氢氧同位素示踪法探测新疆地区防护林和棉花体系水分来源与竞争
2019-11-07蔡东旭兰志龙雷加强
郭 辉,赵 英,蔡东旭,兰志龙,雷加强
1 中国科学院新疆生态与地理研究所,乌鲁木齐 830011 2 西北农林科技大学资源环境学院, 杨凌 712100 3 中国科学院大学,北京 100049
绿洲作为新疆干旱区人类活动的重要载体,生态环境脆弱,使内部各个要素保持健康和谐共存极其重要。由于防护林对沙漠绿洲区具有防风固沙效应[1, 2],因此在新疆绿洲区形成了大面积农田-防护林纵横交错的景观格局。但研究表明防护林根系在趋水趋肥作用下大量分布于农田一侧,通过吸收农作物养分水分,限制农作物生长与产量提高[3, 4]。水分是推动绿洲生态系统各要素健康发展的关键因子[5],平衡农作物与防护林的水分竞争可以提高农作物产量与防护林正常生长。因此通过研究农作物和防护林水分来源及根系分布,量化两者的水分竞争区域,对农林复合系统的和合理配置与管理具有重要指导意义。
传统上采用挖掘法或根钻法,研究植物根系分布,来推断植物水分利用来源[6],且费时费力;其次,根菌的分布也可辅助植物对水分养分利用[7, 8];此外,植物根系活性随着土壤含水量的变化而异,预示着某一层大量根系并不能准确反映同等比例的水分吸收量[9]。稳定同位素示踪技术具有较高的灵敏性和准确性,是确定植物水分来源的重要工具[10]。De Wispelaere等[11]研究表明热带半干旱区植物水分来源随季节而变化,雨季主要利用降雨,旱季利用地下水;Dai等[12]利用氢氧稳定同位素在温带干旱生态系统中发现梭梭等荒漠植物主要利用深层地下水。然而,氢氧稳定同位素并不能准确得出植物根系分布。近年来,人工添加稳定同位素探测植物根系分布和资源利用的范围也逐步得到应用,Beyer等[13]通过添加δD同位素准确地判断出非洲半干旱区域不同植物的根系空间分布,表明灌木的根系深度一般不超过1 m,乔木则可以吸收4 m以下的水分。目前氢氧同位素技术主要集中在野外自然条件下,而在人工植被措施下很少涉及[14]。Wang等[15]对华北地区棉花与玉米水分来源研究发现两者在不同的生长季吸收不同深度土壤水分。
近年来,新疆各地出现大面积防护林死亡现象,部分原因是农民担心防护林对农作物的负面影响,而对防护林减少灌水和病虫害防治所致[16];然而,防护林的死亡将会导致绿洲遭受更加严重风沙侵蚀[17]。面对这一矛盾,本文选择新疆种植面积最大的棉花与主要防护林树种杨树作为研究对象,通过示踪δD同位素及测定不同生长期棉花、杨树及其土壤中的δD与δ18O值,量化棉花与杨树的主要水分来源,分析潜在竞争区域及防护林对农作物水分负面影响程度。研究结果对新疆绿洲地区防护林的合理管护,及绿洲内部生态平衡的建设具有重要的指导意义。
1 材料和方法
1.1 研究区概况
图1 研究区地理位置图Fig.1 Location of the study area
研究区位于古尔班通古特沙漠西南缘的莫索湾盆地绿洲,地理坐标东经86°06′—86°50′,北纬44°60′—45°50′,平均海拔358.8 m,属于典型的大陆干旱荒漠气候(图1)。土壤以灰漠土和沙壤土为主[18]。年平均气温约为4—6℃,夏季天气酷热,蒸发强烈,7月平均温度最高,达22—26℃,气温年较差在42—44℃之间,≥10℃的活动积温为3100—3200 h。年降水量少于120 mm,季节分配较为均匀,但仅有少量能被农作物利用,而天山的积雪融水为农业生产提供了丰富的水源。
1.2 样地选择
试验于2016年7月到9月在莫索湾试验站棉花试验田进行,如图2。2016年7月7号,样地内选择10棵冠幅相似,间距大于20 m,干径约为35 cm的杨树。首先采用土钻在4棵杨树附近依次钻50、100、200、400 cm的洞,每个洞3个重复;同时以20 cm为间隔采集土壤剖面样品(即0—20、20—40、40—60、60—80,80—100 cm等),每层3个重复,将每个样方一部分土壤装入50 mL塑料瓶中,并用封口膜封口,迅速冷冻保存,用于抽提水,另一部分装入铝盒中,用烘干法测土壤含水量;其后选择杨树长势良好枝条,除去外皮和韧皮部迅速装入8 mL玻璃瓶中,用封口膜封口冷冻;最后依次将300 mL水且含量为33% δD小心装入塑料袋放入每个杨树洞底,扎破,迅速将土填充,并在接下来6 d每天采集杨树木质部,冷冻封存。接着在分别距杨树2、5、8 m棉田中,每个距离用土钻分别打50、100 cm深度,每个深度间隔大于10 m(3个重复),依次采集土样棉花样本与棉花正对面杨树样本冷冻,然后进行δD同位素示踪与填土,方法与上同,并在接下来6 d每天采集样点正对面杨树样本。当所有样品采集完成之后,在试验第7天, 再次用土钻取杨树100 cm与200 cm深处示踪点的土壤,每个样点一钻,按照之前方法采土样并冷藏。
最后,2016年8月16日再次分别采集杨树和距离杨树8 m远处棉田200 cm、100 cm深的土样,为防止之前的人工填充土干扰,避开之前的采土样点,并且采集植物样本。
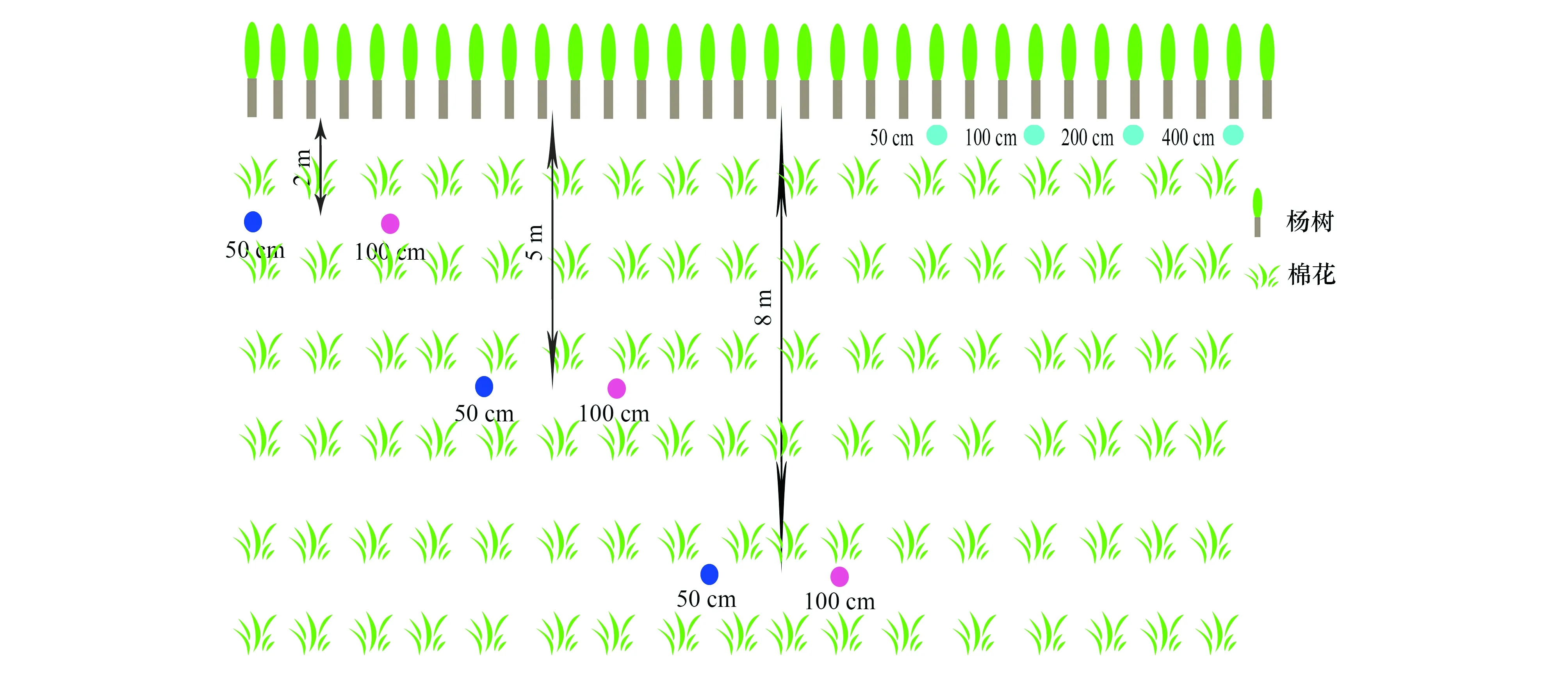
图2 试验设计图Fig.2 Experimental design in farmland of Mosuowan oasis不同颜色圆点表示δD同位素示踪不同的深度
1.3 水分提取与同位素测定
采用低温真空蒸馏法(LI- 2000全自动真空冷凝抽提系统进行水分抽提装置)抽提植物茎干和土壤样品水分,抽提水放入玻璃样品瓶中冷藏,用DLT- 100液态水同位素仪(Lios Gatos Research, Mountain View, USA)测定δD和δ18O值。其中δD的误差不超过±2‰,δ18O为0.25‰。
1.4 数据分析方法
除一些盐生植物外,水分在由植物根系向茎干传输送的过程中氢氧同位素一般未因蒸发或新陈代谢导致分馏,因此木质部中或根茎结合部可以反映植物水分来源[19]。为了避免人工添加的δD同位素在后期实验中可能存在的潜在污染,本文采用δ18O同位素来判断植物主要水分来源,水分来源判定采取两种方法。其一是焦点法,植物样本的稳定同位素值与土样值相等,即可视为其主要水分来源区间;其二是SIAR混合模型(Stable Isotope Analysis in R, SIAR package, version 3.4.2, Parnell等[20]),该方法采用贝叶斯混合运算,考虑到了可能发生的同位素分馏与样本同位素值的多变性与多重来源的非一致性等,已成为当前主要的同位素来源分析方法。
本研究中,分别采用距地表深100 cm且距离杨树8 m棉花土样与200 cm深杨树土样分析水分来源区域。并且根据棉花不同生育期的生理表现,判定7月7日采集为蕾期棉花(棉花开始开花),8月16日为花铃期(棉花将要吐絮)。棉花与杨树根系吸水的潜在来源被认为是不同深度的土壤水。依据前人对棉花与杨树根系分布规律的研究结论[15],可以将棉田土壤划分为0—40 cm 土壤表层,40—100 cm土壤深层;杨树土壤划分为0—40 cm土壤表层,40—120 cm土壤较深层,以及120—200 cm 土壤深层。
1.5 数据分析
利用R语言进行数据分析,用origin 8.0软件绘图。
2 结果分析
2.1 土壤含水量变化
图3表示棉田与杨树土壤含水量随时间和深度的变化。在棉花蕾期(7月7)与花铃期(8月16),棉田各层土壤含水量未呈现出剧烈变化,平均土壤含水量约15%,这主要是棉田在滴灌条件下,灌溉频繁(平均7 d灌溉一次),土壤含水量一直保持在较高水平;又因土样采集时间不一致,分别为灌溉前一天和后一天,导致各层土壤含水量出现较大波动。杨树各土层含水量出现剧烈波动,表现出先增大后减小趋势。首先7月7日杨树未漫灌之前,表层0—60 cm土层含水量较低,60—180 cm土层土壤含水量变化保持相对恒定的值(约14%),与棉田含水量较为一致;在200 cm深处,土壤质地变粗,土壤持水能力较小,导致土壤含水量较小。在8月16日,杨树大水漫灌一周后,土壤含水量在0—100 cm土层显著高于灌溉水之前(P<0.01),而在100 cm以下含水量波动相对较小,表明灌溉一周后,水分入渗土壤剖面锋值约在100 cm深处。

图3 不同生长季棉田与杨树土壤质量含水量(a为棉田土壤含水量,b为杨树土壤含水量)Fig.3 Mean soil moisture content (%) on July 7th and August 16th in the soil profile at (a) cotton and (b) poplar plots
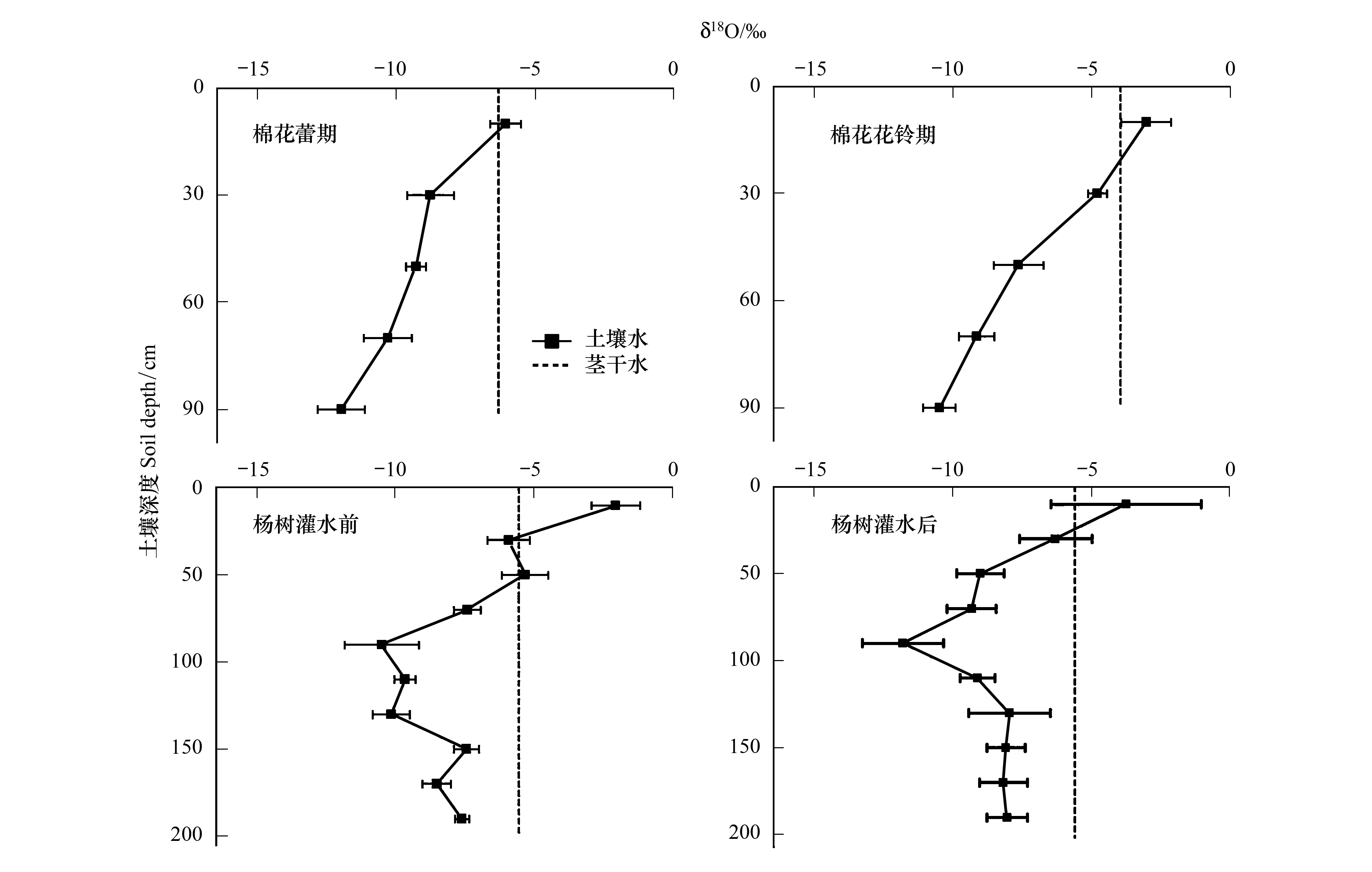
图4 棉花与杨树不同生长时期不同深度土壤水与茎干水的δ18O值Fig.4 δ18O values in soil water, cotton and poplar xylem at each growth stage
2.2 水分来源与贡献率
土壤水中氢氧稳定同位素组成主要受灌溉水(降水)、蒸发、地下水等多因素影响[21]。
图4为不同时期棉田与杨树土壤剖面δ18O的垂直变化,从图可知,δ18O值随着土壤深度增加而在减小,这是因为土壤表层强烈的蒸发作用,导致土壤水发生分馏,δ18O值变大。通过利用焦点法,得出棉花在蕾期,主要的水分来源于0—20 cm土层(图4);而在花铃期,水分主要来源于20—40 cm(图4)。进一步运用SIAR模型,量化各土层水分贡献率(图5),可知棉花在蕾期与花铃期主要吸收表层土壤水分(0—40 cm),贡献率高达94%和86%。
杨树作为新疆防护林主要树种,探测其主要水分来源是否来自农作物表层根系分布区域,是判断两者水分竞争程度的重要依据[3]。图4展示了杨树在漫灌前后,土壤200 cm以上,δ18O值的变化与木质部δ18O值,通过对比发现在灌溉前杨树水分主要吸收20—60 cm土壤水分,灌溉后偏向于吸收0—40 cm范围内土壤水分,呈现出吸水区向上迁移趋势。通过进一步用SIAR模型分析,灌溉前土壤0—40,40—120,120—200 cm分别贡献为41%,30%,29%;灌溉后为43%,26%,31%。灌溉后比灌溉前土壤0—40 cm水分贡献略有提高,但土壤40 cm以下仍然是主要的供水区间,提供了约60%水分来源(图5)。
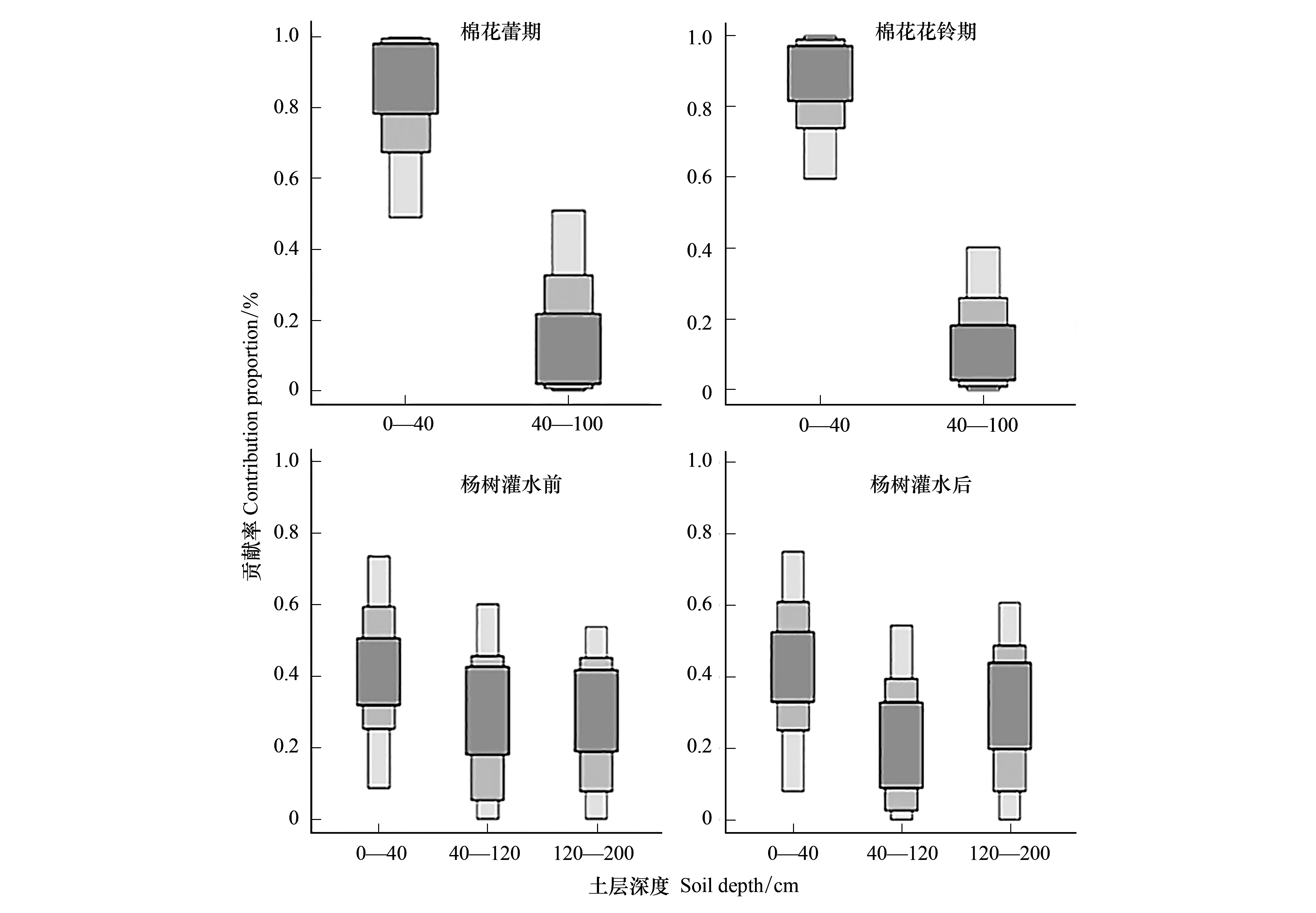
图5 棉花与杨树在不同生长季各土层水分的贡献率Fig.5 Proportion source contribution of different soil depths to cotton and poplar箱形图表示50%,75%,95%的置信区间
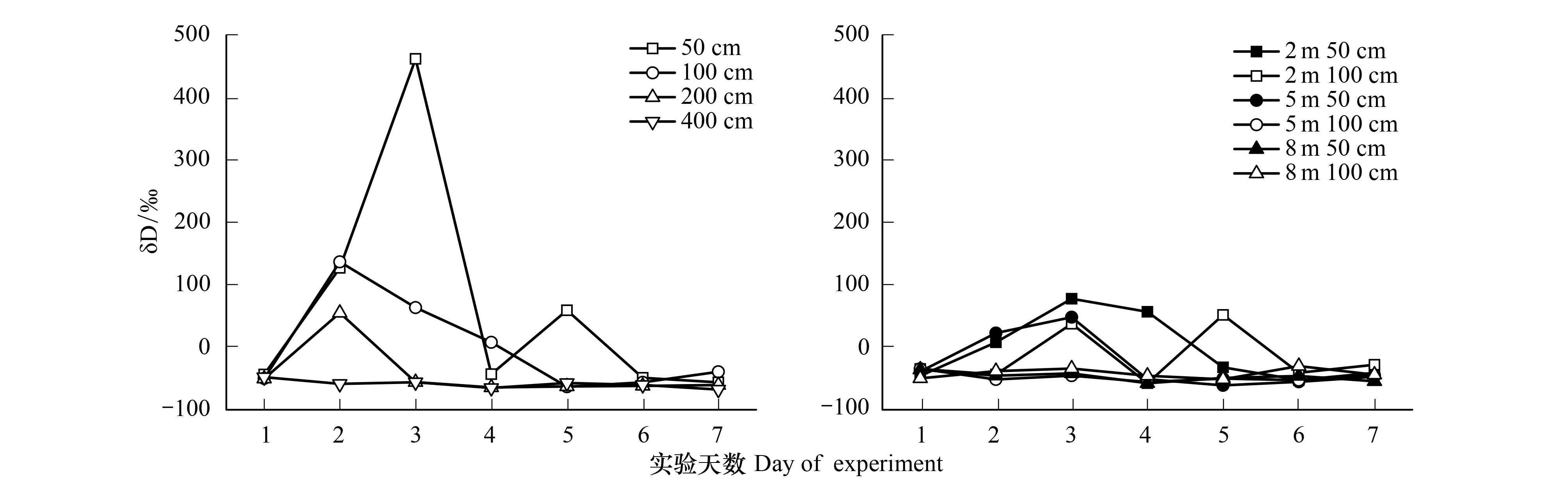
图6 不同土壤深度和距离杨树木质部人工添加δD同位素值Fig.6 The articial δD isotope values of poplar during the experiment for labeling different depths
2.3 人工添加δD同位素方法确定农林复合系统植物根系分布
本研究中,通过添加δD同位素探测杨树根系的分布,判断其可能对棉花生长的影响范围。图6可知,δD同位素在实验处理后信号显著增强,在土壤50、100与200 cm深处都显示出非常高的δD值;然而,在土壤400 cm并没有检测到明显的δD值上升。在探测杨树侧根水平分布于棉田的处理中(图6),发现δD值在距离树干2 m处、距地表50 cm与100 cm深处有明显增加;在距树干5 m处和距地表50 cm棉田深处显示有δD同位素增大信号,100 cm深处并没有发现;而在远距树干8 m处,距地表50 cm 和100 cm深处,未出现杨树δD同位素值异常升高现象。因此,可根据杨树木质部δD同位素值,推测出莫索湾杨树主要根系分布在200 cm土层以内,但深度不超过400 cm;并在棉田一侧,侧根可延伸到5 m远、距地表50 cm土层深处;8 m处并没有大量杨树根系存在此外;在距杨树3 m处、距地表50 cm到100 cm深处都有杨树根系存在,表明此处杨树根系密度较高。
3 讨论
3.1 棉花与杨树水分利用策略
土壤水作为干旱区限制植物生长的重要因素,对整个干旱区生态环境具有重要意义[22]。而干旱区植物根系分布则反应了植物在极端缺水环境中适应性[23]。本研究中,棉花花铃期土壤表层含水量(0—40 cm)高于棉花蕾期(图3),图4与图5中,棉花主要水分来源反而向下迁移,这反映了棉花作为直根系深根作物,随着棉花生育期的进行,根系也不断向下生长,扩大其有效资源利用范围。然而,Wang等[15]分别对华北平原不同生长季棉花水分利用研究,发现棉花在花铃期主要利用50 cm以下土壤水分,这与本文中棉花水分来源在花铃期在40 cm之下仅为14%有较大差距。造成这种差距的原因可能是在本研究中随着灌溉量与灌溉频次的增加,尤其是膜下滴灌技术的应用,土壤浅层含水量增加,棉花根系大量有向表层聚集的趋势[24]。从图3中,莫索湾由于采用膜下滴管技术,表层土壤含水量一直保持在较高水平,并在花铃期灌溉后表层土壤含水量大于深层,导致棉花在未受到干旱胁迫情况下,根系向下生长的驱动力变弱,以吸收浅层土壤水分为主。因此,可推测莫索湾绿洲棉花灌溉水量与频次相对较高,棉花并未过多利用到较深层土壤水分,造成该地区水资源存在潜在的浪费。可采取适当降低灌溉水量,迫使棉花扩大根系分布,提高水分利用范围和效率。
杨树水分利用策略在大水漫灌前后短期内并未发生明显的水分利用迁移现象。虽在漫灌后,主要水分来源从20—60 cm土层迁移到0—40 cm(图4),但总体上0—40 cm土层贡献率差别并不明显(41%与43%, 图5),120—200 cm土层可供给总需水量约30%。从图3可知,杨树在漫灌前深层土壤含水量较表层大。表层土壤由于长时间未得到充足的灌溉水补给(2016年8月之前没有灌溉),再加上本地区较小的降水量与强烈的蒸散发,导致表层土壤含水量较小。Yi等[25]研究表明农田的侧面补给是河西走廊防护林重要的深层土壤水分来源,而较深层来自农田的侧面补给,导致土壤含水量相对较高,杨树根系大量分布于此,而表层根系分布相对较少。因此即使表层土壤含水量迅速增加,但水分贡献率变化幅度依然有限。进一步通过δD同位素示踪,杨树根系至少深入200 cm土层(图6),较大的根系分布范围为其提供了较大的水分利用区间。这与 Everson 等[26]在南非发现干旱环境中,大量防护林根系分布在土壤深层的结果较为一致。
3.2 水分竞争与协同
农林复合系统中平衡乔木与农作物竞争与协作是研究的主要内容之一[27]。乔木与农作物适当的种植距离,根系空间分布互补对于提高作物产量和养分水分利用效率具有重要意义[28]。本文中杨树对棉花水分养分的主要影响是通过其侧根延伸入棉田。从图6可推测,杨树对棉花水分养分的竞争一般发生在8 m以内,5 m内棉花生长必定受到杨树影响。在距杨树3 m处杨树根系密度较高,对棉花水分养分的潜在竞争增大。虽然杨树与棉花主要水分利用区间有一定偏差(杨树约40%的水来自0—40 cm,而棉花则为80%以上),但由于杨树巨大的耗水量(Fu等[18]发现莫索湾同一地区杨树单月最高耗水量可达307 mm),对棉花水分养分竞争依然强烈。而通过人为干预,如修剪乔木冠幅,增加肥料投入等可以降低乔木对农作物影响[29]。本研究区中5 m范围内杨树对棉花影响最为严重,因此在5 m范围内增加化肥投入与灌溉量部分抵消杨树对棉花水分养分的竞争影响,并进一步通过在5 m范围内对杨树根系的修剪可显著降低其对农作物的影响。从图4与图5,棉花主要利用表层土壤水分,对40 cm以下的土壤水分利用有限,而通过杨树根系吸收深层水分,提高了整个干旱区生态系统的水分利用效率。此外由于防护林在新疆绿洲中的生态意义,通过量化杨树对棉花的影响范围估算农民损失,从而为平衡防护林生态价值与经济价值意义重大。
3.3 人工添加同位素的应用
植物根系分布决定了其所能利用的资源范围[30],根系范围越大,表明其可以利用资源选择余地就越大。本研究中,杨树根系侧向分布范围越大,意味着对棉花潜在的水分养分竞争。然而,单纯的用稳定同位素分析方法,并不能准确判断出植物根系分布范围,由于植物根系活性的可塑性[9],稳定同位素方法只能计算出某一时的植物水分来源,用它很难准确推断植物根系分布;此外,如果土壤剖面同位素值变异较小,或者拐点较多,用传统的稳定同位素方法出现的误差较大。本文通过应用人工添加同位素方法,量化了杨树的根系分布,并通过观察试验处理后第7天δD同位素在土壤剖面的分布,发现并未出现显著向上迁移(图7)。这与Beyer等[13]结论相一致,表明人工δD同位素添加可以探测植物根系分布,可作为一种稳定同位素研究植物根系水分的辅助方法去推广应用。

图7 100 cm和 200 cm土壤深度人工添加δD同位素运移Fig.7 The dynamic of articial δD isotope after seven days of experiment for labeling depths of 100 cm and 200 cm in poplar plots
4 结论
(1)棉田在膜下滴灌技术下,土壤含水量较高,从蕾期到花铃期水分主要来源向下迁移(0—20 cm至20—40 cm),0—40 cm土层贡献了85%以上棉花水分来源。杨树由于表层的土壤水分蒸发,0—40 cm土壤含水量较低,迫使根系大量生长于土壤较深层,导致杨树在灌溉前后,土壤40 cm以下为其主要供水来源区间(贡献率约60%),同时土壤主要供水层在灌溉后出现向上迁移趋势(20—60 cm至0—40 cm)。因此综合得出棉花与杨树主要水分来源的土壤深度有明显分层,但由于杨树巨大的含水量,对棉花的水分竞争不可避免,主要发生在土壤0—40 cm深度处。
(2)通过人工添加δD同位素,探测出杨树根系垂直深度在土壤200—400 cm处,水平可延伸到棉田5—8 m处,超过8 m杨树根系分布的可能性较小,且在3 m处根系大量分布。这意味着距树干5 m内杨树与棉花肯定存在强烈的水分竞争,5—8 m是过渡区;这对防护林与农作物的配置管理和对农民经济补偿有指导和借鉴意义,进而维持整个新疆干旱区生态平衡。
(3)本文通过焦点法与SIAR模型,较为精确的算出棉花与杨树的主要水分利用区间,并进一步通过人工添加δD同位素方法弥补了上述方法上的不足,并且量化了杨树根系的空间分布,从而多维角度对植物水分和根系其进行分析解释,对以后稳定同位素方法在生态环境中应用具有借鉴意义。
