莫海威芽孢杆菌产壳聚糖酶的分离纯化及酶学性质分析
2019-09-23刘进杰吕娟娟陈国忠
刘进杰 吕娟娟 陈国忠

摘要:纯化从烟台海域沙质土壤分离得到的莫海威芽孢杆菌菌株amyP216来源壳聚糖酶,并对其进行酶学性质分析。以发酵液为原料,依次进行超声波破碎菌体、20%~70%硫酸铵分级盐析、纤维素DE-32阴离子交换层析、葡聚糖凝胶G-75过滤层析,得到壳聚糖酶。采用SDS-PAGE凝胶电泳测定分子量为30 ku,全波段扫描结果显示在 324 nm 处出现最高峰。然后对壳聚糖酶的酶学性质进行研究。结果表明,最适反应温度为55 ℃,且在一定范围内温度越低,其热稳定性越高;最适反应pH值为5.5,中性条件下酶活性最稳定;在金属离子中Mn2+对其激活作用最明显,其他金属离子对其有抑制作用;该酶具有相对较好的底物特异性。
关键词:壳聚糖酶;莫海威芽孢杆菌;分离纯化;酶学性质;底物特异性
中图分类号: S188+.3 文献标志码: A 文章编号:1002-1302(2019)14-0231-05
壳寡糖在食品、医药、农业等行业都有着很高的应用价值,具有抗疲劳、调节免疫力和抗肿瘤的功效[1-4],可有效抑制肝损伤[5-6],还具有降血脂、降血糖的功能[6-9]。壳寡糖还可用于调节植物生长、延长果蔬保鲜等[10-13]。壳寡糖是由壳聚糖酶专一性地降解壳聚糖后生成的,因此寻求具备高效降解能力的壳聚糖酶,具有较高的工业应用价值。
壳聚糖酶是一类可催化氨基葡萄糖间的β-1,4-糖苷键断裂的酶[14],其来源广泛,广泛存在于一些真菌、细菌、放线菌中,甚至在植物病原菌、病毒中也有存在[15-19]。由于壳聚糖酶来源不同,酶学性质也就不同,酶解产物壳寡糖的聚合度也会有所不同[20]。因此,对不同微生物来源的壳聚糖酶进行分离纯化,研究其酶学性质,已经成为国内外的研究热点。
前期工作中,笔者在烟台海域沙质土壤中分离到1株壳聚糖酶产生菌株——莫海威芽孢杆菌(Bacillus mojavensis)amyP216,并对其发酵产生壳聚糖酶的条件进行了优化[21]。在本研究中,对发酵液中的壳聚糖酶进行分离纯化,探讨其酶学性质,为该壳聚糖酶的更深入研究奠定理论基础,也为该酶的工业应用提供依据。
1 材料与方法
1.1 试验材料与设备
1.1.1 菌株 莫海威芽孢杆菌(Bacillus mojavensis)菌株amyP216,由笔者所在实验室从烟台海岸带沙质土壤分离筛选获得。
1.1.2 试剂 磷酸二氢钠、磷酸氢二钠、醋酸、乙酸钠、3,5-二硝基水杨酸、氢氧化钠、四水酒石酸钾钠、苯酚、无水亚硫酸钠,均为国产分析纯;葡聚糖凝胶G-75(Pharmacia)、纤维素DE-32,购自江苏南京奥朵福尼生物科技有限公司;壳聚糖,购自山东济南海得贝生物工程有限公司;标准蛋白marker,由中国科学院上海生物化学研究所监制。
1.1.3 仪器设备 Ф3.5 cm×70 cm层析柱,购自上海华美实验仪器厂;HL-2恒流泵,购自上海沪西仪器厂有限公司;SBS-100数控记滴自动部分收集器,购自上海青浦沪西仪器厂;Z-323-K高速冷冻离心机、TDL-60B低速台式离心机,购自上海安亭科学仪器厂;WFJ7200可见光分光光度计,购自尤尼科(上海)仪器有限公司;MD34透析袋(美国Viskase);BS224S电子分析天平,购自北京赛多利斯仪器有限公司;KDS超声波细胞粉碎机,购自浙江宁波新芝科技有限公司;HH-6水浴锅,购自江苏省金坛市双捷实验仪器厂。
1.2 菌株培养
1.2.1 培养基 LB斜面培养基:酵母浸粉5 g/L、蛋白胨 10 g/L、NaCl 2.5 g/L、琼脂20 g/L,pH值7.0。
液体种子培养基:胶体壳聚糖10 g/L、酵母浸粉5 g/L、MgSO4 0.5 g/L、NaCl 2.5 g/L、KH2PO4 2 g/L,pH值6.5。
发酵培养基:壳聚糖20 g/L、酵母浸粉10 g/L、MgSO4 0.5 g/L、NaCl 2.5 g/L、KH2PO4 2 g/L、CaCl2 0.1 g/L,pH值6.5[21]。
1.2.2 培养方法 菌种活化:取甘油管保藏菌种划线至LB斜面培养基,30 ℃培养24 h。
种子液:挑取单菌落接种至液体种子培养基,30 ℃、200 r/min 培养24 h。
摇瓶发酵:500 mL三角瓶装50 mL发酵培养基,按5%接种量将种子液接入发酵培养基,30 ℃、220 r/min培养60 h。
发酵罐培养:按5%接种量将种子液接入装有3 L发酵培养基的5 L自控发酵罐中,转速600 r/min,通气量 2 L/(L·min),温度30 ℃[21]。
1.3 壳聚糖酶的分离纯化
取发酵罐培养的发酵液,4 ℃、8 000 r/min离心15 min后,收集菌体。超声波破碎20 min后,4 ℃、6 000 r/min离心10 min,取上清液,加入硫酸銨至饱和度20%,4 ℃过夜,10 000 r/min 低温离心30 min,取上清液,继续加入硫酸铵固体至饱和度70%,4 ℃过夜,10 000 r/min低温离心30 min,收集沉淀。将沉淀溶解于0.02 mol/L磷酸盐缓冲液(pH值8,含1 mol/L NaCl)中,透析浓缩后得粗酶液[16,22]。
取粗酶液3 mL进行纤维素DE-32柱层析,并用 0.02 mol/L 磷酸盐缓冲液(pH值8,含1 mol/L NaCl)进行洗脱,合并有酶活的部分并浓缩。然后采用葡聚糖凝胶G-75柱层析,并用0.02 mol/L磷酸盐缓冲液(pH值8,含1 mol/L NaCl)作为洗脱液进行洗脱,合并有酶活的部分并进行浓缩[22-24]。
浓缩后的样品采用SDS-PAGE凝胶电泳的方法进行酶蛋白的纯度分析以及相对分子质量的测定,分离胶的质量分数为15%,浓缩胶质量分数为5%[25]。
1.4 壳聚糖酶的酶学性质研究
1.4.1 温度及最适温度的测定 将酶液与壳聚糖底物分别放于35、45、55、65、75、85 ℃水浴中反应15 min,以确定酶的最适反应温度。将酶液在上述温度中保温120 min,以探讨热稳定性。
1.4.2 pH值及最适pH值的测定 将酶液与壳聚糖底物放于pH值分别为4、5、6、7、8、9的磷酸缓冲液中反应30 min,测定酶活,以确定最适反应pH值。将酶液放于上述的缓冲液中,保温2 h,测定酶活性,以探讨pH值稳定性。
1.4.3 金属离子对酶活性的影响 等量酶液与底物混合,加入不同金属离子,使离子浓度达到0.5 mmol/L。不同金属离子分别为Mn2+(MnSO4)、Mg2+(MgSO4)、Ca2+(CaCL2)、Cu2+(CuSO4)、Fe3+[Fe2(SO4)3]、Ba2+(BaSO4)、Li+(Li2SO4)、Zn2+(ZnSO4),以不加金属离子为对照,计算相对酶活性并作图,以探讨金属离子对酶活性的影响。
1.4.4 底物特异性 分别取1%壳聚糖醋酸溶液、胶体壳聚糖、羧甲基壳聚糖、胶体甲壳素、羧甲基纤维素钠作为底物反应,保温15 min,以壳聚糖为底物作为对照,计算相对酶活性并作图,以探讨壳聚糖酶对底物的专一性。
1.5 检测方法
1.5.1 壳聚糖酶活性测定 采用3,5-二硝基水杨酸(DNS)测定还原糖的方法测定壳聚糖酶的活性[16,24]。以氨基葡萄糖为标准绘制标准曲线,其回归方程为y=0.083 93x+0.033 7。式中:y为吸光度D值;x为浓度,μmol/mL;r2=0.996 77。酶活性单位定义为1 mL发酵液 1 min 下释放生成的微摩尔还原糖。
1.5.2 蛋白质浓度测定 蛋白质浓度采用Bradford法[25],以牛血清白蛋白(BSA)为标准绘制标准曲线,牛血清蛋白标准曲线的回归方程为y=1.593 43x-0.042 12。式中:y为D值;x为浓度,μmol/mL;r2=0.999 26。
1.6 数据处理与分析
利用SPSS 19.0进行数据处理和误差分析,用Origin 8绘图软件绘制图形。每组试验做3个平行试验,以3组数据进行误差分析以得到准确的试验结论。
2 结果与分析
2.1 壳聚糖酶的分离纯化
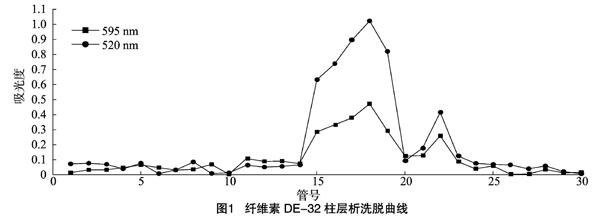
将破壁、盐析、透析后壳聚糖酶粗酶液上样于磷酸盐缓冲液平衡好的DEAE层析柱,随后用0.02 mol/L磷酸盐缓冲液(pH值8,含1 mol/L NaCl)进行洗脱,洗脱结果如图1所示。试验共收集30管样品,每管2.5 mL。在第18管处出现第1个酶活性和蛋白质浓度最高峰,且酶活性峰值与蛋白浓度峰值基本重合;在第22管处出现第2个酶活性和蛋白质浓度高峰,但其酶活性和蛋白质含量明显低于第1个峰。可见,壳聚糖酶的酶活性主要集中在第1个蛋白峰处。
把经纤维素DE-32洗脱得到的第1个蛋白峰处的样品收集、浓缩,上样到葡聚糖凝胶G-75层析柱进一步分离纯化,洗脱图谱如图2所示。试验中共收集17管样品,每管 4 mL,酶活性检测发现第6管和第11管处的蛋白样有明显的壳聚糖酶活性,第6管酶活性较高。蛋白峰出现在第5管和第10管处,第5管处蛋白浓度略高。
由表1可知,经过盐析、纤维素DE-32交换层析和葡聚糖凝胶G-75层析等一系列纯化后,壳聚糖酶比活力达到 8.01 mol/(min·mg),回收率为4.4%。
对葡聚糖凝胶G-75层析中第6管处样品进行SDS-PAGE电泳检测其分析纯度和相对分子质量,结果如图3所示。样品为单一条带,说明所分离的壳聚糖酶纯度较高,与标准蛋白marker比对,可以得出分子量约为30 ku,这与Shimosaka等(31.9 ku)[19]、孙菲等(30 ku)[23]、段妍等(30.5 ku)[24]所纯化的壳聚糖酶相对分子质量大致相当;但是与Gao等(41 ku)[26]、Muslim等(45 ku)[18]、韩宝芹等(66.2 ku)[27]、Wang等(21、18 ku)[28]、Shehata等(130 ku)[29]纯化相差较大。可见,不同来源的壳聚糖酶的相对分子质量有明显的差异。
由图4可知,壳聚糖酶在324 nm处有最大吸光度,数值为0.265,具有一般酶蛋白的紫外吸收特征。
2.2 壳聚糖酶酶学性质研究
2.2.1 温度及温度稳定性测定 由图5可知,曲线呈现先上升后下降的趋势,最高点出现在55 ℃处。55~65 ℃,曲线呈直线下降,当超过65 ℃时,酶活性降至很低,可能是因为温度过高使壳聚糖酶变性失去活性。可见,壳聚糖酶反应的最适温度在55 ℃,这与杨立红等所纯化的壳聚糖酶的最适温度[16,22,30]相同,但是与Gao等(60 ℃)[26]、Wang等(50 ℃)[28]、Shehata等(40 ℃)[29]所纯化的壳聚糖酶的最适温度不同。
由图6可见,在一定范围内,温度越低,对酶的稳定性越好。所有曲线都呈现下降趋势。在保温60 min后50、60 ℃ 保温条件下,酶活性均消失,在60 ℃下壳聚糖酶更快失去酶活;保溫90 min后,30、40 ℃也失去了酶活,40 ℃保温条件下,相对酶活性下降得更快,即壳聚糖酶更容易失去酶活性。
2.2.2 pH值及pH值稳定性测定 pH值对壳聚糖酶活性的影响如图7所示,曲线呈先上升后下降的趋势。在pH值5.5之前,酶活性随pH值的升高而上升,当pH值达到6以后,酶活性急剧下降,在pH值5.5处出现最高峰,可见壳聚糖酶反应的最适pH值为5.5,这与Shehata等(40 ℃)[29]、隋斯光等[31]研究的壳聚糖酶的最适pH值相同。当pH值为 6.0 时,本试验所纯化得到的壳聚糖酶的活性大大降低,但是Gao等纯化的壳聚糖酶的最适pH值却是6.0[26,32]。这表明不同来源的壳聚糖酶的最适pH值差距较大,莫海威芽孢杆菌amyP216来源的壳聚糖酶的最适pH偏酸性。
pH值对酶活稳定性的影响如图8所示。时间越长,相对酶活性越小,在pH值为7即中性环境下,相对酶活性降低较慢,说明保温时间对酶活性的影响最小;pH值6环境下,酶活性降低程度次之;pH值4和5对酶活性的稳定性影响较大,pH值为4时相对酶活性下降最快,对酶活性稳定性影响最大。由此可得,pH值7最適合壳聚糖酶的保存。
2.2.3 金属离子对酶活性的检测 金属离子对壳聚糖酶的影响结果如图9所示,本试验所验证的金属离子中,正1价金属离子Li+对酶活性有明显的抑制作用,在浓度达到 0.5 mmol/L 时,其相对酶活性为76.7%。正2价金属离子中,Mn2+对酶活性有激活作用,在金属离子浓度为0.5 mmol/L时,其相对酶活性达到263.1%,其原因可能与酶的活性中心有关。Mg2+、Ca2+、Cu2+、Ba2+、Zn2+等都有一定的抑制作用,其中Ba2+的抑制作用最弱,相对酶活性为90.1%,Cu2+的抑制作用最强,相对酶活性达到39.7%。正3价金属离子Fe3+对酶活性同样有严重的抑制作用,是以上8种离子中抑制作用最强的,相对酶活性达到34.3%。
在本试验所研究的金属离子中,只有Mn2+对酶活性有激活作用,其他金属对酶活性均有抑制作用。这与其他来源的壳聚糖酶的试验结果也有所不同。Wang等研究发现Mn2+会抑制Serratia marcescens TKU011来源的壳聚糖酶[28],周念波等研究发现Zn2+、Ca2+对Bacillus sp. LS产的壳聚糖酶有一定的激活作用[22],这进一步表明不同微生物来源的壳聚糖酶的酶学性质是有差异的。
2.2.4 底物特异性检测 如表2所示,壳聚糖酶对壳聚糖的水解能力最强,对羧甲基纤维素钠、DEAE纤维素和甲壳素也有一定的水解能力,其水解能力依次降低。可见,纯化所得的壳聚糖酶底物特异性较好。
3 结论
壳聚糖酶是理想的制备壳寡糖的酶制剂,本研究以高产壳聚糖酶的莫海威芽孢杆菌(Bacillus mojavensis) amyP216发酵液为原料,经过超声波破壁处理、硫酸铵分级沉淀、纤维素DE-32阴离子交换柱层析、葡聚糖凝胶过滤层析等方法纯化后,得到了电泳纯壳聚糖酶,纯化倍数达到25.21倍,相对酶活性为8.01 mol/(min·mg),回收率为4.4%。电泳法测定莫海威芽孢杆菌来源壳聚糖酶相对分子质量为30 ku,全波段扫描结果显示在324 nm处出现最高峰,峰值为0.265 Abs。
经酶学性质研究发现,该壳聚糖酶的最适反应温度为 55 ℃,在试验范围内,温度越低,酶的稳定性越好;最适反应pH值为5.5,在pH值7时酶的稳定性较好;在试验范围内,金属离子Mn2+对其激活作用最明显,Mg2+、Ca2+、Cu2+、Ba2+、Zn2+等金属离子对其有抑制作用;该酶具有相对较好的底物特异性。下一步研究将对该酶进行基因工程研究,以提高酶的表达量,为其产业化奠定基础。
参考文献:
[1]尹雨芳,林 强,曹建民,等. 壳寡糖抗运动疲劳及对运动性免疫抑制的影响[J]. 中国实验方剂学杂志,2016,22(4):146-149.
[2]张 沛,韩宝芹,陈列欢,等. 用酶解法制备壳寡糖及其对机体免疫功能的调节作用[J]. 中国免疫学杂志,2013,29(2):191-196.
[3]杨欢欢,周艳芬,武金霞,等. 壳寡糖诱导肺癌细胞A549凋亡及其机制初探[J]. 时珍国医国药,2013,24(2):268-270.
[4]刘晶莹,刘 洋,谭子强. 壳寡糖对S180肉瘤小鼠抗肿瘤作用的研究[J]. 国外医药(抗生素分册),2015,36(1):23-24.
[5]许青松,宫德正,邹 原,等. 两种壳寡糖对急性肝损伤模型小鼠的保护作用[J]. 医药导报,2008,27(2):153-155.
[6]申 杰,叶希韵,沈 菊,等. 壳寡糖对高脂血症小鼠降血脂及肝脏保护的作用[J]. 西北农林科技大学学报(自然科学版),2007,35(9):35-38,44.
[7]刘 冰,秦贞奎,林祥梅,等. 壳寡糖及其衍生物对糖尿病大鼠血脂代谢紊乱的影响及对肝脏的保护作用[J]. 暨南大学学报(自然科学与医学版),2009,30(4):388-394.
[8]王 斌,苏谦俊,张思聪,等. 壳寡糖和壳聚糖对高脂膳食小鼠的降脂功能比较研究[J]. 天然产物研究与开发,2017,29(6):1025-1029,1047.
[9]赵金山,张 燕,延 岩,等. 壳寡糖降血糖作用的临床观察研究[J]. 食品与药品,2011,13(3):120-122.
[10]徐小菊,林玉平,金 伟,等. 壳寡糖对夏黑葡萄果实膨大和品质的影响[J]. 中外葡萄与葡萄酒,2017(2):26-29.
[11]狄文伟. 不同分子量壳寡糖对黄瓜穴盘苗生长的影响[J]. 江苏农业科学,2016,44(4):196-198.
[12]闫佳琪,张忆楠,赵玉梅,等. 壳寡糖控制果蔬采后病害及诱导抗病性研究进展[J]. 食品科学,2015,36(21):268-272.
[13]邓丽莉,尹保凤,曾凯芳. 壳寡糖和壳聚糖处理对“乙烯褪绿蜜橘”果实品质和抗病性的影响[J]. 现代食品科技,2017,33(2):167-175.
[14]季更生,陈爱春. 微生物壳聚糖酶的研究进展[J]. 食品科学,2010,31(3):297-301.
[15]史凤玉,蔡爱军,田 辉,等. 来自拟青霉属真菌的壳聚糖酶的分离纯化、理化性质及降解产物的分析[J]. 菌物学报,2013,32(4):721-728.
[16]杨立红,程仕伟,冯志彬,等. 链霉菌壳聚糖酶的纯化及其酶学性质[J]. 生物加工过程,2013,11(3):52-58.
[17]刘怀伟,鲍晓明. 腐皮镰孢菌壳聚糖酶的酶学性质研究及其在酿酒酵母工业菌株中的表达[J]. 微生物学报,2009,49(12):1607-1612.
[18]Muslim S N,Al-Kadmy I M,Hussein N H,et al. Chitosanase purified from bacterial isolate Bacillus licheniformis of ruined vegetables displays broad spectrum biofilm inhibition[J]. Microbial Pathogenesis,2016,100:257-262.
[19]Shimosaka M,Kumehara M,Zhang X Y,et al. Cloning and characterization of a chitosanase gene from the plant pathogenic fungus Fusarium solani[J]. Journal of Fermentation and Bioengineering,1996,82(5):426-431.
[20]Somashekar D,Joseph R. Chitosanases - properties and applications:a review[J]. Bioresource Technology,1996,55(1):35-45.
[21]冯志彬,薛 钰,陈国忠,等. 1株产壳聚糖酶细菌的分离、鉴定和发酵条件优化[J]. 食品科学,2016,37(19):171-176.
[22]周念波,李轶群,涂绍勇. Bacillus sp.LS 壳聚糖酶的分离纯化及性质研究[J]. 食品科技,2008,33(2):4-7.
[23]孫 菲,陈山岭,李宜海,等. 壳聚糖酶高产菌株的筛选、产酶条件的优化及壳聚糖酶的分离纯化[J]. 现代食品科技,2006,22(3):21-23,20.
[24]段 妍,韩宝芹,董 文,等. 产壳聚糖酶菌株发酵条件优化及壳聚糖酶的分离纯化研究[J]. 海洋科学,2009,33(1):1-7.
[25]陈钧辉,李 俊,张太平,等. 生物化学实验[M]. 4版.北京:科学出版社,2008.
[26]Gao X A,Ju W T,Jung W J,et al. Purification and characterization of chitosanase from Bacillus cereus D-11[J]. Carbohydrate Polymers,2008,72(3):513-520.
[27]韩宝芹,杨菊林,刘万顺,等. 壳聚糖酶的分离纯化及性质研究[J]. 中国海洋大学学报(自然科学版),2006(2):255-260.
[28]Wang S L,Peng J H,Liang T W,et al. Purification and characterization of a chitosanase from Serratia marcescens TKU011[J]. Carbohydrate Research,2008,343(8):1316-1323.
[29]Shehata A N,Abd El Aty A A,Darwish D A,et al. Purification,physicochemical and thermodynamic studies of antifungal chitinase with production of bioactive chitosan-oligosaccharide from newly isolated Aspergillus griseoaurantiacus KX010988[J]. International Journal of Biological Macromolecules,2017,107(Part A):990-999.
[30]王艳君,卓少玲,陈 盛,等. 产壳聚糖酶菌株的筛选、鉴定及酶学特性分析[J]. 微生物学通报,2012,39(12):1734-1745.
[31]隋斯光,方文建,郑连英. 壳聚糖酶的分离提纯及其酶学性质研究[J]. 高校化学工程学报,2007(5):814-819.
[32]蔡婀娜,黄惠莉,黃双燕. 海洋烟曲霉菌产壳聚糖酶的酶学性质初探[J]. 暨南大学学报(自然科学与医学版),2011,32(5):509-512.
