不同种类蔬菜幼苗对多环芳烃胁迫的生理反应
2019-09-17唐璇龙明华乔双雨
唐璇 龙明华 乔双雨

摘要:为了探索多环芳烃对蔬菜幼苗生长和生理特性的影响。分别以0.3、0.6、0.9 mg/L等不同浓度的16种多环芳烃对黄瓜、菜心、萝卜进行胁迫培养,以不加多环芳烃的幼苗为对照,研究多环芳烃胁迫下各蔬菜幼苗的生长和生理响应。结果表明,黄瓜的叶宽、鲜质量、CAT活性、POD活性以及萝卜的叶绿素b含量、CAT活性在0.3 mg/L PAHs处理时达到最小值。处理浓度为0.6 mg/L时,黄瓜的POD活性、菜心的维生素C含量和萝卜的MDA含量最小。黄瓜的干质量、菜心的鲜质量以及萝卜的鲜质量、POD活性在0.9 mg/L PAHs处理下达到最小值。不同浓度的PAHs处理对黄瓜和菜心叶片叶绿素的合成和光合作用以及萝卜的维生素C含量均有促进作用。随着PAHs处理浓度的升高,黄瓜和菜心的MDA含量呈上升趋势,在0.9 mg/L PAHs处理下达到最大值。不同种类蔬菜幼苗对PAHs具有不同生理反应的原因之一,可能与它们叶片结构的不同导致吸收PAHs量的差异有关。
关键词:多环芳烃;蔬菜幼苗;生长;生理生化
中图分类号: Q945.78 文献标志码: A 文章编号:1002-1302(2019)07-0166-04
多环芳烃(polycyclic aromatic hydrocarbons,简称PAHs)是一类由2个或2个以上经过高温裂解得到的苯环以直链状、角状或者簇状排列组成的稠环化合物,具有疏水性及低水溶性,易吸附于固体颗粒表面,能长期存在于环境中,是一种持久性有机污染物[1-3]。1976年美国环保局(USEPA)提出的129种“优先污染物”(priority pollutants)的黑名单中,有16种是PAHs类化合物[4-5];1990年,我国国家环境保护总局将7种致癌性较强的多环芳烃列入了我国环境优先污染物的黑名单[6]。PAHs在环境中难以降解,由于其具有半挥发性和脂溶性的特点,因而可以通过大气沉降作用经植物叶片进入植物体内或进入土壤中由植物根系吸收,并在植物体内迁移、代谢和积累,进而通过食物链危害到人类身体健康[7]。PAHs进入植物体的途径主要有2种:土壤-植物途径和空气-植物途径[8-9]。有报道认为,这种空气-植物体的吸收途径是植物吸收PAHs的主要途径[10-11],影响这种吸收途径的因素包括空气中PAHs的浓度、PAHs的性质、PAHs在空气中的存在形态、叶表面的特征及温度等。此外,植物叶表面的形态、表皮蜡质化学成分、气孔数量和分布以及绒毛生长的情况不同,对应植物与空气的相互作用过程也不同[12],且植物吸收PAHs的速率与植物本身的适应性及PAHs的生物有效性成正比[13]。
PAHs污染是全球面臨的一个紧迫的环境问题,由于其具有致癌、致畸、致突变的作用,严重影响人类健康,从而引起人们的高度重视。环境中的PAHs可以在植物体内迁移、代谢、累积,从而影响植物的正常生长,并可在食物链中被逐级放大,影响人类健康。目前,关于PAHs的研究主要集中在PAHs对环境的污染和PAHs污染土壤的治理(生物修复)方面,而关于PAHs胁迫对蔬菜幼苗影响的研究相对较少,本试验研究不同种类蔬菜幼苗对PAHs胁迫的生理反应,旨在为进一步探索不同种类蔬菜吸收及富集PAHs的特性打下基础。
1 材料与方法
1.1 试验材料
1.1.1 供试品种 F508油青甜菜心王,由广州市圣吉亚农业科技有限公司生产。白沙短叶13号早萝卜,由汕头市白沙蔬菜原种研究所生产。秀川701黄瓜,由广州市伟兴利种子有限公司生产。
1.1.2 供试土壤 采用基质和黄壤土(体积比1 ∶ 8)混合作为试验用土壤。
1.1.3 多环芳烃混合液的配制 16种PAHs混合标准品(10 mg/mL)购于美国Sigma-Alorich公司。16种多环芳烃分别为萘(NAP)、苊烯(ANY)、芴(FLU)、菲(PHE)、蒽(ANT)、荧蒽(FLT)、芘(PYR)、苯并(a)蒽(BaA)、(CHR)、苯并(b)荧蒽(BbF)、苯并(k)荧蒽(BkF)、苯并(a)芘(BaP)、茚并(1,2,3-c,d)芘(IPY)、二苯并(a,h)蒽(DBA)、苯并(g,h,i)二萘嵌苯(BPE)、苯并(e)芘(BeP)。
分别吸取一定体积的多环芳烃混合液于500 mL的丙酮溶液中,配成0.3、0.6、0.9 mg/L浓度的多环芳烃混合液,并放入4 ℃冰箱保存备用。
1.2 试验方法
1.2.1 试验设计 试验于2017年7月11日至10月20日于广西大学农学院蔬菜基地温室大棚内进行。本试验采用穴盘育苗,待幼苗长至2叶1心时,移栽于长60 cm、宽45 cm、高30 cm的泡沫箱中,每箱种9株。移栽3 d后,用不同浓度的PAHs混合液对幼苗叶片进行涂抹处理,以未作处理的蔬菜幼苗为对照。每隔3 d处理1次,共处理3次。每个处理设3个重复。
1.2.2 指标测定 生长指标的测定:第3次处理后的第3天进行株高、叶长和叶宽、鲜质量和干质量的测定。每处理设3个重复。之后采取幼苗的地上部分带回实验室,用水清洗叶片表面并用滤纸吸干表面水分,进行称量。称好质量后,用锡箔纸包好,并用液氮迅速冷却后,放入-40 ℃冰箱中保存备用。
生理指标的测定:叶绿素含量采用丙酮提取法[14]测定;维生素C含量采用2,6-二氯靛酚滴定法[15]测定;丙二醛(MDA)含量采用硫代巴比妥酸(TBA)法测定;过氧化物酶(POD)活性采用氧化愈创木酚法测定;过氧化氢酶(CAT)活性采用紫外分光光度法[16]测定。每个处理设3个重复。
1.3 数据处理
试验数据采用Excel 2013进行计算并制图,采用SPSS 17.0进行单因素方差分析,采用最小显著差数法(P<0.05,Duncans)进行不同处理间均值的差异显著性比较。
2 结果与分析
2.1 PAHs胁迫对蔬菜幼苗生长的影响
从表1、表2和表3可以看出,各PAHs处理之间以及各PAHs处理与对照之间的蔬菜幼苗株高差异不显著。对照处理的黄瓜幼苗叶长、鲜质量和干质量显著高于各PAHs处理,但各PAHs处理之间无显著差异。黄瓜的叶宽与鲜质量在低浓度PAHs处理下达到最小值,随着PAHs处理浓度的增高呈现上升的趋势,0.6、0.9 mg/L PAHs处理较0.3 mg/L PAHs处理分别上升了13.36%、18.19%和9.98%、19.57%;干质量随着PAHs浓度的增加逐渐减小,0.3、0.6、0.9 mg/L PAHs处理分别较对照处理下降了34.25%、36.46%、38.12%。菜心与萝卜的叶宽和干质量各处理间均无显著差异。菜心和萝卜在高浓度处理下鲜质量最小,但菜心在对照处理时鲜质量最大,而萝卜在中浓度PAHs处理下鲜质量最大。
2.2 PAHs胁迫对蔬菜幼苗叶绿素含量的影响
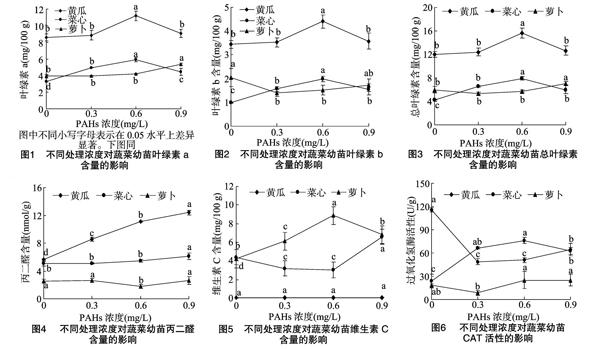
从图1、图2和图3可以看出,在0.6 mg/L PAHs处理下,黄瓜和菜心的叶绿素a、叶绿素b、总叶绿素含量最大。黄瓜的叶绿素a、叶绿素b、总叶绿素含量在0.3、0.9 mg/L PAHs处理和对照处理之间无显著差异。菜心的叶绿素a、叶绿素b、总叶绿素在对照处理下含量最小。萝卜的叶绿素a、总叶绿素在0.9 mg/L PAHs处理下含量最高,其他处理和对照处理无显著性差异。萝卜的叶绿素b含量在对照处理下最大,其他处理之间无显著性差异。表明0.6 mg/L PAHs处理对黄瓜和菜心的叶绿素含量有促进作用,0.9 mg/L PAHs处理对萝卜的叶绿素含量有促进作用。
2.3 PAHs胁迫对蔬菜幼苗丙二醛含量的影响
从图4可以看出,黄瓜和菜心的丙二醛含量随着PAHs处理浓度的增加呈现逐渐上升的趋势,0.3、0.6、0.9 mg/L PAHs处理分别较对照处理上升了51.85%、96.86%、120.24%和0.38%、6.98%、20.17%。萝卜的丙二醛含量在0.6 mg/L PAHs处理下最小,其他处理之间均无显著差异。结果表明,PAHs中低浓度胁迫下,黄瓜和菜心的膜系统能正常工作,高浓度PAHs胁迫下其膜脂过氧化作用加强,膜透性增加。
2.4 PAHs胁迫对蔬菜幼苗维生素C含量的影响
从图5可以看出,在对照和各浓度PAHs处理下,黄瓜的
维生素C含量均无显著性差异。在0.9 mg/L PAHs处理下,菜心的维生素C含量达到最大值,在0.6 mg/L PAHs处理下,维生素C的含量最小。在0.6 mg/L PAHs处理下,萝卜的维生素C含量达到最大值,对照处理的维生素C含量最小。结果表明,高浓度的PAHs处理对菜心的维生素C含量有明显的促进作用,中浓度的PAHs处理对菜心的维生素C含量有明显的抑制作用。各浓度的PAHs处理对萝卜的维生素C含量均有明顯的促进作用,其中中浓度PAHs处理对萝卜的促进作用最显著。
2.5 PAHs胁迫对蔬菜幼苗CAT活性的影响
从图6可以看出,0.3、0.6 mg/L PAHs处理对黄瓜的CAT活性有显著的抑制作用,对菜心的CAT活性有显著的促进作用。0.3 mg/L PAHs处理对萝卜的CAT活性有抑制作用,0.6、0.9 mg/L PAHs处理对萝卜的CAT活性有促进作用,但作用不显著。结果表明,黄瓜的CAT活性对中低浓度的PAHs较为敏感,萝卜在PAHs胁迫处理下还可以维持较好的清除自由基能力。
2.6 PAHs胁迫对蔬菜幼苗POD活性的影响
从图7可以看出,0.9 mg/L PAHs处理对黄瓜的POD活性有显著的促进作用,0.6 mg/L PAHs处理对黄瓜的POD活性有显著的抑制作用。0.3、0.9 mg/L PAHs处理对菜心的POD活性有显著的抑制作用,0.6 mg/L PAHs处理对菜心的POD活性有显著的促进作用。0.6 mg/L PAHs处理对萝卜的POD活性有显著促进作用,其他处理之间无显著差别。结果表明,不同蔬菜幼苗的POD活性对于PAHs胁迫有不同的响应。
3 结论与讨论
3.1 PAHs胁迫对蔬菜幼苗生长的影响
在不同浓度PAHs处理下,供试蔬菜幼苗的生长受到不同程度的影响。黄瓜的叶宽和鲜质量在0.3 mg/L PAHs处理时达到最小值,分别比对照处理减少了21.97%和38.84%;随着PAHs处理浓度的增大数值略微增加,但都比对照处理小。黄瓜的干质量随着PAHs处理浓度的增加逐渐减小,在0.9 mg/L PAHs处理下达到最小值,比对照处理减少了38.12%。菜心的鲜质量随着PAHs处理浓度的处理增加先增后减,在0.9 mg/L PAHs处理下达到最小值,比对照处理减少了27.88%。与对照相比,萝卜的鲜质量随着PAHs处理浓度的增加先减后增再减,在0.9 mg/L PAHs处理下达到最小值,对比照处理减少了17.97%。由结果可知,供试的3种蔬菜幼苗生长发育受PAHs危害大小的顺序为黄 瓜> 菜心>萝卜。
3.2 PAHs胁迫对蔬菜幼苗生理生化指标的影响
不同浓度的PAHs处理对黄瓜和菜心叶片叶绿素的合成和光合作用均有刺激作用。在0.3 mg/L PAHs处理下,萝卜的叶绿素b含量达到最小值,比对照处理减少了31.80%。随着PAHs处理浓度的增加萝卜的叶绿素b含量逐渐增加,但都比对照小。结果表明,PAHs处理对萝卜的叶绿素b含量有抑制作用。不同浓度的PAHs处理对萝卜的维生素C含量有促进作用;在0.6 mg/L PAHs处理下,菜心的维生素C的含量达到最小值,比对照减少了31.14%;随着处理浓度的增加维生素C的含量先减少再增加,在0.9 mg/L处理下高于对照。结果表明,中低浓度的PAHs处理对萝卜的维生素C的含量有抑制作用。丙二醛(MDA)是生物膜系统膜脂过氧化的产物之一,其在植物体内的含量可反映机体内脂质过氧化的程度并间接反映细胞受损的程度,植物在受到外界因素的胁迫时,MDA的含量会增加,膜受到的损伤会随之增大[17]。随着PAHs处理浓度的升高,黄瓜和菜心的MDA含量呈上升趋势,在0.9 mg/L PAHs处理下达到最大值,分别比对照处理增加了120.24%和20.17%。PAHs处理浓度为0.6 mg/L时,萝卜的MDA含量低于对照,这可能是由于中浓度的PAHs对萝卜的正常应激性机制有一定伤害,导致MDA含量降低。结果表明,PAHs的高浓度处理胁迫会引起黄瓜和菜心的脂质过氧化。
酶活性是植物在PAHs胁迫下较为敏感的生理指标,黄瓜和萝卜的CAT活性对低浓度的PAHs处理有一定的应激性反应,均在0.3 mg/L PAHs处理下达到最小值,分别比对照处理降低了57.68%和54.06%。在0.3 mg/L PAHs处理下菜心的POD活性达到最小值,比对照处理降低了30.13%。在0.6 mg/L PAHs处理下黄瓜的POD活性达到最小,比对照处理降低了 38.27%。在0.9 mg/L PAHs处理下,萝卜的POD活性达到最小值,比对照处理降低了13.13%。结果表明,黄瓜和萝卜CAT活性以及菜心的POD活性对低浓度的PAHs处理较为敏感,黄瓜的POD活性对中浓度的PAHs处理较为敏感,萝卜的POD活性对高浓度的PAHs处理较为敏感。
叶片吸收PAHs的速率受叶片比表面积的影响很大。Schrelber等对5种针叶植物吸收PAHs速率的研究表明,不同植物的吸收速率与比表面积具较好的相关性,同一环境下叶片的比表面积差异导致其吸收速率可能相差一个数量级[18]。叶片吸收积累PAHs的能力与叶片表面粗糙程度或绒毛疏密程度有较好的相关性。一方面有茸毛的植物叶片更能吸附空气中的颗粒污染物,密集的茸毛可以阻止小水滴接触叶片的蜡质表层,从而防止它们带走叶片表面附着的颗粒物[19]。Howsam等对同一环境中榛树、梣树和橡树进行对比研究,发现绒毛细密的榛树叶片中的PAHs浓度比绒毛稀疏的梣树、橡树叶片高1倍[20]。另一方面,叶片有排水的生理现象,这样会消除一部分叶片上滞留的颗粒物。黄勇发现,樟树、桂花、广玉兰、红檵木4个树种叶片解剖结构与富集PAHs能力的相关关系均达到显著水平,其中栅栏组织厚度、海绵组织厚度和叶片总厚度与叶片PAHs含量呈负相关关系,叶片栅栏组织厚度、海绵组织厚度、叶片总厚度越薄,越方便气态和颗粒态进入,吸附量也越多,对气态和颗粒态PAHs产生较强的富集作用;而种间其他叶片结构与叶片PAHs含量间相关关系不显著,可能是因为生理特性的差异,表现为某些结构上的差异对吸附不敏感,如有的角质层可能较难穿透、不同植物间气孔能力不一致、不同树种叶片气孔对粒径的选择不一致等[21]。Barber等研究发现,角质层较难穿透且气孔密度较高时,气孔吸收途径相对重要,而当角质层极易穿透时,气孔的作用几乎为零[22]。由此可知,不同种类蔬菜幼苗对PAHs具有不同的生理反应的原因之一,可能与它们叶片结构的不同导致吸收PAHs量的差异有关。
参考文献:
[1]李玉龙. 土壤中多环芳烃的迁移转化规律及其对植物生长的影响[D]. 西安:西安建筑科技大学,2015.
[2]Hadibarata T,Kristanti R A. Fate and cometabolic degradation of benzo[a]pyrene by white-rot fungus Armillaria sp. F022[J]. Bioresource Technology,2012,107:314-318.
[3]Quilliam R S,Rangecroft S,Emmett B A,et al. Is biochar a source or sink for polycyclic aromatic hydrocarbon (PAH) compounds in agricultural soils?[J]. Global Change Biology Bioenergy,2013,5(2):96-103.
[4]Keith L H,Telliard W A. Priority pollutants Ⅰ. A perspective view[J]. Environmental Science and Technology,1979,13(4):416-423.
[5]Carmichael L M,Christman R F,Pfaender F K. Desorption and mineralization kinetics of phenanthrene and chrysene in contaminated soils[J]. Environmental Science and Technology,1997,31(1):126-132.
[6]劉魏魏. 多环芳烃污染农田土壤的生物协同修复及有机废物调控强化修复技术[D]. 南京:南京农业大学,2009.
[7]殷 婧,夏忠欢,周彦池,等. 临汾市售蔬菜中多环芳烃污染特征及致癌风险分析[J]. 生态毒理学报,2016,11(3):265-271.
[8]Cobbett C S,Meagher R B. Arabidopsis and the genetic potential for the phytoremediation of toxic elemental and organic pollutants[J]. The Arabidopsis Book,2002,1:e0032.
[9]Harveyp J,Campanella B F,Castro P M L,et al. Phytoremediation of polyaromatic hydrocarbons,anilines and phenoles[J]. Environmental Seience Pollution Research International,2002,9(1):29-47.
[10]Wild S R,Jones K C. The significance of polynuclear aromatic hydrorbons applied to agricultural soils in sewage sludges in the UK[J]. Waste Management and Research,1994,12(1):49-59.
[11]Horstmann M,Mclachlan M S. Atmospheric deposition of semivolatile organic compounds to two forest canopies[J]. Atmospheric Environment,1998,32(10):1799-1809.
[12]Trapp S,McFarlane J C. Plant contamination:modeling and simulation of organic chemical process[M]. Boca Raton:Lewis,1995:254.
[13]Xu S Y,Chen Y X,Wu W X,et al. Enhanced dissipation of phenanthrene and pyrene in spiked soils by combined plants cultivation[J]. Science of the Total Environment,2006,363(1/2/3):206-215.
[14]張志良,瞿伟菁,李小芳. 植物生理学实验指导[M]. 北京:高等教育出版社,2009.
[15]刘 萍,李明军. 植物生理学实验技术[M]. 北京:科学出版社,2007.
[16]路文静,李奕松. 植物生理学实验教程[M]. 北京:中国林业出版社,2012.
[17]Tewari R K,Kumar P,Sharma P N. Magnesium deficiency induced oxidative stress and antioxidant responses in mulberry plants[J]. Scientia Horticulturae,2006,108(1):7-14.
[18]Schrelber L,Schonherr J. Uptake of organic chemicals in conifer needles:surface adsorption and permeability of cuticles[J]. Environmental Science & Technology,1992,26:153-159.
[19]Little P,Wiffen R D. Emission and deposition of petrol engine exhaust Pb-Ⅰ. Deposition of exhaust Pb to plant and soil surface[J]. Atoms Environ,1977,11(5):437-447.
[20]Howsam M,Jones K C,meason P. PAHs associated with the leaves of three deciduous tree species:Ⅰ. Concentrations and profiles[J]. Environmental Pollution,2000,108:413-424.
[21]黄 勇. 城市植物叶片PAHs特性及对土壤微生物与酶的影响[D]. 长沙:中南林业科技大学,2011.
[22]Barber J L,Kurt P B,Thomas G O,et al. Investigation into the importance of the stomatal pathway in the exchange of PCBs between air and plants[J]. Environmental Science & Technology,2002,36:4282-4287.李俊强,林利华,张 帆,等. 施肥模式对茶叶营养累积及土壤肥力的影响[J]. 江苏农业科学,2019,47(7):170-174.
