水稻株型相关性状QTL定位研究
2019-11-28张习春张应洲圣忠华龙武华吴健强朱速松魏祥进
张习春 张应洲 圣忠华 龙武华 吴健强 朱速松 魏祥进


摘要:以粳稻日本晴(NIP)和籼型超级稻中嘉早17(YK17)构建的重组自交系(recombinant inbred line,简称RIL)群体为研究材料,于2015—2016年分3季种植于杭州和海南,成熟期考察株型相關性状(分蘖数、株高、剑叶长、剑叶宽、剑叶长宽比、穗长)。并应用这2种群体已构建的分子连锁图谱,对株型相关性状进行QTL分析,共检测到34个相关QTL,分布于除第9染色体外的其他染色体上,LOD值介于2.68~14.39之间,表型贡献率变幅为4.00%~26.62%,3个环境下重复检测到10个QTL。对QTL与环境互作分析发现,24个QTL与环境互作,其中2个QTL对环境敏感。株型相关性状QTL主要分布在第1、2、4、7染色体上,呈簇分布。qPH1、qLFL1和qLFLW1a在3个环境下重复检测到,且贡献率较高,有待于进行克隆和功能分析。
关键词:水稻;重组自交系(RIL);株型性状;QTL分析
中图分类号: S511.03文献标志码: A
文章编号:1002-1302(2019)18-0102-06
收稿日期:2018-05-28
基金项目:“十二五”农村领域国家科技计划课题(编号:2014AA10A604-11);贵州省科学技术基金(编号:黔科合J字[2015]2098号);贵州省科研机构服务企业行动计划(编号:黔科合服企[2014]4005号);贵州省现代农业产业体系项目(编号:GZCYTX2018-06);贵州省科技重大专项(编号:黔科合重大专项字[2013]6023号);贵州省科技合作计划(编号:黔科合LH字[2015]7074号);黔农科院青年基金(编号:黔农科院青年基金[2018]号)。
作者简介:张习春(1991—),男,贵州织金人,硕士,研究实习员,主要从事水稻遗传育种研究。E-mail:18358168852@163.com。
通信作者:魏祥进,博士,副研究员,主要从事水稻分子遗传学研究。E-mail:weixiangjin@caas.cn。
水稻(Oryza sativa L.)是世界上主要的粮食作物,分别约占我国粮食播种面积与总产量的27%和35%,我国约有60%人口以稻米为主粮,因此,其在保障粮食安全中扮演着重要角色[1]。株型改良是提高产量的有效途径;被誉为第一次“绿色革命”的矮化育种降低了水稻株高,实现了单位面积产量和总产量的大幅提升。近年来,育种家倡导理想株型的超级稻品种选育理念,力求实现水稻产量的新突破,因此开展株型相关性状的遗传研究对理想株型的超级稻品种选育具有重要的理论意义和实际参考价值。
已有研究表明,株型相关性状属于复杂的数量性状,其遗传表达容易受到环境条件的影响。不同群体甚至同一群体控制株型相关性状在不同年份和环境条件下的QTL数量、位置、加性效应值和贡献率可能出现明显的差异[2-3]。张玲等选用泸灰99和沈农265的RIL群体,株高、穗长、剑叶长、剑叶宽在2个环境下分别检测到4、2、2、5个QTL[4];林泽川利用BG1和XJL的RIL群体,分别检测到株高、穗长、剑叶长、剑叶宽性状的QTL分别是2、5、5、8个,两者报道的数量和位置都存在较大差异[5]。结合其他研究[6]发现:同一性状在不同遗传群体和环境条件下检测到的QTL可能存在明显差异,部分QTL重复检测性极低,主效QTL具有较高可重复性,因此有必要使用不同遗传群体开展相关研究,寻找能在不同群体中重复出现的QTL位点,为理想株型选育提供理论依据。
QTL定位是克隆基因的重要方法,目前已克隆到几个调控株型性状的重要基因,如IPA1(OsPL14)是1个克隆自少蘖粳稻的半显性QTL,该基因调控水稻分蘖发生,同时影响水稻株高和穗长[7]。Huang等从沈农265中分离的DEP1是1个控制水稻产量性状的主效QTL,它能调控水稻穗长、穗弯曲度、着粒密度[8]。这些已克隆的基因初步阐明部分株型性状的遗传机制,为水稻株型改良种提供理论依据和基因聚合材料,进一步挖掘相关基因能为育种工作提供更加丰富和准确的选择。
目前已克隆控制株型相关性状的基因较少,不足以揭示水稻株型复杂的遗传机制以及满足高产育种应用需求。本研究利用日本晴和中嘉早17构建的重组自交系群体,发掘在不同环境下能稳定表达的株高、分蘖和穗长等相关性状的QTL,为揭示水稻株型复杂的遗传机制和理想株型高产育种应用提供理论依据。
1 材料与方法
1.1 RIL群体构建
以日本晴(Nipponbare,NIP)为母本、中嘉早17(Zhongjiazao17,YK17)为父本,杂交产生F1代,使用单粒连续自交,构建性状覆盖度广且遗传相对稳定的200个株系重组自交系群体(recombinant inbred lines,简称RILs)。
1.2 材料种植及性状考察
2地3季种植群体亲本和200个RIL株系:2015年夏季种植于杭州富阳(E1表示),2015年冬季种植于海南陵水(E2表示),2016年夏季种植于杭州富阳(E3表示)。亲本和群体每个株系种植8行×8列,田间管理同常规大田管理。成熟时,于每个区块中部选取5株进行考种,具体参考Moncada等提出的方法[9]分别对分蘖数(tiller number per plant,简称TN)、株高(plant height,简称PH)、剑叶长(flag leaf length,简称LFL)、剑叶宽(flag leaf width,简称LFW)、剑叶长宽比(flag leaf length width rate,简称FLWR)、穗长(panicle length,简称PL)6个农艺性状进行考察,取5株的平均值。
1.3 遗传连锁图谱
利用629对SSR引物筛选NIP和YK17 2个亲本间的多态,挑选出163个均匀分布在水稻12条染色体的多态性标记,检测群体基因型,获得163个标记的基因型数据。应用MAPMARKER/EXP V3.0进行连锁分析,以LOD≥3.0进行标记分组,利用Kosambi函数将重组率转为图距,利用MapDraw 2.1绘制染色体,构建连锁图谱。连锁图谱总图距约为1 479.40 cM,覆盖水稻基因组的93.85%,标记间平均图距为9.08 cM。
1.4 QTL定位分析
利用Windows QTL Cartographer 2.5软件复合区间作图法(CIM),每1 cM步长进行全基因组扫描,检测目标性状QTL。以Permutation 1 000次模拟运算确定LOD阈值,即当实际检测到某个区段LOD值大于LOD阈值时,则认为该区段存在1个QTL,同时软件运算给出对应该QTL的加性效应值和表型贡献率[10]。检测到的QTL遵循McCouch等QTL命名原则命名[11]。利用QTL Network 2.0软件,基于混合模型的复合区间作图法(MCIM),分析QTL与环境互作效应[12]。
2 结果与分析
2.1 RIL群体农艺性状表型分析
由表1、图1可知,RIL群体中不同株系在株型相关性状中具有亲本多种重组类型,考察的6个性状呈连续性分布,表现为双向超亲分离,表明这些性状由多个数量性状基因座位控制;3个环境下峰度和偏度绝对值除了剑叶长宽比峰值在E1中大于1,其余均小于1,遵循正態分布,符合QTL分析的作图要求。
2.2 RILs群体各性状间的相关性
由表2可知,在3个环境下分蘖数与株高、剑叶长、剑叶宽、穗长呈极显著负相关;株高、剑叶长、剑叶宽和穗长4个性状间呈显著或极显著正相关;剑叶长宽比与剑叶宽呈负相关,与株高、剑叶长、穗长呈极显著正相关。说明株型各性状之间密切相关,同时在不同环境下相关系数存在一定差异,表明遗传因素是调控株型形成的主要因素,环境条件对株型形成具有一定的影响。
2.3 株型性状QTL分析结果
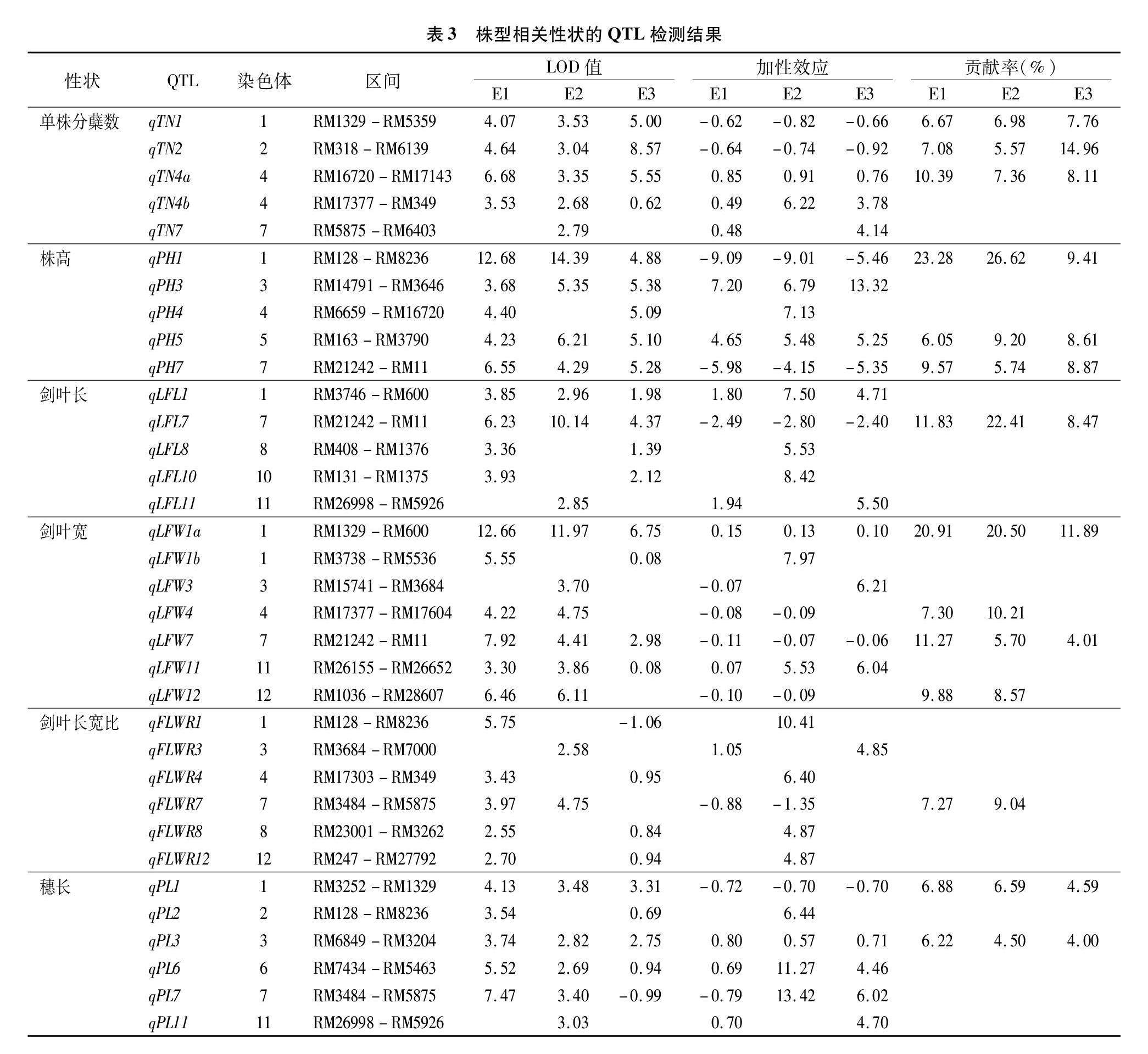
对不同环境下的6个株型相关农艺性状表型值进行全基因组关联分析,共检测到34个QTL,分布于除第9染色体外的11条染色体上,LOD值介于2.55~14.39之间,表型贡献率变幅为3.78%~26.62%。E1、E2、E3的环境下分别检测到23、18、24个QTL,3个环境重复检测到11个QTL(表3)。具体如下:
分蘖数(TN):3个环境下共检测到5个控制分蘖数QTL,分布在第1、2、4、7染色体上,LOD值介于2.68~8.57之间,贡献率变幅为3.78%~14.96%。其中,qTN1、qTN2和qTN4a在3个环境下重复检测到,平均贡献率为7.14%、920%、862%。qTN1和qTN2加性效应为负值,增效等位基因来自NIP,qTN4a加性效应为正值,增效等位基因来自YK17。
株高(PH):3个环境下共检测到5个控制株高QTL,分布于第1、3、4、5、7染色体上,LOD值介于3.68~14.39之间,贡献率变幅为5.74%~26.62%。其中,qPH1、qPH5、qPH7在3个环境下重复检测到,平均贡献率分别为19.80%、7.95%、8.06%,qPH1和qPH7加性效应为负值,增效等位基因来自NIP,qPH5加性效应为正值,增效等位基因来自YK17。
剑叶长(LFL):3个环境下共检测到5个剑叶长QTL,位于第1、7、8、10、11染色体上,LOD值介于2.85~10.14之间,贡献率变幅为4.71%~22.41%。其中qLFL7在3个环境下
重复检测到,平均贡献率14.24%,加性效应为负值,增效等位基因来自NIP,其余4个QTL加性效应为正值,增效等位基因来自于YK17。
剑叶宽(LFW):3个环境下共检测到7个剑叶宽QTL,分布于第1、3、4、7、11、12染色体上,LOD值介于3.30~12.66之间,贡献率变幅为4.01%~20.91%。其中qLFW1a和qLFW7在3个环境下重复检测到,平均贡献率分别为17.77%和699%;qLFW1a的加性效应为正值,增效等位基因来自中YK17,qLFW7的加性效应为负值,增效等位基因来自NIP。
剑叶长宽比(FLWR):3个环境下共检测到6个剑叶长宽比QTL,分布于第1、3、4、7、8、12染色体,LOD值介于2.55~5.75,贡献率变幅为4.85%~10.41%,未检测到在3个环境下重复出现的QTL。
穗长(PL):3个环境下共检测到6个穗长QTL,分布于第1、2、3、6、7、11染色体上,LOD值介于2.69~7.47之间,贡献率变幅为4.00%~13.42%。qPL1和qPL3在3个环境下重复检测到。qPL1加性效应为负值,增效等位基因来自NIP,qPL3加性效应为正值,增效等位基因来自YK17。
2.4 QTL与环境互作
应用QTL Network 2.0的MCIM分析方法联合检测株型相关性状与环境互作效应,共检测到24个QTL与环境互作(表4),占QTL总数的70.59%,其中控制株高性状的qPH1在E3环境下互作效应达到显著,控制穗长性状的qPL6在E2和E3环境下互作效应达到显著;其余QTL与环境互作均未达到显著,表明环境对大部分QTL遗传产生不同程度的影响,株高和穗长性状更易受环境影响。
3 讨论和结论
水稻株高、剑叶长、剑叶宽、穗部性状等性状遗传机制较为复杂,易受环境条件的影响,QTL重复检测率低。姚晓云等选用沈农265和丽江黑谷构建RIL群体对株高和穗长进行QTL检测,3年3个环境下检测的9个QTL中仅有3个可重复检测到[13];张斌利用两优培九和培矮64的RIL群体,在不同环境下检测到20个株型相关的QTL中只有5个可重复[14]。本研究中,6个株型相关性状在3个环境下共检测到36个QTL,其中qTN1、qTN2、qTN4a、qPH1、qPH5、qPH7、qLFL7、qLFW1a、qLFW7、qPL1和qPL3在3个环境下能重复检测,分别占单个环境QTL总数的39.13%、50.00%、37.50%,表明这些QTL具有稳定的重复性。此外,分析与环境的互作效应发现大部分株型相关性状的QTL与环境互作,但只有qPH1和qPL6对环境敏感,说明3个环境下重复检测到的QTL受环境影响有限,具有较好地遗传稳定性,在不同环境下能稳定表达,可进一步进行基因克隆和功能分析。
本试验检测到株型性状的部分QTL与已报道的结果位于相同或相近的区间。如位于第1染色体标记RM128-RM8236之间的qPH1,该区间内包含已克隆的矮秆基因sd1和d61,推测该QTL较高的表型贡献率与该区间内sd1和d61功能有关[15-16];qPH5和qPH7的区间分别包含影响株高的EUI1和Ghd7[17-18]。qLFL7与李睿等报道的qLFL7-1 位于相同区间[19];qLFW1a区间包含影响叶宽的Osa-miR319b[20];qTN2区间包含调控有效穗数的MADS57[21];qPL3区间包含影响穗长OsPHR1[22]。部分重复检测到的QTL区间内包含已克隆控制相关株型性状的基因,这些基因可能对相应QTL位点较高的遗传效应具有重要作用。qTN4a、qLFL1a和qPL1在3个环境下重复检测到,具有较好的遗传稳定性,区间尚未报道克隆相关基因,有必要进一步进行精细定位和基因功能分析。
QTL多效性是指多个性状表型受到同一QTL的影响,这种情况在作物中是普遍存在的。圣忠华等在第10染色体标记RM1375-RM25664之间检测到控制株高、剑叶长、剑叶宽、穗长和有效穗数的QTL[23];占小登等在第5染色体标记RM500-RM18之间检测到控制株高和穗长的QTL[24]。本试验使用日本晴和中嘉早17的RIL群体检测到控制株型相关性状的QTL主要分布在第1、3、4和7染色体上,多表现为成簇分布。在第1染色体标记RM1329-RM600之间同时检测到控制剑叶长和剑叶宽且效应值较大的QTL;位于第7染色体标记RM21242-RM11之间同时检测到控制株高、剑叶长、剑叶宽、剑叶长宽比、穗长的QTL。这些多效QTL位点对株型塑造具有整体协同作用,有待深入研究,为解释株型遗传机理和培育理想株型品种奠定理论基础。
参考文献:
[1]Zhang Q F. Strategies for developing green super rice[J]. Proceedings of the National Academy of Sciences of the United States of America,2007,104(42):16402-16409.
[2]彭茂民,杨国华,张菁晶,等. 不同遗传背景下水稻剑叶形态性状的QTL分析[J]. 中国水稻科学,2007,21(3):247-252.
[3]赵建国,蒋开锋,杨 莉,等. 水稻产量相关性状QTL定位[J]. 中国水稻科学,2013,27(4):344-352.
[4]张 玲,李晓楠,王 伟,等. 水稻株型相关性状的QTL分析[J]. 作物学报,2014,40(12):2128-2135.
[5]林泽川. 水稻株型相关性状的QTL定位[D]. 北京:中国农业科学院,2013.
[6]王 兰,李 智,郑杏梅,等. 普通野生稻矮化突变体的株高与分蘖基因的QTL定位及主效基因的遗传分析[J]. 华北农学报,2014,29(5):5-9.
[7]Jiao Y,Wang Y,Xue D,et al. Regulation of OsSPL14 by OsmiR156 defines ideal plant architecture in rice[J]. Nature Genetics,2010,42(6):541-544.
[8]Huang X Z,Qian Q,Liu Z B,et al. Natural variation at the DEP1 locus enhances grain yield in rice[J]. Nature Genetics,2009,41(4):494-497.
[9]Moncada P,Martínez C P,Borrero J,et al. Quantitative trait loci for yield and yield components in an Oryza sativa × Oryza rufipogon BC2F2 population evaluated in an upland environment[J]. Theoretical and Applied Genetics,2001,102(1):41-52.
[10]Li J,Wang S C,Zeng Z B. Multiple-interval mapping for ordinal traits[J]. Genetics,2006,173(3):1649-1663.
[11]Blair M W,Panaud O,McCouch S R. Inter-simple sequence repeat (ISSR) amplification for analysis of microsatellite motif frequency and fingerprinting in rice (Oryza sativa L.)[J]. Theoretical and Applied Genetics,1999,98(5):780-792.
[12]Yang J,Hu C C,Hu H,et al. QTLNetwork:mapping and visualizing genetic architecture of complex traits in experimental populations[J]. Bioinformatics,2008,24(5):721-723.
[13]姚曉云,李 清,刘 进,等. 不同环境下水稻株高和穗长的QTL分析[J]. 中国农业科学,2015,48(3):407-414.
[14]张 斌. 不同环境下水稻剑叶形态与产量相关性状的QTL分析[D]. 北京:中国农业科学院,2014.
[15]Sasaki A,Ashikari M,Ueguchi-Tanaka M,et al. Green revolution:a mutant gibberellin-synthesis gene in rice[J]. Nature,2002,416(6882):701-702.
[16]Yamamuro C,Ihara Y,Wu X,et al. Loss of function of a rice brassinosteroid insensitive1 homolog prevents internode elongation and bending of the lamina joint[J]. Plant Cell,2000,12(9):1591-1606.
[17]Zhang Y Y,Zhu Y Y,Peng Y,et al. Gibberellin homeostasis and plant height control by EUI and a role for gibberellin in root gravity responses in rice[J]. Cell Research,2008,18(3):412-421.
[18]Xue W,Xing Y,Weng X,et al. Natural variation in Ghd7 is an important regulator of heading date and yield potential in rice[J]. Nature Genetics,2008,40(6):761-767.
[19]李 睿,赵姝丽,毛 艇,等. 水稻剑叶形态性状QTL分析[J]. 作物杂志,2010(3):26-29.
[20]Wang S T,Sun X L,Hoshino Y,et al. MicroRNA319 positively regulates cold tolerance by targeting OsPCF6 and OsTCP21 in rice (Oryza sativa L.)[J]. PLoS One,2014,9(3):e91357.
[21]Guo S Y,Xu Y Y,Liu H H,et al. The interaction between OsMADS57 and OsTB1 modulates rice tillering via DWARF14[J]. Nature Communications,2013,4:1566.
[22]Guo M N,Ruan W Y,Li C Y,et al. Integrative comparison of the role of the PHOSPHATE RESPONSE1 subfamily in phosphate signaling and homeostasis in rice[J]. Plant Physiology,2015,168(4):1762-1776.
[23]圣忠華,朱子亮,马 宁,等. 超级稻品种中嘉早17产量相关性状QTL定位研究[J]. 中国水稻科学,2016,30(1):35-43.
[24]占小登,于 萍,林泽川,等. 利用大粒籼/小粒粳重组自交系定位水稻生育期及产量相关性状QTL[J]. 中国水稻科学,2014,28(6):570-580.
