文章编号:1002-1302(2019)18-0155-05
收稿日期:2019-05-23
基金项目:现代农业产业技术体系建设专项资金(编号:CARS-23-G11);青岛市农业科学研究院博士基金(编号:2019-14-23)。
作者简介:张功臣(1986—),男,山东青岛人,博士,助理研究员,从事设施蔬菜栽培與环境调控技术研究。E-mail:gczhangnky@163.com。
通信作者:张守才,研究员,从事黄瓜遗传育种与栽培技术研究。E-mail:zsc403@163.com。
设施蔬菜生产高度集约化,栽培管理难度大,特别是连作和长期不平衡施肥导致的土壤酸化、次生盐渍化、土传病害加剧、微生物群落功能劣变等土壤障碍问题逐年加重,严重影响设施蔬菜产业可持续发展[1]。因此,如何改善土壤微生态环境,增加土壤酶活性和根区微生物多样性,缓解土壤连作障碍的发生,实现优质、高产、可持续栽培是设施园艺研究者和生产者关心的重要问题。
生物质炭是由农业废弃物如秸秆、花生壳等经高温(350~750 ℃)、限氧裂解形成的含碳量丰富、具有独特多孔性结构的物质。相关研究报道,土壤中施用生物质炭可有效改善土壤理化性质,提高土壤的矿质养分含量,增加土壤有机质含量、碳氮比和pH值,提高根际有益菌的多样性[2-3]。前期研究发现,连作土壤中施用小麦秸秆炭可以增加土壤肥力,改善土壤酶活性,促进黄瓜生长,提高产量[4]。然而,目前生物质炭生产成本较高,生产上一般作为炭基肥原料和高附加值园艺作物土壤改良剂使用。微生物菌剂“宁盾”主要是由2种芽孢杆菌和1种沙雷氏菌组成的复合微生物菌剂,对甜瓜枯萎病、番茄青枯病等土传病害均有较好的防治效果[5],此外,施用“宁盾”还可提高土壤微生物多样性,增加植株根围土壤酶活性,对土壤肥力和结构具有良好的改善作用[6]。然而,由于酸性肥料和杀菌剂的频繁施用,导致设施土壤理化性状恶化,微生物菌剂施用后,存在适应性差、促进作物生长及防控土壤病害的效果不明显等问题。
生物质炭发达的多孔性结构、丰富的官能团以及良好的酸碱缓冲能力可以为土壤微生物提供良好的附着空间和繁殖条件,因此与微生物菌肥配施可发挥协同增效作用[7-8]。目前,关于生物质炭和微生物菌剂配施在设施蔬菜栽培中的应用研究报道较少。为探讨生物质炭和微生物菌剂配施改良设施连作土壤、促进作物生长的效果,本研究采用温室盆栽试验研究小麦秸秆炭和微生物菌剂“宁盾”配施对土壤理化性状以及黄瓜生长、产量的影响,分析土壤酶活性和根际或根区可培养土壤微生物数量变化,以期为生产中生物质炭和微生物菌剂的施用提供科学依据。
1 材料与方法
1.1 试验材料
供试土壤取自青岛市农业科学研究院黄瓜育种温室耕作层(0~20 cm),壤质为潮棕壤,试验开始前充分晾干混匀备用。该温室自1996年开始种植黄瓜,每年种植2茬,至取样时已连作黄瓜22年。小麦秸秆炭购自河南三利新能源有限公司,由小麦秸秆在450~550 ℃高温限氧条件下裂解而成,过2 mm筛后使用,其基本性状为:有机碳含量303.3 g/kg、速效氮含量185.60 mg/kg、速效磷含量363.20 mg/kg、速效钾含量6.50 g/kg、全氮含量1.19%、全磷含量0.20%、全钾含量7.75%、pH值9.93、总盐分含量9.94 g/kg。微生物菌剂“宁盾”的主要成分为复合芽孢杆菌,购自南京本源生态农业科技有限公司。试验用植物材料黄瓜翠龙和南瓜砧木14F24,均由青岛市农业科学研究院蔬菜研究所提供。
1.2 试验设计
盆栽试验在青岛市农业科学研究院试验基地玻璃温室内进行。试验采用完全随机区组设计,设连作土壤单施0.25%(m/m)小麦秸秆炭(T1);单施微生物菌剂(T2);0.25%小麦秸秆炭和生物菌剂配施(T3);以未进行任何处理的连作土壤作为对照(T0),每个处理3次重复。小麦秸秆炭在土壤装盆前一次性混匀施入,“宁盾”用水稀释至有效菌含量约为1×107 CFU/mL后,在黄瓜定植时按照1 L/株灌根,对照处理灌同等体积的清水。栽培方式按照笔者所在实验室前期报道的方法[4]进行。采用吊蔓栽培方式,春季于2018年4月4日定植,秋季于2018年8月20日定植,每盆定植黄瓜2株,均匀摆放于温室中,施肥灌溉采用常规水肥一体化的方式进行。春季控制白天平均温度为26 ℃,夜间平均温度为18 ℃,湿度控制在65%~80%范围内;秋季控制白天平均温度为30 ℃,夜间平均温度为25 ℃,湿度控制在65%~85%范围内。
1.3 土壤理化性质及酶活性的测定
土壤理化性质测定参照鲍士旦的方法[9]进行。在黄瓜结瓜期追肥之前,采用多点混合法取地表面5 cm以下的根围土壤充分混匀后,风干过2 mm筛,分别测定其全氮含量、全磷含量、全钾含量、速效氮含量、有效磷含量、速效钾含量、有机质含量、pH值和总盐分含量。土壤酶活性测定参照关荫松等的方法[10]进行,分别在定植后0、20、40 d取根区土壤样品,风干过筛后,利用分光光度法测定不同处理条件下土壤酶活性。
1.4 可培养微生物平板计数
土壤可培养微生物数量采用稀释平板法[11]测定。在黄瓜拉秧时将根系从土壤中轻轻拔出,用无菌小刀把根外大部分土块去除,轻轻抖落松散地附在根上的土粒,收集10 g新鲜土样,加入含有100 mL无菌水的250 mL三角瓶中,130 r/min,振荡15 min,制备成根区(非根际)土壤悬液。取抖落表面土壤的根系10 g,置于含有100 mL无菌水的250 mL 三角瓶中,130 r/min,振荡15 min,制备成根际土壤悬液,用无菌镊子取出根系,滤纸吸干水分,称根质量,洗剂前后质量差即为根际土的质量。将以上2种方法所得土壤悬液按10倍梯度稀释,根际细菌采用稀释至10-6~10-5浓度梯度的悬液接种,根际真菌采用稀释至10-4~10-3浓度梯度的悬液接种;根区细菌采用稀释浓度梯度10-4~10-3浓度梯度的悬液接种,根区真菌采用稀释至10-3~10-2浓度梯度的悬液接种,各取100 μL涂布于平板上,在温度为30 ℃条件下培养2~5 d,统计土壤中可培养微生物数量。
1.5 黄瓜生长、产量和品质的测定
春季、秋季2季分別在定植后30、40、50 d测定不同处理黄瓜植株的株高。在结瓜盛期测定不同处理植株的叶面积。拉秧后整株收获,测定黄瓜地上部鲜质量。结瓜期分批采收商品瓜(瓜条长度18~22 cm,直径3.5~4.0 cm),累计计产。黄瓜产量采用平均单株产量估算,产量=不同处理单株平均产量×52 500。其中52 500为设施生产中1 hm2平均定植黄瓜株数。结瓜盛期不同处理采收均匀一致的10个黄瓜果实,参照王学奎的方法[12]测定黄瓜品质,其中粗蛋白含量采用凯氏定氮法测定;可溶性糖含量采用3,5-二硝基水杨酸比色法测定;维生素C含量采用2,6-二氯靛酚滴定法测定;硝酸盐含量采用水杨酸比色法测定;果实干物质量采用烘干法测定,并计算干物率。
1.6 数据统计
所得数据用“平均值±标准差”表示,采用IBM SPSS 200统计软件进行方差分析,并使用Duncans测验(P<005)比较不同处理数据间的差异显著性。
2 结果与分析
2.1 小麦秸秆炭与微生物菌剂配施对设施土壤养分的影响
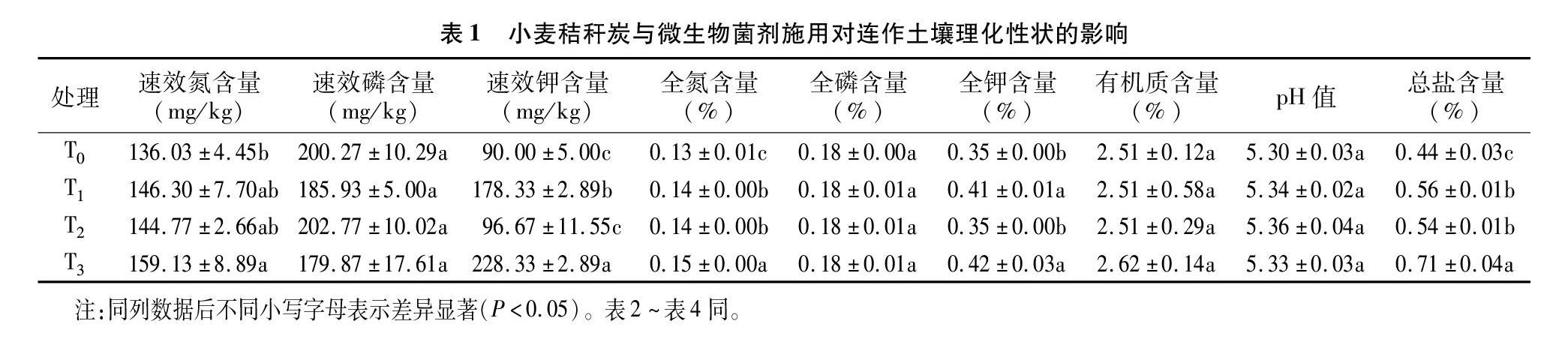
春季黄瓜结瓜期追肥前测定不同处理土壤养分含量从表1可以看出,小麦秸秆炭和微生物菌剂配施处理条件下,土壤中的速效氮含量、速效钾含量、全氮含量、全钾含量与对照相比显著增加;生物质炭(小麦秸秆炭)处理条件下土壤中速效磷含量与对照相比降低,但不同处理间差异不显著。生物质炭和根际促生菌施用后,土壤pH值与对照相比差异不显著,而土壤总盐分含量显著增加,特别是二者配施处理条件下,土壤盐分含量与对照相比增加61.36%,这可能与小麦秸秆炭自身所含盐分含量较高以及微生物菌剂施用促进难溶性矿质养分降解有关。
2.2 小麦秸秆炭与微生物菌剂施用对土壤酶活性的影响
土壤酶活性可在一定程度上反映土壤肥力和土壤微生物代谢情况。秋季分别在黄瓜定植后0、20、40 d测定不同处理条件下的土壤过氧化氢酶、蔗糖酶、脲酶、酸性磷酸酶活性。从图1可以看出,黄瓜生长过程中,土壤过氧化氢酶和脲酶活性总体呈现下降的趋势(图1-B、图1-C),而蔗糖酶活性总体呈现升高的趋势(图1-A),酸性磷酸酶活性随栽培时间延长变化不明显(图1-D)。在T2、T3处理条件下,土壤蔗糖酶活性显著高于对照,而土壤脲酶活性显著低于对照。除定植后40 d外,T2、T3处理条件下土壤过氧化氢酶活性显著高于对照。定植后40 d时,生物质炭和微生物菌剂处理条件下土壤酸性磷酸酶活性显著高于对照处理。
2.3 小麦秸秆炭与微生物菌剂配施对土壤可培养微生物数量的影响
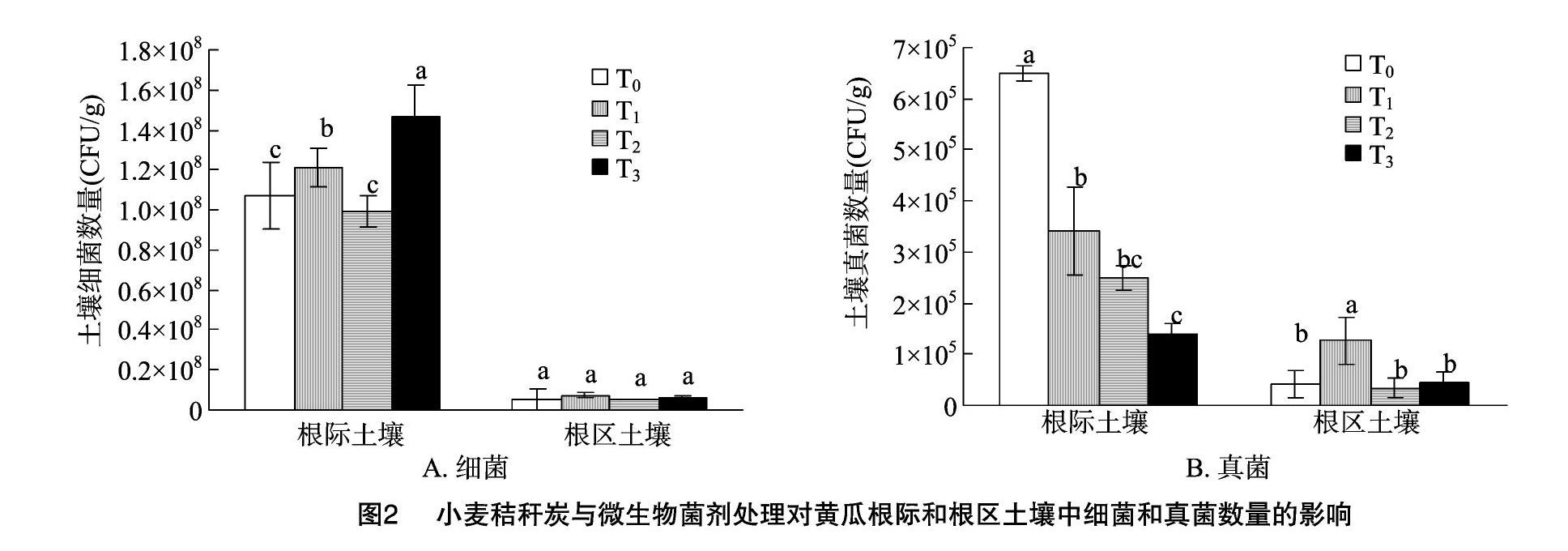
从图2可以看出,设施黄瓜根际土壤中的细菌和真菌数量明显高于根区土壤。与对照相比,T1、T3处理条件下,根际土壤中细菌数量增加,而真菌数量降低。其中以T3处理最为显著,根际土壤中细菌数量显著高于对照,而真菌数量显著低于对照。在根区土壤中,除T1处理的真菌数量显著高于对照外,其他处理条件下可培养细菌和真菌数量与对照相比差异不显著。
2.4 小麦秸秆炭与微生物菌剂对黄瓜生长及产量的影响
2.4.1 小麦秸秆炭与微生物菌剂配施对黄瓜生长的影响
春季在定植后30、40、50 d分别测定黄瓜的株高。结果(表2)表明,连作土壤中施用0.25%小麦秸秆炭可促进黄瓜生长,特别是小麦秸秆炭和微生物菌剂配施处理条件下,黄瓜株高显著高于对照,而单一施用根际微生物菌剂对黄瓜的株高影响不显著。结瓜盛期测定不同处理黄瓜植株主茎最大叶叶面积,T1、T3处理条件下,黄瓜叶面积显著高于对照;T2单施微生物菌剂处理条件下,黄瓜叶面积与对照差异不显著。T3处理条件下,黄瓜地上部鲜质量明显增加,但与对照相比差异不显著。
秋季单施小麦秸秆炭和生物质炭与微生物菌剂配施条件下,黄瓜的株高、叶面积高于对照,但与对照处理相比差异不显著。
2.4.2 小麦秸秆炭与微生物菌剂配施对黄瓜产量的影响
从表3可以看出,单施小麦秸秆炭和单施微生物菌剂处理条件下,春、秋2季黄瓜产量与对照相比差异不显著,然而二者配施条件下黄瓜的增产效果最好,总产量显著高于对照。二者配施条件下,春、秋2季黄瓜的前期产量分别为47.32、60.58 t/hm2,与对照相比分别增加28.13%、4.54%;总产量分别为114.41、85.80 t/hm2,与对照相比分别增加15.89%、12.36%。
2.4.3 小麦秸秆炭与微生物菌剂施用对黄瓜品质的影响 从表4可以看出,生物质炭与微生物菌剂配施条件下,黄瓜果实的维生素C含量显著增加,新鲜果实中维生素C含量为9.94 mg/100 g,比对照增加21.22%。单施微生物菌剂以及生物质炭与微生物菌剂配施均可显著降低果实中的硝酸盐含量,T2、T3处理条件下,果实硝酸盐含量与对照相比分别降低20.19%、37.63%。不同处理条件下黄瓜果实干物率、可溶性总糖含量、粗蛋白含量差异不显著。
3 讨论与结论
秸秆来源的生物质炭含碳量丰富,特别是钾含量较高,因此施入土壤中可增加土壤养分含量,提高肥力。本试验中,小麦秸秆来源的生物质炭,与花生壳炭和稻壳炭相比容重小,因此单位质量相同条件下,其所占体积较大。少量小麦秸秆炭(0.25%)和微生物菌剂配合施用后土壤速效氮含量、速效钾含量、全氮含量、全钾含量较对照显著增加,而土壤全磷、速效磷含量与对照相比差异不显著。然而,生物质炭处理条件下,土壤中速效磷含量与对照相比降低。这可能是由于生物质炭对有效磷具有一定的吸附作用,引起土壤中植物可利用磷含量降低[13]。诸海焘等报道,黄瓜对土壤中氮磷钾养分的吸收比例一般为1 ∶0.48 ∶1.34[14],然而,设施黄瓜生产中,由于长期不平衡施肥导致土壤中的磷大量累积,土壤中的磷肥处于过剩状态[15]。因此,设施土壤中施用生物质炭可吸附一部分土壤中的磷素,减少磷的流失,提高磷肥的利用效率。
设施栽培的集约化特性以及长期的过量和不平衡施肥导致设施土壤极容易发生次生盐渍化现象,盐渍化土壤的盐分组成以NO3-、SO42-、Ca2+为主。优化施肥,降低设施土壤盐分含量,是设施栽培的重要目标。本研究结果表明,设施土壤中施用生物质炭和微生物菌肥可显著提高连作土壤的总盐含量,一方面因为小麦秸秆炭自身的可溶性钾、速效氮、速效磷等养分含量较高[8],引起土壤中可溶性盐分含量升高;另一方面可能是复合芽孢杆菌菌剂的施用,促进了土壤有益微生物的大量繁殖,加速土壤中难溶性养分的分解[16-17]。因此,设施生产中应适量施用生物质炭,以免引起土壤中可溶性总盐分过量积累,降低作物产量和品质。
土壤微生物区系的恶化失调是设施土壤连作障碍发生的重要原因[18]。设施黄瓜连作可导致土壤由细菌型向真菌型转变,根际微生物平衡状态被破坏,特别是引起植物病原真菌的积累[19]。本研究发现,生物质炭和微生物菌剂处理对黄瓜根际土壤微生物的数量影响较大,特别是在二者配施条件下,根际土壤中细菌数量显著高于对照,而真菌数量显著低于对照,类似的现象在水稻栽培上也有报道[20]。首先,生物质炭自身的吸附作用及独特的理化性质,可改善土壤根际微生物数量及群落结构。Egamberdieva等研究發现,土壤中施用2%通过水热方法制备的生物质炭可显著提高大豆根际微生物的多样性,并且使根际促生细菌的比例增加,从而证明了生物质炭可以间接通过改变根际微生物群落结构,增加根际有益微生物,进而促进作物生长[3]。其次,微生物菌剂和小麦秸秆炭配施增加了根部土壤细菌数量,而这些有益菌对部分土壤病原真菌又有直接的拮抗作用,因此,可减少真菌的数量[21]。
生物质炭作为一种新型的土壤改良剂,对黄瓜和番茄等多种设施蔬菜具有促进生长和增产的作用[22-24]。本研究发现,连作土壤中施用小麦秸秆炭可促进黄瓜生长,增加产量。特别是小麦秸秆炭和微生物菌剂配施对黄瓜的增产效果优于单一施用小麦秸秆炭,表现出协同效应。这一方面是由于生物质炭独特的结构,可改善土壤容重,增加土壤肥力和持水力,促进黄瓜的生长。另一方面,本试验所用复合芽孢杆菌菌剂,可改善植株根围土壤理化性状,增加根围土壤肥力,进而促进黄瓜吸收养分和生长发育。此外,微生菌剂对土壤中的枯萎病等病原菌会产生拮抗作用,减少土传病害的发生,间接地提高作物产量[5]。生物质炭和微生物菌剂配施条件下,生物质炭的多孔隙结构,能够为微生物提供附着位点和较大的生存空间。同时,生物质炭所含有的易分解有机物质可以为微生物提供碳源和氮源。因此,二者配施可以促进微生物菌剂在根际土壤中定殖,提高菌体的成活率,发挥协同增效作用。
维生素C、硝酸盐含量是评价蔬菜品质的重要指标,施用生物质炭或生物质炭基肥可以提高蔬菜中的维生素含量,降低硝酸盐含量[25]。本研究也发现,在连作土壤中添加生物质炭和微生物菌肥处理条件下,黄瓜果实维生素C含量明显高于对照,而硝酸盐含量与对照相比明显降低。生物质炭可通过影响植物对养分的吸收,影响维生素C和硝酸盐的合成或代谢途径。刘晓雨等研究表明,生物质炭施用后小白菜叶片硝酸盐含量与钾含量呈极显著的负相关关系,随着小白菜体内钾素的增加硝酸盐含量降低[26],这是因为钾离子能够诱导硝酸盐还原酶的合成,增强其活性,有利于硝酸盐还原。而微生物菌剂可直接降低植物根际土壤中硝酸盐含量,减少植物对硝酸盐的吸收。从而减少果实中硝酸盐的积累[27]。
综上所述,黄瓜连作土壤中小麦秸秆炭和微生物菌剂配施可增加连作土壤养分含量,改善土壤酶活性,增加根际土壤可培养细菌数量且降低根际真菌数量,促进黄瓜生长,提高产量及品质。
参考文献:
[1]李天来,杨丽娟. 作物连作障碍的克服——难解的问题[J]. 中国农业科学,2016,49(5):916-918.
[2]Gul S,Whalen J K,Thomas B W,et al. Physico-chemical properties and microbial responses in biochar-amended soils:mechanisms and future directions[J]. Agriculture Ecosystems & Environment,2015,206:46-59.
[3]Egamberdieva D,Wirth S,Behrendt U,et al. Biochar treatment resulted in a combined effect on soybean growth promotion and a shift in plant growth promoting rhizobacteria[J]. Frontiers in Microbiology,2016,7:1-11.
[4]张功臣,陈建美,赵征宇,等. 生物质炭对设施连作土壤性质及黄瓜生长和产量的影响[J]. 土壤通报,2018,49(3):659-666.
[5]邢卫峰,于侦云,陈刘军,等. 生物肥料“宁盾”对甜瓜枯萎病的防治效果[J]. 江苏农业科学,2014,42(3):78-81.
[6]杨 威,闫海霞,张贝贝,等. 施用微生物菌肥“宁盾”对辣椒根围细菌多样性及土壤酶活性的影响[J]. 江苏农业科学,2018,46(9):99-103.
[7]Nadeem S M,Imran M,Naveed M,et al. Synergistic use of biochar,compost and plant growth-promoting rhizobacteria for enhancing cucumber growth under water deficit conditions[J]. Journal of the Science of Food and Agriculture,2017,97(15):5139-5145.
[8]Akhtar S S,Andersen M N,Naveed M,et al. Interactive effect of biochar and plant growth-promoting bacterial endophytes on ameliorating salinity stress in maize[J]. Functional Plant Biology,2015,42(8):770-781.
[9]鲍士旦. 土壤农化分析[M]. 3版.北京:中国农业出版社,2000.
[10]关松荫,张德生,张志明. 土壤酶及其研究法[M]. 北京:农业出版社,1986.
[11]林先贵. 土壤微生物研究原理与方法[M]. 北京:高等教育出版社,2010.
[12]王学奎. 植物生理生化实验原理和技术[M]. 2版.北京:高等教育出版社,2006.
[13]Xu G,Sun J N,Shao H B,et al. Biochar had effects on phosphorus sorption and desorption in three soils with differing acidity[J]. Ecological Engineering,2014,62:54-60.
[14]诸海焘,李建勇,朱 恩,等. 水肥一体化条件下设施黄瓜的氮磷钾吸收分配规律研究[J]. 上海农业学报,2017,33(1):74-78.
[15]万述伟,张守才,赵 明,等. 设施栽培黄瓜的氮磷钾肥料效应研究[J]. 中国土壤与肥料,2012(5):44-49.
[16]Rai A,Nabti E. Plant growth-promoting bacteria:importance in vegetable production[M]//Microbial strategies for vegetable production. Cham:Springer,2017:23-48.
[17]Pii Y,Penn A,Terzano R,et al. Plant-microorganism-soil interactions influence the Fe availability in the rhizosphere of cucumber plants[J]. Plant Physiology and Biochemistry,2015,87:45-52.
[18]邹春娇,齐明芳,马 建,等. Biolog-ECO解析黄瓜连作营养基质中微生物群落结构多样性特征[J]. 中国农业科学,2016,49(5):942-951.
[19]胡元森,刘亚峰,吴 坤,等. 黄瓜连作土壤微生物区系变化研究[J]. 土壤通报,2006,37(1):126-129.
[20]Chen J H,Liu X Y,Zheng J W,et al. Biochar soil amendment increased bacterial but decreased fungal gene abundance with shifts in community structure in a slightly acid rice paddy from Southwest China[J]. Applied Soil Ecology,2013,71:33-44.
[21]Han L J,Wang Z Y,Li N,et al. Bacillus amyloliquefaciens B1408 suppresses Fusarium wilt in cucumber by regulating the rhizosphere microbial community[J]. Applied Soil Ecology,2019,136:55-66.
[22]韩召强,陈效民,曲成闯,等. 生物质炭对黄瓜连作土壤理化性状,酶活性及土壤质量的持续效应[J]. 植物营养与肥料学报,2018,24(5):1227-1236.
[23]沈 盟,蒋芳玲,王 珊,等. 生物质炭施用量对土壤性状和番茄产质量的影响[J]. 土壤,2017,49(3):534-542.
[24]李 麗,王雪艳,田彦芳,等. 生物质炭对土壤养分及设施蔬菜产量与品质的影响[J]. 植物营养与肥料学报,2018,24(5):1237-1244.
[25]李大伟,周加顺,潘根兴,等. 生物质炭基肥施用对蔬菜产量和品质以及氮素农学利用率的影响[J]. 南京农业大学学报,2016,39(3):433-440.
[26]刘晓雨,刘 成,王贺东,等. 添加生物质炭不同组分对不结球白菜产量和品质的影响[J]. 南京农业大学学报,2018,41(6):1070-1077.
[27]Chu S H,Zhang D,Zhi Y E,et al. Enhanced removal of nitrate in the maize rhizosphere by plant growth-promoting Bacillus megaterium NCT-2,and its colonization pattern in response to nitrate[J]. Chemosphere,2018,208:316-324.
