体质人类学视角下的人类耐力跑步假说
2019-11-26任占兵吴进龙陈小英
任占兵 吴进龙 陈小英
摘 要:通過对人类耐力跑步相关的人体形态学和生活方式演变等文献进行回顾与分析,通过比较的方法,深入分析人类与灵长类黑猩猩的身体结构、肌肉结构以及生物力学等特征,并进一步从原始人类饮食结构、狩猎方式以及觅食距离特征阐释人类耐力跑步假说。人类耐力跑步可以追溯到人类的起源与进化的整个过程,人类耐力跑步的形成是一个极其复杂的演化过程,受到人类遗传、生物变异以及自然选择等多种因素的共同影响。耐力跑步的形态学基础主要体现在人类进化过程中逐渐形成更适合跑步的后颅特征、下肢更长、脚趾指骨更短、足弓较高且弹性更好、髋关节变窄、臀大肌变得更加宽大等;耐力跑步的生活方式演变因素主要体现人类饮食结构的变化,肉食的出现以及肉类作为饮食的重要比例开始增加、持久性的耐力狩猎方式出现、人类觅食距离的增加等。体质人类学视角下的耐力跑步假说具有一定的合理性,现代社会耐力跑步热现象有其形成的进化根源,在人类进化的过程中,自身形态结构的适应性变化是耐力跑步假说成立的内在原因;人类生活方式的演变及对环境的适应是耐力跑步假说成立的外在原因。
关 键 词:体质人类学;耐力跑步假说;人类进化
中图分类号:G822.3 文献标志码:A 文章编号:1006-7116(2019)05-0086-10
Abstract: By reviewing and analyzing the literature on human morphological and lifestyle evolution in relation to human endurance running, by means of comparison, the authors carried out an in-depth analysis on the body structures, muscle structures and biomechanical characteristics of human beings and primate chimpanzees, and further interpreted the human endurance running hypothesis from the perspectives of the characteristics of the diet structure, hunting manner and foraging distance of primitive human beings. Human endurance running can be traced back to the entire process of human origination and evolution; the formation of human endurance running was an extremely complicated process of evolution, subject to the joint influence of various factors such as human heredity, biomutation and natural selection. The morphological foundation of endurance running was mainly embodied in the post-cranial characteristic, longer lower limbs, shorter toe phalangeal bones, higher and more elastic foot arch, narrower hip joint and wider gluteus maximus, which were more suitable for running, gradually formed in the process of human evolution; endurance running lifestyle evolution was mainly embodied in the change of the human diet structure, the appearance of meat and meat as an important proportion of diet starting to increase, the appearance of the way of persistent endurance hunting, the increase of human foraging distance, etc. From the perspective of physical anthropology, the endurance running hypothesis has certain rationality, the phenomenon of the endurance running enthusiasm in modern society has the evolution root for its formation; in the process of human evolution, the adaptive change of humans own morphological structure is the intrinsic reason for the establishment of the endurance running hypothesis; human lifestyle evolution and human adaptation to environments are the extrinsic reasons for the establishment of the endurance running hypothesis.
Key words: physical anthropology;endurance running hypothesis;human evolution
耐力跑步(Endurance Running)主要是指以有氧代谢为主持续运动一段时间或距离的跑步形式。现代科学研究发现,耐力跑步对人类身心有诸多益处,例如,它可以有效降低人体心血管系统疾病的风险[1-2],降低高血压患者的死亡率[2],改善睡眠、情绪和注意力[3]等。近年来,随着人民健康意识的增强,参加耐力跑步的人数开始迅速增加,参加马拉松赛事的人数也逐年增长。根据中国田径协会的数据,我国马拉松赛事的跑步人数由2010年的40万人增加到2018年的530万人,越来越多的人实现了从不跑步到跑完全程马拉松的人生跨越,将实现个人目标和生命意义作为马拉松跑步的主要动机[4]。人类天生的耐力跑步能力不仅是为了自身的生存而对自然界所产生的一种适应,更是人类为了追求自身生命价值的主体性得以回归的表现[5]。随着耐力跑步运动风靡全球,科学家们也开始从体质人类学的视角追溯人类耐力跑步的渊源,探索耐力跑步对人类发展的意义和价值。
体质人类学是从生物和文化的视角来研究人类体质特征在时间和空间上的变化及其发展规律的科学[6-7],体质人类学着眼于人的群体,研究范围涉及到人类在自然界的位置、人类的起源和发展、人类与动物特别是灵长类的联系和区别等[8-10]。在人类的起源与发展过程,双足直立运动是人类进化的关键标志,是人类为了适应自然环境生存而不得不对自身的运动方式进行的结构生优化。而关于人类为什么要选择双足直立运动的解释,仍然是众说不一,至今大约有30多个不同的版本,比如,类似于猩猩在细树枝上站立觅食行为假说[11]、使用工具假说[12]、运动经济性假说[13]等。通过对古人类化石的分析以及与其他灵长类的比较,科学家们提出了人类进化的耐力跑步假说[14-15],认为耐力跑步是200万年以来人类在演化过程中延续下来的一种特殊生存能力,双足直立的耐力跑步形式可以使人类进行长距离的狩猎并获取食物,并且可以使人类的跑步能量消耗更加节省化,因此,在人类进化过程发挥了重要作用。
人类是遵循生物进化法则从自然中孕育出来的,并且同样受着进化规律的支配[16],人类的直立行走不仅可以减少能量消耗、解放双手、开阔视野,而且引发了人类体质形态学上的巨大改变,是形成人类独特身体结构的重要机制[17]。从体育人类学的视角,双足直立运动为体育性质的身体活动奠定了生物学基础[18]。在人类进化的过程中,身体永远处于进化的最前沿,双足直立运动是体育运动基本技能得以呈现的基础,因此,走、跑、跳、投等身体行为是人类进化过程的结晶和成果[19]。從体质人类学的视角审视耐力跑步假说,可以发现因进化而导致的人类种族特征与运动表现的关系[20],可以为竞技体育的发展、全民健身的实施以及国民健康的改善提供理论依据[17]。耐力跑步假说为从体质人类学的视角追溯人类进化的规律提供了一个新的视角,从体质人类学的视角回顾人类演化的耐力跑步假说及相关研究进展,无论是对解释人类双足直立运动的演化,还是对体育理论的发展都具有重要价值。
1 人类耐力跑步假说的提出
人类双足直立运动是最适合人类的运动方式吗?人体是否为耐力跑步而设计直立运动呢?这些问题看起来似乎很奇怪,但是已经争论了几十年。从这些争论中产生了两个主要的观点:一种观点认为,双足直立运动并不是最适合人类的运动方式,这种运动方式相对哺乳动物的四足运动,能量消耗更大,运动经济性更低。比如,通过比较人类与黑猩猩的研究发现,人类双足跑步能耗是四足哺乳动物跑步能耗的2倍多[21-22];另一种观点认为,双足直立运动相比灵长类黑猩猩四足运动能量更加节省、运动经济性更好,这种运动方式最适合人类。比如,在相同体重的情况下,人类双足行走11 km所消耗的能量相当于雄性黑猩猩四足行走4 km所消耗的能量,即人类双足直立运动通过能量节省化进而延长了人类行走的距离[13]。坚持这一观点的学者认为,通过双足直立的方式,早期原始人可以长途跋涉寻找水、食物和其他资源,同时还可以逃避捕食者[23-25]。
而关于“人体是否为耐力跑步而设计?”1984年,美国密歇根大学的Carrier[14]根据前人的研究提出人类进化的耐力跑步假说(Endurance Running Hypothesis),他认为,人类良好的汗腺发育、较少的体毛、高效的体温调节、良好的呼吸机制以及持续性的捕猎能力等塑造了人类独特的耐力跑步能力,这种耐力跑步能力对人类的演化具有重要意义。2004年,研究人员通过对古人类化石的分析,进一步肯定了耐力跑步假说,他们认为,耐力跑步在200万年以来人类进化中的确发挥了重要作用,人类的演化可以在一定程度上被解释为对耐力跑步的适应。人类的耐力跑步能力与其他灵长类动物相比,在塑造人类谱系方面起着关键作用,并且这一观点得到了人体解剖学研究的支持[15]。耐力跑步假说认为,人类之所以天生就会跑主要归因于人类自身的解剖结构优势、体温调节功能以及饮食结构的变化等因素[15,26]。除此之外,耐力跑步假说还认为,自从人类形成了双足直立的运动方式以后,下肢逐步演化成为身体活动的主要支撑器官,短而直的脚趾、高而有弹性的足弓、修长而发达的跟腱、发达的臀部肌肉等奠定了人类似乎天生拥有耐力跑步的优势[18]。另外,1980年人类学家通过比较黑猩猩与人类的能量消耗发现,人类的双足直立运动在进化过程中表现出了能量节省化的优势[27],这也为人类耐力跑步奠定了生物能量学基础。本研究重点阐述人类身体结构的适应性变化以及生活方式演变对人类耐力跑步假说形成的内在原因和外在原因。
2 人类身体结构的适应性变化是耐力跑步假说的内在原因
追溯适应人类耐力跑步的身体结构特征主要基于两种路径,一种是基于出土的古人类化石,通过身体形态学测量来推断人类身体结构与耐力跑步的关系;另一种是基于人类与黑猩猩等灵长类动物的比较,根据测量结果推断人类适合耐力跑步的优势。虽然人类与黑猩猩的基因差异很小,但是运动方式却差异显著。黑猩猩等灵长类动物的身体解剖学结构决定了其拥有良好的爬树技能却不擅长耐力跑步,虽然可以迅速冲刺,但是这种能力也只能局限在百米左右,距离很短[28-29]。相比之下,人类在灵长类动物中拥有独特的耐力跑步能力,比如,人类可以持续跑完马拉松(42.195 km),而黑猩猩却很难完成,人类这种耐力跑步能力在一定程度上归因于人类在进化过程中所形成的特殊身体结构。研究人员通过对早期原始人类化石的形态结构特征分析表明,伴随着人类开始狩猎等生活方式的演化,人类的身体结构也发生着适应性的变化,行走或跑步更加经济,更有利于人类的生存和发展。在人类进化的过程中,人类身体结构在遗传、生物变异以及自然选择等作用下逐渐发生了适应耐力跑步的变化,这种适应性变化主要体现在肌肉形态、人体骨关节和环节等。
2.1 人类肌肉结构适应性变化支持耐力跑步假说
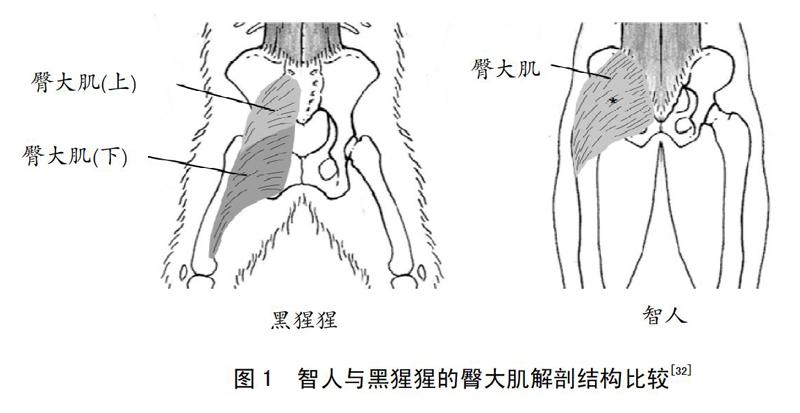
人类适合耐力跑步的肌肉形态适应性变化主要体现在人类的臀大肌变得更加宽大。直立人(Homo erectus)生活在距今大约180万年到200万年前,考古学家对肯尼亚图尔卡纳东部出土的直立人髋骨化石进行了研究,结果发现,直立人的骨盆已经开始变得相对较窄[30]。研究人员进一步根据直立人的骨盆结构变化,推测出原始人类的直立运动逐渐使臀大肌开始变得更加宽大[30-31]。智人生活在距今大约190万年到40万年前,相比直立人,智人更接近现代人类。研究人员进一步比较了智人与黑猩猩的臀大肌形态学结构,探索臀大肌在跑步过程中的作用[32],结果发现,双足直立运动的智人要比四足爬行的黑猩猩的臀大肌更加宽大(见图1)。
黑猩猩的臀大肌主要由上下两部分构成,结构细长且较窄,相比之下,智人没有像黑猩猩一样细长的臀大肌下部,智人臀大肌的这种结构特点也许是人类长期直立运动所造成的结构性适应,并且为人类耐力跑步奠定了基础。作为髋关节的伸展肌群,在跑步过程中臀大肌产生的力作用于骨盆,可以抵抗由于自身体重作用造成的躯干过渡前倾。臀大肌通过稳定跑步过程的躯干前倾,减缓摆动腿的速度,进而保持身体的平衡性。研究人员通过肌电测试观察了现代人在跑步过程中臀大肌的激活变化,结果发现,跑步过程中人类的臀大肌激活程度相比走路更加明显,臀大肌在跑步中的明显激活,说明了臀大肌对人类双足直立跑步不仅产生了结构性适应,而且也产生了功能性适应[32]。
2.2 人类身体环节适应性变化支持人类耐力跑步假说
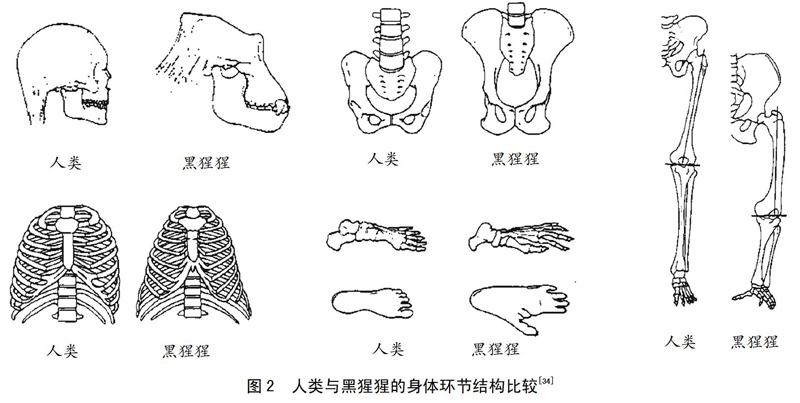
与黑猩猩相比,人类的身体环节出现了更适合双足直立行走及跑步的特征(见图2):头颅靠后、颈部较长,更适合奔跑;胸廓明显扩张,可能意味着呼吸功能增强;骨盆相对较短,呈盆状;下肢增长。脚趾相对较短,足弓更加明显,高而富有弹性的足弓可以使人类在跑步过程中拥有良好的缓冲性能,较短的脚趾减少了人类在跑步支撑阶段的能量消耗,下肢较大的关节面以及明显增大的跟骨结节等可以改善跑步经济性[15]。南方古猿被认为是世界上最早的原始人类,是正在进化中的人类晚期人科动物,是从猿类到人类进化的第一阶段,分布在迄今大约360万年前的非洲北部和东部地区。从化石分析表明,南方古猿已经习惯于在陆地进行双足直立运动,但是,南方古猿保留了与树栖运动相关的身体环节形态学特征,包括手臂较长、下肢较短、趾骨長而弯曲、肩部较窄、躯干呈漏斗状、肩胛骨关节窝面向上方等[33]。另外,早在更新世(约250万年前)人的脑容量开始迅速增大之前,早期南方古猿就拥有了适合双足直立运动的后颅形态。总体上,从人类与黑猩猩身体环节的解剖结构可以发现,人类双足直立运动以后身体结构发生了显著变化,这种改变更有利于降低人类长距离跑步的能量消耗,人类耐力跑步的优势更加明显。
1)人类足弓结构适应性变化支持耐力跑步假说。
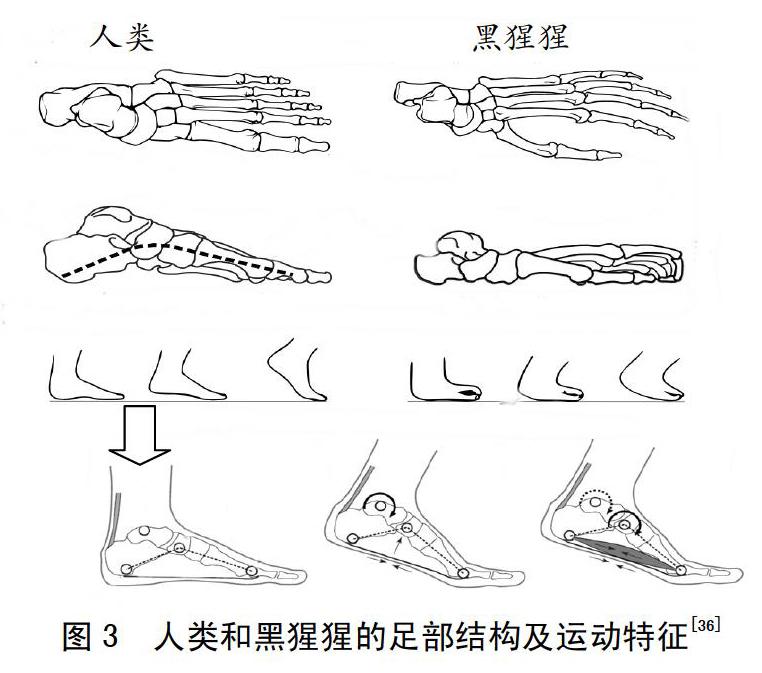
人类的双足在结构上有一个纵向足弓及较短的足趾,这与所有其他灵长类动物的足有本质区别[35]。纵向足弓在结构上主要由跗骨和跖骨以及周围的韧带和肌腱共同组成,足底腱膜近端与跟骨相连,远端与趾骨近端相连,足舟骨的侧面高度能够反映纵向足弓高度;纵向足弓在功能上可以加强足中部区域的联结,提高了足的承载力和稳定性,为人类双足直立运动提供一个稳定的支撑和有效的推进杠杆。长期以来,进化生物学家对现代人类足部的这种解剖学特征一直很感兴趣。纵向足弓结构不仅可以区分人类和类人猿的足,而且还可以确定足的功能是适用于双足直立运动还是树栖运动。人类在双足直立奔跑的支撑阶段(见图3),跖趾关节背屈,足底腱膜包裹着跖骨的近端,产生拉力,这种线性力将跟骨向前拉,有效地抬起纵弓,这使得足部成为从踝关节有效传输能量的杠杆;另外,足部固有的肌肉加强了足弓刚性支撑,使足关节产生动力,从而使踝关节产生蹬力。而黑猩猩跑步支撑阶段,足部如同“扁平足”,基本上与地面保持接触,没有形成拱形结构,不利于承受跑步过程的冲击力。相比之下,人类拥有像弹簧一样的纵向足弓,能够承受快速奔跑过程产生的高强度冲击力,并且在竖向冲击载荷作用下产生水平推力,更有利于人类奔跑。人类足弓结构性的适应性变化或许也是人类长期双足直立运动尤其是长距离耐力跑步运动造成的结果[36]。
2)人类耐力跑步假说的跟骨长度适应性变化原因。
从生物力学的视角,力矩臂的长度在确定肌腱能量储存方面起着重要作用。在力矩一定的前提下,较小的力矩臂可以引起肌腱能量的增加,进而有利于人类双足直立运动的经济性。人类在跑步过程中,作用于跟骨的跟腱表现出弹簧的特征,研究发现与黑猩猩比较,人类的跟腱明显较长,从而提高了肌肉肌腱的弹性能量储存[15]。在人类跑步支撑阶段,如果作用于跟骨的跟腱产生的肌力矩足够大,就会产生良好的缓冲和蹬伸效果。虽然不能从化石中确定肌腱长度,但是,基于跟骨长度也可以间接反映出运动的经济性。由于作用于跟骨的肌力矩主要取决于跟腱作用力以及作用于跟骨的力矩臂(跟腱作用线到踝关节中心的垂直距离),因此,如果跟骨长度越短,那么跟腱力臂则可能越短,在肌力矩一定的前提下,就能更加节省机械能量消耗,运动经济性则会更好。跑步经济性主要是指在稳定状态下跑步,摄氧量达到稳定状态时每单位体重和每公里的摄氧量。有研究发现,人类跟骨长度与双足直立跑步经济性呈现负相关,而与双足直立行走经济性无关,说明跟骨的长度越长,跑步经济性越差[37]。研究人员进一步比较了现代人和尼安德特人(距今12万年前生活在欧洲及西亚的古人类)的跟骨长度,发现尼安德特人与现代人类相比跟骨结节相对较长,这会增加他们的跑步能量消耗;现代人则表现出更有利于耐力跑步的生物力学优势,例如,现代人的跟骨结节长度较短,跟腱的力矩臂也会较短,跟腱就会存储和释放更大的弹性能量,从而降低能量消耗,提高跑步经济性,这里所涉及的跟骨长度主要是指跟骨结节后边缘与距骨结节边缘的最大距离。尽管这种测量不是精确的肌腱力臂,但是,在古人类化石缺少肌肉肌腱的前提下,间接通过跟骨长度也为跑步的生物力学的动力学研究提供了机会。通过和尼安德特人的比较,也从侧面说明人类在进化过程中跟骨结节发生了结构性变化,使人类更适合耐力跑步。
3)人类耐力跑步假说的髋关节适应性变化原因。
髋关节连接着躯干和下肢,由股骨和骨盆构成,它的周围环绕着人体最强大的肌肉和韧带,适合人类耐力跑步相关的髋关节适应性变化主要体现在骨盆、股骨及周圍的肌肉结构。
(1)现代人类髂骨窄、股骨颈短,使耐力跑步更加节省能量。关节适应性变化主要体现在髋关节在人类演化过程开始逐渐变窄,从生物力学角度对人类跑步具有积极意义。通过对出土南方古猿化石分析发现[38],南方古猿髋关节的髂骨翼相比现代人更加宽大,现代人较窄的髋关节更适合陆地双足直立运动。也有学者认为,与非人类灵长类动物相比,人类骨盆的基本改变促进了人类两足直立运动[39]。通过比较发现,南方古猿的髂骨宽大、股骨颈长,意味着臀大肌力臂较大,进而臀大肌对抗体重的力矩增大,导致关节反作用力产生更大的股骨轴弯矩。另外,骨盆使髋关节能够产生较大的伸肌力矩,所以猿类具有较强的攀爬能力。但是,由于髋关节伸展的范围有限,所以导致南方古猿行进的时候表现出蹲伏的姿态,运动效率受到一定的影响(见图4)[40]。现代人的股骨颈相比南方古猿较短,下肢较长,臀大肌对抗体重的力矩减小,相对于髋关节的弯矩也变小,关节反作用力产生较小的股骨轴弯矩,人类的骨盆使髋关节伸肌力矩降低,髋关节伸展范围增大,从而大大改善了人类运动效率,这对人类耐力跑步具有积极意义。
(2)人类髋关节伸展幅度的改善降低了耐力跑步的能量消耗。通过比较黑猩猩和人类的步态生物力学分析发现,除了解剖学结构,步态的生物力学差异可以很好地解释双足和四足行进以及黑猩猩和人类之间能量消耗的差异,人类运动的能量消耗降低与髋关节伸展幅度增大相关,黑猩猩双足和四足运动的机械能耗差异不显著,人类运动的机械能耗比黑猩猩四足和双足运动的能量消耗均低75%,他们将人类双足直立行进的机械能耗节省化归因于人类更窄的髋关节和更长的下肢[41]。研究人员通过对原始人类化石分析以及比较人类和黑猩猩的生物力学特征表明,人类双足直立运动确实比四足运动的能量消耗更低,双足运动的经济性为人类擅长双足耐力跑步奠定了基础,于是指出习惯性的双足直立运动行为可能更有利于改变髋关节结构以允许身体表现出更经济的跑步姿势。
(3)南方古猿髋关节伸展幅度以及较长的股骨形态(见图2),表明原始人类具有适合耐力跑步的下肢形态基础,具有能量节省化的下肢形态学解剖优势。虽然南方古猿的膝关节和髋关节屈曲幅度相比现代人更大,但是有学者认为,南方古猿双膝存在弯曲可能会进一步增加髋关节的伸展,进而节省能量消耗,使南方古猿在直立运动过程表现出较好的经济性[42]。另外,有学者发现,与现存猿类相比南方古猿的股骨结构更接近现代人,这说明南方古猿在双足直立运动过程中,其髋关节运动学与人类相似;研究结果还表明,南方古猿股骨头小梁更加强健,这表明与现存猿类相比,其肌肉骨骼系统的整体负荷增加了,下肢能够承受更大的冲击载荷,这些结果同时也为原始人类双足直立运动提供了新的证据,人类骨盆及髋关节结构的适应性变化,奠定了人类适合耐力跑步的生物学基础[43]。
4)人类耐力跑步假说的下肢长度适应性变化原因。
现代人类下肢长度要明显长于大猩猩,或许可以改善了人类耐力跑步的效率。但是迄今为止,现代人类下肢长度的增加是否会改善运动效率仍然是争论的焦点。人类在进化过程中下肢长度逐渐增加,已确定直立人(距今200万年前)的下肢长度明显长于南方古猿(距今360万年前)[44]。效率反应了生物能转换为机械能的多少,一种观点认为,下肢长度与运动效率没有关系,例如,研究人员测量了博物馆标本中的古人类下肢长度发现[45],当去除体重因素时,下肢长度和运动能量消耗没有显著关系;一种观点认为,人类下肢长度增加会提高人体运动效率,降低人类在运动过程的机械能量消耗,进而使人类运动的距离更远,有助于人类耐力能力的塑造[46];然而,也有观点认为,下肢长度增加降低了人类的运动效率。例如,根据南方古猿化石通过生物力学分析发现[47-48],南方古猿下肢长度较短,机械做功效率实际上更有利,据此认为,南方古猿比现代人类具有更好的机械运动效率,因为南方古猿相比现代人类下肢长度更短,转动惯量就越小,能量消耗就越低。但是,南方古猿拥有较短下肢,他们只能在慢速下运动效率表现更好,更适合在食物资源相对充足的环境中慢速运动,这恰恰限制了他们的日常运动范围和距离,不利于发展耐力;而近代人类的日常运动范围要比南方古猿大20%以上,在食物资源更加分散的环境中,近代人类需要更高的步行速度和更长的运动距离,这在一定程度上促进了现代人类的耐力跑步水平。实际上,影响效率的因素有多种,机械功也不是整个能量利用的主要决定因素[49],因此,需要同时考虑人类耐力跑步的生物能量学特征。一般认为,下肢较长可以改善人类奔跑的效率。
3 生活方式演变是人类耐力跑步假说成立的外在原因
生活方式演变主要体现在人类饮食结构的变化,这种变化与自然环境的改变密不可分。在中新世晚期(约600万年前),非洲地区的环境显得更温暖和更潮湿,原始人类和黑猩猩的共同祖先们都生活在这种雨林环境中,主要依靠树上的果实为生。但是,由于全球和地区气候的变化,非洲气候总体上开始变的寒冷和干燥,这些气候变化进一步造成雨林生态环境的变化,雨林变的更加分散和开阔[50]。这个时期,对于主要以水果为食的原始人类来说,更开阔的栖息地对生存造成一定的压力,因为其赖以生存的水果开始变得更加分散和更有季节性。于是,原始人类的觅食行为方式开始发生改变,比如,除了采集树上的食物之外,腐肉或捕食猎物成为另一种能量来源。这种选择性的生态环境压力不但迫使原始人类获取食物的多样化,也迫使原始人类的活动空间更加广阔。于是,在捕食猎物的过程中,觅食空间的增大也逐渐塑造了原始人类独特的耐力跑步能力。
3.1 适合人类耐力跑步的饮食结构改变
肉食的出现以及肉类作为饮食的重要比例开始增加是人类进化的重要体现。原始人类一般通过采集获得食物,除了爬树采摘果实以及挖掘根茎之外,在外界自然环境选择的压力之下,原始人类也开始通过长途跋涉进行狩猎以获取肉食,肉食切割石材工具的出现和一些原始社会遗址的屠宰证据支持了这一观点。研究发现,德马尼西人在大约180万年前居住在南高加索的北部温带栖息地[51]。这是在非洲以外闻名的最早原始人类,从出土的石器发现,德马尼西人已经可以狩猎并用石器切割肉[41]。学者们研究了德马尼西人后肢的形态学,以验证饮食结构变化尤其是狩猎使人类耐力跑步能力增加的假设。通过比对现代人、黑猩猩以及其他原始人类化石数据发现,德马尼西人后肢功能与现代人相似。比如,具有纵向的足弓、增加的下肢长度以及踝关节形态等,均与现代人相似;足部(特别是跖骨)以及胫骨形态等均与原始人类相似[41]。这些结果支持原始人类的狩猎可以改善耐力跑步表现的假设。
3.2 适合人类耐力跑步的持久性狩猎方式的出现
耐力狩猎(Endurance Hunting)是一种狩猎技术,通过持久性的跑步、行走和追踪等方式组合来追踪猎物,直到獵物体力耗尽,然后将其捕获。相对于完全使用工具狩猎,耐力狩猎历史更加悠久。研究发现,人类直立行走的时间大约在360万年前,而人类在180万年前才开始使用石器工具[52],使用工具的时间要落后于开始双足直立行走的时间。因此,至少180万年以前的人类祖先还不能够使用工具狩猎,只能依靠长距离的耐力跑步或行走的方式追赶捕获猎物。
研究认为,早期的人类是通过耐力跑步方式来捕获猎物,持久性的耐力狩猎通常在一天中最热的时候,人类在体温调节方面的优势可以使人类降低能量消耗,进而在炎热的天气能够以更经济的长距离跑步进行狩猎[53-54]。而动物可能需要喘气才能降温,这也意味着它们必须减速来缓和核心温度的增高,这样就为人类耐力狩猎提供了机会。人类通过自身的体温调节能力,在炎热的天气首先通过长距离慢速奔跑持续驱赶猎物,使猎物体力消耗殆尽,然后在近距离安全地捕获那些筋疲力尽的猎物,这种长距离跑步狩猎的方式对人类耐力跑步能力的提升具有重要意义。另外,研究发现非洲直立人较长的下肢和纤细的身体构造可能更适合在炎热的气候中维持较高的活动水平狩猎,修长的人体形态结构增加了皮肤表面积与体重的比例,进而可以促进体内热量与环境对流,减少热应激伴随的能量需求[41]。人类身体的散热优势为耐力狩猎提供了机会,也逐渐塑造了人类的耐力跑步能力。
人类体温调节优势让人类更适应在热环境下进行耐力狩猎。如果代谢热产生的速度比消除的速度快,那么体温就会上升,进而限制奔跑距离。大多数哺乳动物在奔跑时都依靠蒸发冷却来维持体温平衡,蒸发冷却主要通过两种独立的机制来实现,一种机制是通过鼻粘膜、口腔以及舌头表面的呼吸蒸发散热(比如,喘气),另一种机制是通过体表的汗液蒸发散热,而人类更多的是依靠汗液蒸发散热[54]。与其他动物相比,人类的汗腺可能比其他动物的汗腺受到更高水平的神经控制,因而单位面积出汗量具有优势,进而散热能力较好。此外,人类汗腺发育良好,人体毛发也相对较少,稀少的毛发会具有更大的导热系数,跑步时散热的速度大大加快,人类主要通过出汗的调节体温机制可能比其他哺乳动物更有效地消除跑步过程中产生的代谢热[54]。相比大多数哺乳动物只能在相对凉爽的条件下行走或跑步,人类在热环境中进行狩猎具有独特的耐力优势。
据报道,近代以来许多部落仍然保留着这种耐力狩猎的方式[14]。比如,生活在非洲南部地区的一个原始狩猎-采集民族——布须曼人(Bushmen),他们会在雨季猎杀杜鹃和大羚羊等,在炎热的旱季猎杀角马和斑马。生活在墨西哥北部的塔拉乌马拉印第安人(Tarahumara Indians),能够在墨西哥北部的群山中追逐鹿,直到这些动物精疲力竭而倒下,然后用手捕获它们。美国西南部的帕尤特人(Paiutes)和纳瓦霍人(Navajo)用耐力狩猎的方法猎杀叉角羚,这种羚羊是所有哺乳动物中速度最快的一种。此外,澳大利亚西北部的土著居民以耐力狩猎的方式成功地捕猎袋鼠而闻名。虽然被捕食动物的耐力都很出色,但是,都败在了人类的耐力之下。人类通过双足直立的方式进行持久性耐力狩猎的出现,从侧面反映了人类的耐力跑步能力或许是经历了上百万年进化后,人类为了生存而对自然界产生的一种适应。
3.3 适合人类耐力跑步的觅食距离的增加
从南方古猿到人类的过渡中,人类的觅食行为发生变化,日常运动距离开始增加。研究发现,食肉动物每天运动的距离大约是食草动物的4倍,并且较长的运动距离可以提升耐力水平,与现存哺乳动物的比较研究表明,食肉动物在狩猎过程运动量显著增加,该研究也进一步证实原始人类饮食结构的变化有助于耐力跑步能力的提升,为了获得肉食而增加的狩猎距离也为人类天生的耐力跑步优势提供了间接证据[55]。
环境因素是人类觅食距离增加的重要影响因素,由于寒冷的温度和干燥的气候环境使雨林分布更分散,增加了原始人类觅食的距离[56-57。如果说原始人类需要移动更长的距离才能找到食物(比如,分散的果实或者肉食)的话,那么选择双足直立运动确实节省了自身的能量消耗,提高了运动效率[27]。相比人类而言,黑猩猩75%以上的食物主要依赖于水果,平均每天只走2~3 km[58]。在此情况下,黑猩猩依靠四肢运动过程中,肢体姿势处于一种弯曲状态,其运动过程每单位体重每单位距离消耗的能量是人类双足运动的4倍[59]。在觅食距离增加的情况下,人类双足直立运动会获得巨大的能量收益。因此,人类双足运动的经济性优势为原始人类提供了明显的进化优势,人类可以在不增加日常能量消耗的前提下运动更远的距离[27,60],进而通过双足直立运动从森林转移到草原去寻找更丰富的食物[27,60-61],或节省更多的能量用于增加繁殖、延长寿命和优化大脑等[62]。
3.4 关于人类耐力狩猎方式的不同观点
通常认为,耐力跑步是原始社会人类耐力狩猎的一种方式。也有学者持怀疑态度,认为通过耐力跑步持续狩猎在现代人类中非常罕见[15],尤其在非常开放和非常热的栖息地更是稀少等。有研究者观察到现代人可以通过耐力跑步进行狩猎,虽然成功率比较低。例如,有学者曾经对非洲南部的卡拉哈里沙漠中部地区的现代人耐力跑步狩猎进行了观察,在20年的观察过程中,观察到两次耐力跑步狩猎是自发组织且成功的,这种耐力跑步狩猎的成功率显的比较低[63]。还有学者认为,人类主要是通过耐力行走狩猎,而非耐力跑步狩猎,人体的构造是为行走而非跑步设计的[64]。原始人类在狩猎的过程中需要放缓脚步顺着动物的节奏前进,同时还要时刻注意脚下的危险,一方面不能让脚下发出较大的声音,另一方面还要小心翼翼地避开不平坦的路面,直到足够接近猎物才能将其捕获。在实际的狩猎过程中,原始人采用的是缓慢行走的狩猎方式,而不是持续奔跑。因为对于原始人类来讲,奔跑具有诸多危险,很有可能踩在洼地或者磕绊石块进而发生严重的脚踝扭伤,这种小事故对于原始人类来说却是一个灾难性的事故。由此看来,作为人类耐力狩猎的一种方式,跑步对原始人类还是非常危险的,即使是现代人也会感受到在不平坦的路面奔跑的风险。迄今为止,由于缺少充足的证据,耐力狩猎在学术界依然存在争论。
综上所述,从体质人类学的视角探索人类耐力跑步的渊源,一方面为人类双足直立运动寻找一种解释,另一方面也从体质人类学的角度去阐释人类或许天生就拥有良好的耐力跑步能力。通过文献回顾发現,人类其实天生拥有着良好的耐力基础,这种耐力跑步能力是200万年以来原始人类演化的结果。人类自身的解剖结构、体温调节以及饮食结构等因素决定了人类天生具有出色的耐力跑步能力,现代社会的马拉松跑步运动具有深刻的进化根源。虽然现代人类已经不再将耐力跑步作为谋生的手段,但是现代人类双足运动的经济性特征以及所具有的耐力跑步能力是人类固有特征的现代体现。
当前,随着经济社会的不断发展,科技在不断进步,人类身体活动也面临着分化,久坐不动的生活方式开始出现,人类的体力活动在不断减少,身体素质在不断下降,运动能力在不断退化,各种现代文明病不断出现,已经威胁到了人类的生存、健康与发展。面对疾病、面对压力,人类并没有完全就此屈服,一部分人通过主动积极的耐力跑步来提高生命的质量,风靡全球的马拉松热现象也从一个侧面折射出了人类面对现代文明病威胁的一种本能抗争。从体质人类学的视角,现代社会的耐力跑步热现象有其形成的内在原因,或许是在人类起源以来的漫长历程中人类为了生存与发展自然选择的结果。
在人类进化的过程中,人类自身形态结构的适应性变化是耐力跑步假说成立的内在原因;人类生活方式的演变及对环境的适应是耐力跑步假说成立的外在原因。耐力跑步的形态学基础主要体现在人类进化过程中逐渐形成了更适合跑步的后颅特征、下肢更长、脚趾指骨更短、足弓较高且弹性更好、髋关节变窄、臀大肌变得更加宽大等;耐力跑步的生活方式演变因素主要体现人类饮食结构的变化,肉食的出现以及肉类作为饮食的重要比例开始增加、持久性的耐力狩猎方式出现、人类耐力狩猎距离的增加以及双足直立运动经济性增加了人类耐力狩猎范围等。总而言之,现代社会的耐力跑步热现象有其形成的进化根源,人类耐力跑步可以追溯到人类起源与进化的整个过程,人类耐力跑步的形成是一个极其复杂的演化过程,受到人类遗传、生物变异以及自然选择等多种因素的共同影响。然而,耐力跑步对人类进化的解释只能称之为一种假说,未来关于耐力跑步假说仍然需要更多的研究进一步证实。
参考文献:
[1] WILLIAMS P T. Relationship of distance run per week to coronary heart disease risk factors in 8283 male runners. The National Runners' Health Study[J]. Arch Intern Med,1997,157(2):191-198.
[2] WILLIAMS P T. Walking and running produce similar reductions in cause-specific disease mortality in hypertensives[J]. Hypertension,2013,62(3):485-491.
[3] KALAK N,GERBER M,KIROV R,et al. Daily morning running for 3 weeks improved sleep and psychological functioning in healthy adolescents compared with controls[J]. J Adolesc Health,2012,51(6):615-622.
[4] 邢晓燕. 我国大型马拉松参赛者跑步训练参赛的动机内容与结构分析——基于跑步爱好者动机量表(MOMS)中文简版的信效度检验[J]. 体育成人教育学刊,2016,32(5):1-7.
[5] 刘转青,练碧贞. 自由视角下的“马拉松热”[J]. 北京体育大学学报,2017,40(8):34-39.
[6] 张实. 体质人类学[M]. 昆明:云南大学出版社,2003.
[7] 朱泓. 体质人类学[M]. 长春:吉林大学出版社,1993.
[8] 胡小明. 体育人类学进展[J]. 北京体育大学学报,2004,27(3):289-293.
[9] 胡小明. 体育人类学的兴起[J]. 体育学刊,1999,16(1):54-57.
[10] 吴汝康. 试论体质人类学的体系和内容[J]. 人类学学报,1985(3):286-287.
[11] THORPE S K, HOLDER R L, CROMPTON R H. Origin of human bipedalism as an adaptation for locomotion on flexible branches[J]. Science. 2007,316 (5829):1328-1331.
[12] WASHBURN S L. Tools and human evolution[J]. Scientific American. 1960,203(203):63.
[13] RODMAN P S. Foraging and social systems in Orangutans and chimpanzees[G]//RODMAN P S,CANT J G H. Adaptations for foraging in nonhuman primates:Contributions to an organismal biology of Prosimians,Monkeys,and Apes. New York:Columbia University Press,1984.
[14] CARRIER D R,KAPOOR A K,KIMURA T,et al. The energetic paradox of human running and hominid evolution[J]. Current Anthropology,1984,25(4):483-495.
[15] BRAMBLE D M,LIEBERMAN D E. Endurance running and the evolution of Homo[J]. Nature,2004,432(7015):345-352.
[16] 程大力,周西寬. 由英雄美女说起——论奥林匹克的人类生物共性和自然或原始基础[J]. 体育文化导刊. 2004(3):22-26.
[17] 乔玉成. 进化·退化:人类体质的演变及其成因分析——体质人类学视角[J]. 体育科学,2011,31(6):87-97.
[18] 耿业进,胡小明. 体育的生物学本源[J]. 体育与科学,2001,22(5):5-6.
[19] 刘欣然,乐严严. 基于人类身体本能的体育运动哲学解读[J]. 体育学刊,2009,16(12):27-31.
[20] 李力研. 人类种族与体育运动[J]. 中国体育科技,2001,37(6):4-9.
[21] BARTHOLOMEW G A, BIRDSELL J B. Ecology and the Protohominids[J]. American Anthropologist,1953,55(4):481-498.
[22] TAYLOR C R,SCHMIDT-NIELSEN K,RAAB J L. Scaling of energetic cost of running to body size in mammals[J]. Am J Physiol,1970,219(4):1104-1107.
[23] BALKE B,SNOW C. Anthropological and physiological observations on Tarahumara endurance runners[J]. Am J Phys Anthropol,1965,23(3):293-301.
[24] GROOM D. Cardiovascular observations on Tarahumara Indian runners-the modern Spartans[J]. Am Heart J,1971,81(3):304-314.
[25] DEVINE J. The versatility of human locomotion[J]. American Anthropologist,1985,87(3):550-570.
[26] SINCLAIR A R,LEAKEY M D,NORTON- GRIFFITHS M. Migration and hominid bipedalism[J]. Nature,1986,324(6095):307-308.
[27] RODMAN P S,MCHENRY H M. Bioenergetics and the origin of hominid bipedalism[J]. Am J Phys Anthropol,1980,52(1):103-106.
[28] HUNT K D. Mechanical implications of chimpanzee positional behavior[J]. Am J Phys Anthropol,1991,86(4):521-536.
[29] ISBELL L A,PRUETZ J D,LEWIS M,et al. Locomotor activity differences between sympatric patas monkeys (Erythrocebus patas) and vervet monkeys (Cercopithecus aethiops): Implications for the evolution of long hindlimb length in Homo[J]. Am J Phys Anthropol,1998,105(2):199-207.
[30] ROSE M D. A hominine hip bone,KNM-ER 3228,from East Lake Turkana,Kenya[J]. Am J Phys Anthropol,1984,63(4):371-378.
[31] JOHANSON D C,WHITE T D. A systematic assessment of early African hominids[J]. Science,1979,203(4378):321-330.
[32] LIEBERMAN D E,RAICHLEN D A,PONTZER H,et al. The human gluteus maximus and its role in running[J]. J Exp Biol,2006,209(Pt 11):2143-2155.
[33] ALEMSEGED Z,SPOOR F,KIMBEL W H,et al. A juvenile early hominin skeleton from Dikika,Ethiopia[J]. Nature,2006,443(7109):296-301.
[34] JONES S,MARTIN R D,PILBEAM D R. The Cambridge encyclopedia of human evolution[M]. Cambridge [England]; New York,NY,USA:Cambridge University Press,1992.
[35] HOLOWKA N B,O'NEILL M C,THOMPSON N E,et al. Chimpanzee and human midfoot motion during bipedal walking and the evolution of the longitudinal arch of the foot[J]. J Hum Evol,2017,104:23-31.
[36] HOLOWKA N B,LIEBERMAN D E. Rethinking the evolution of the human foot:Insights from experimental research[J]. J Exp Biol,2018,221(Pt 17).
[37] RAICHLEN D A,ARMSTRONG H,LIEBERMAN D E. Calcaneus length determines running economy: Implications for endurance running performance in modern humans and Neandertals[J]. J Hum Evol,2011,60(3):299-308.
[38] STERN J J,SUSMAN R L. The locomotor anatomy of Australopithecus afarensis[J]. Am J Phys Anthropol,1983,60(3):279-317.
[39] GRUSS L T,SCHMITT D. The evolution of the human pelvis:Changing adaptations to bipedalism,obstetrics and thermoregulation[J]. Philos Trans R Soc Lond B Biol Sci,2015,370(1663):20140063.
[40] RICHMOND B G,JUNGERS W L. Orrorin tugenensis femoral morphology and the evolution of hominin bipedalism[J]. Science,2008,319(5870):1662-1665.
[41] PONTZER H,ROLIAN C,RIGHTMIRE G P,et al. Locomotor anatomy and biomechanics of the Dmanisi hominins[J]. J Hum Evol,2010,58(6):492-504.
[42] SOCKOL M D,RAICHLEN D A,PONTZER H. Chimpanzee locomotor energetics and the origin of human bipedalism[J]. Proc Natl Acad Sci U S A,2007,104(30):12265-12269.
[43] RYAN T M,CARLSON K J,GORDON A D,et al. Human-like hip joint loading in Australopithecus africanus and Paranthropus robustus[J]. J Hum Evol,2018,121:12-24.
[44] JUNGERS W L. Lucy's limbs:Skeletal allometry and locomotion in Australopithecus afarensis[J]. Nature,1982,297(5868):676-678.
[45] STEUDEL K,BEATTIE J. Does limb length predict the relative energetic cost of locomotion in mammals?[J]. J. Zool. (Lond.),1995,235(3):501-514.
[46] STEUDEL-NUMBERS K L,TILKENS M J. The effect of lower limb length on the energetic cost of locomotion:Implications for fossil hominins[J]. J Hum Evol. 2004,47(1-2):95-109.
[47] KRAMER P A. Modelling the locomotor energetics of extinct hominids[J]. J Exp Biol,1999,202(Pt 20):2807-2818.
[48] KRAMER P A,ECK G G. Locomotor energetics and leg length in hominid bipedality[J]. J Hum Evol,2000,38(5):651-666.
[49] 孫明运,孙泊,章翔,等. 能量平衡模型结合运动生物力学手段测试人体运动做功效率[J]. 中国组织工程研究. 2013(11):2032-2039.
[50] CERLING T E,WYNN J G,ANDANJE S A,et al. Woody cover and hominin environments in the past 6 million years[J]. Nature,2011,476(7358):51-56.
[51] LORDKIPANIDZE D,JASHASHVILI T,VEKUA A,et al. Postcranial evidence from early Homo from Dmanisi,Georgia[J]. Nature,2007,449(7160):305-310.
[52] ISAAC G. The food-sharing behavior of protohuman hominids[J]. Sci Am,1978,238(4):90-108.
[53] LIEBERMAN D E,BRAMBLE D M. The evolution of marathon running:Capabilities in humans[J]. Sports Med,2007,37(4-5):288-290.
[54] LIEBERMAN D E. Human locomotion and heat loss:An evolutionary perspective[J]. Compr Physiol,2015,5(1):99-117.
[55] CARBONE C,COWLISHAW G,ISAAC N J,et al. How far do animals go? Determinants of day range in mammals[J]. Am Nat,2005,165(2):290-297.
[56] CERLING T E,HARRIS J M,MACFADDEN B J,et al. Global vegetation change through the Miocene/ Pliocene boundary[J]. Nature,1997,389(6647):153-158.
[57] CERLING T E,EHLERINGER J R,HARRIS J M. Carbon dioxide starvation,the development of C4 ecosystems,and mammalian evolution[J]. Philos Trans R Soc Lond B Biol Sci,1998,353(1365):159-170,170-171.
[58] PONTZER H,WRANGHAM R W. Ontogeny of ranging in wild chimpanzees[J]. International Journal of Primatology,2006,27(1):295.
[59] PONTZER H,RAICHLEN D A,RODMAN P S. Bipedal and quadrupedal locomotion in chimpanzees[J]. J Hum Evol,2014,66:64-82.
[60] ISBELL L A,YOUNG T P. The evolution of bipedalism in hominids and reduced group size in chimpanzees:Alternative responses to decreasing resource availability[J]. J Hum Evol,1996,30(5):389-397.
[61] WHITE T D,ASFAW B,BEYENE Y,et al. Ardipithecus ramidus and the paleobiology of early hominids[J]. Science,2009,326(5949):75-86.
[62] PONTZER H,BROWN M H,RAICHLEN D A,et al. Metabolic acceleration and the evolution of human brain size and life history[J]. Nature,2016,533(7603):390-392.
[63] LIEBENBERG L. The relevance of persistence hunting to human evolution[J]. J Hum Evol,2008, 55(6):1156-1159.
[64] PETER A. Why running is not for people[J]. BMJ,2003,327(7429):1476.
