43种植物CaM1基因密码子使用特征及遗传差异分析
2019-09-02李晨辉赵子捷陈文烨焦义然杨帆刘永伟董福双杜进民周硕
李晨辉 赵子捷 陈文烨 焦义然 杨帆 刘永伟 董福双 杜进民 周硕

摘要:为深入了解不同类植物CaM1基因(calmodulin 1,简称CaM1)在抵抗逆境进化过程所形成的密码子的使用特征和遗传差异。运用CodonW、BioEdit和MEGA软件对43种植物序列进行分析,并构建系统发育树。结果表明,CaM1基因对密码子的使用具有极强的偏好性,CUC、AUC、UCU、CCA、ACU、UGA为其最优密码子,禾本目植物更偏好使用G/C碱基。G、C碱基含量和位置是引起CaM1基因密码子偏性的主要原因。遗传距离分析表明,山柑目与白花菜目之间的亲缘关系最近,豆目和无患子目间的亲缘关系次之,与聚类分析结果相符。
关键词:CaM1;密码子偏性;遗传距离;亲缘关系
中图分类号: S188 文献标志码: A 文章编号:1002-1302(2019)05-0028-04
收稿日期:2017-11-30
基金项目:国家自然科学基金青年科学基金(编号:31600216);河北省自然科学基金青年基金(编号:C2017301066);河北省现代农业创新工程项目(编号:2019-4-8)。
作者简介:李晨辉(1984—),男,河北保定人,博士,工程师,主要从事生物工程研究。E-mail:li.chen.hui@163.com。
通信作者:杜进民,博士,教授,主要从事生物技术研究,E-mail:13933106571@163.com;周 硕,博士,副研究员,主要从事植物分子遗传学研究,E-mail:zhoushuobio@163.com。
植物体内存在众多的信号通路,其中钙离子介导的细胞生命活动是其重要的调节机制之一[1]。钙调素(calmodulin,简称CaM) 不仅参与信号转导[2],其活化后还能结合多种酶来调节酶活性,形成相应的生理生化反应,从而对植物细胞分裂、伸长、生长、发育和抗逆等活动进行调节[3],并对花粉萌发和花粉管伸长具有促进作用[4]。在一种植物体内,可存在多种CAM亚型基因,且不同亚型基因间的序列具有高度的同源性,但存在的这些差异也使其不同的亚型具有相应的生理功能[5-6]。脱氧核糖核酸或核糖核酸与其编码蛋白之间的关系可以通过密码子来表示[7]。一共有64种遗传密码来编码20种氨基酸[8],这种编码同一个氨基酸的密码子称为同义密码子,在翻译成氨基酸的整个环节中,基因对同义密码子的使用具有不平衡性,还存在物种的差异性等特征[9]。基因密码子偏好性的研究有利于基因水平转移及其家族分化的研究[10]。因此,CaM1基因密码子偏好性的分析能更好地揭示其在系统分类研究中的有效性。
在同一种植物体的不同组织和不同发育阶段,CaM基因的表达存在差异[11]。程玉豆等克隆出鸭梨CaM基因2种亚型(命名为PbCaM1和PbCaM2),发现这2种亚型在成叶、盛花期花瓣及幼果期的表达量较高[12]。郭丽君发现美国山核桃CaM基因在不同组织和发育阶段也不一致,在叶中的表达量最高,其次分别是嫩叶和内茎[13]。赵芊等在研究拟南芥7个CaM亚型基因时发现,不同亚型的表达量均受到细菌信号3-羰基辛酰基高丝氨酸内酯不同程度的诱导,且野生型CaM1基因处理6 h达到表达高峰[5]。薛敏等在非生物胁迫条件下,克隆出沙冬青CaM1基因,通过RT-PCR方法证实其在低温、干旱和盐胁迫下表达水平上调[14]。赵晶等发现灰皮支黑豆和感病品种辽豆15根部在线虫侵染后的第5天,抗感品种的CaM1基因的表达量最高[15]。
对植物CaM1基因在不同条件下的表达量进行深入研究,对揭示钙离子参与的信号通路有重要意义,但目前为止还未见CaM1基因密码子使用模式的报道,这对后期开展异源表达和遗传转化相关试验都是不利的。本研究以43种植物CaM1基因密码子使用特征为切入点进行其进化和分类的研究,通过CodonW分析了43种(12类)植物CaM1基因密码子偏好性,运用Bioedit和MEGA软件对遗传距离和亲缘关系进行分析,以期为CaM1基因在植物中的进化和分类提供理论依据。
1 材料与方法
1.1 试验的时间和地点
本试验于2017年进行,目的基因的搜集在河北科技大学生物科学与工程学院完成,生物信息学分析在河北菲尼斯生物技术有限公司和河北省农林科学院遗传生理研究所完成。
1.2 目的基因的导入
本研究所用基因资料都来源于Nucleotide(https://www.ncbi.nlm.nih.gov/nuccore/)数据库,43种植物CaM1基因含有禾本目、蔷薇目、管状花目、白花菜目、山柑目、伞形目、茄目、豆目、菊目、鼠李目、无患子目和芸香目。43种植物CaM1基因序列都是完整的编码区序列,序列全长为450 bp。如表1所示,利用MEGA 6.0对上述序列进行碱基分析。
1.3 不同密码子使用偏性指标分析
利用CodonW软件计算ENc、GC、GC3s、CBI和相对同义密码子使用度(relative synonymous coon usage,RSCU)的值,分析CaM1基因密码子使用偏性; 并计算GC、GC3s、蛋白质亲水
1.4 遗传距离和系统发育树的构建
利用BioEdit软件对43种植物CaM1基因cDNA序列进行比对,将比对结果保存为fasta格式,通过MEGA软件中Kimura双参数模型计算遗传距离。利用MEGA软件的邻近法建立系统发育树,并通过自举检验法(Bootstrap method=1 000)校正。
2 结果与分析
2.1 不同植物CaM1基因序列碱基特征
如表1所示,编码CaM1基因的43种植物的核苷酸全长均是450 bp。在ATCG碱基分布中,禾本目的小麦、玉米、水稻、二穗短柄草、大麥和甘蔗,蔷薇目的节节麦、小米、花生、芸豆和绿豆,管状花目的芝麻,白花菜目的芜菁和醉蝶花,上述物种的A+T碱基含量均低于G+C碱基含量,其余物种G+C碱基含量高于A+T碱基含量,表明CaM1基因更倾向于使用G或C碱基。
2.2 密碼子使用偏性指标及其影响因素分析
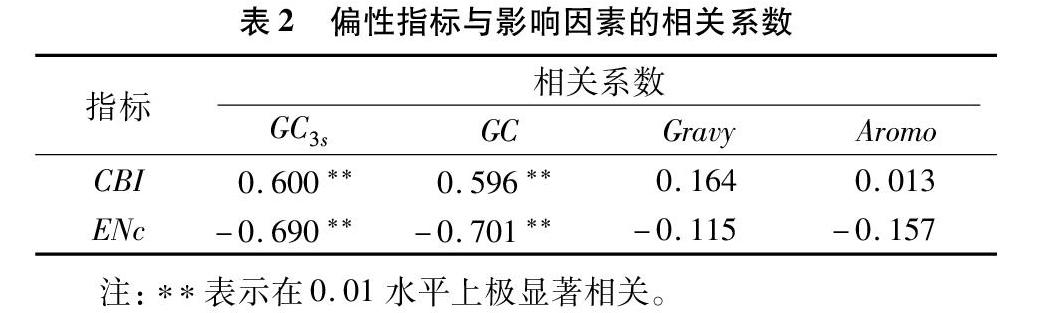
如表1所示,玉米的ENc值最小,为29.15,且禾本目其余植物的ENc值也明显小于其他植物,表明禾本目的植物对密码子使用具有较强的偏好性,43种植物中,只有12种植物的ENc值高于55,ENc值最高的植物为鹰嘴豆和葡萄(ENc=61.00),表明大多数植物CaM1基因对同义密码子的使用具有较强的偏好性。禾本目植物的GC含量值和密码子第3位的GC含量值是所有植物中最高的,表明禾本目植物偏爱使用G、C碱基。在43种植物中,禾本目的玉米、水稻,蔷薇目的小米,白花菜目的醉蝶花,山柑目的拟南芥、亚麻荠的CBI值均大于0.2,禾本目的二穗短柄草、大麦,蔷薇目的节节麦、花生,白花菜目的萝卜、芜菁的CBI值均大于0.1,其余植物的CBI值接近0,表明禾本目、蔷薇目、白花菜目和山柑目的CaM1基因密码子使用偏好性大于其他植物。同义密码子相对使用度的结果表明,43种植物对CUC、AUC、UCU、CCA、ACU、UGA(终止密码子)、CAU和UGC具有极强的偏好性(RSCU>1.5),除上述密码子外,禾本目植物更偏向使用GUC、UCC、UCG、AGC、ACC、UAC、CAG、GAG、CGC、GGC(RSCU>1.5),这与其他11种分类植物密码子的使用特征不一致。如表2所示,GC含量和密码子第3位的GC含量均与密码子偏爱指数呈极显著正相关,但与有效密码子数呈极显著负相关,表明G、C碱基的含量和位置对CaM1基因密码子偏性具有重要影响。
2.3 不同植物CMA1基因的遗传距离分析
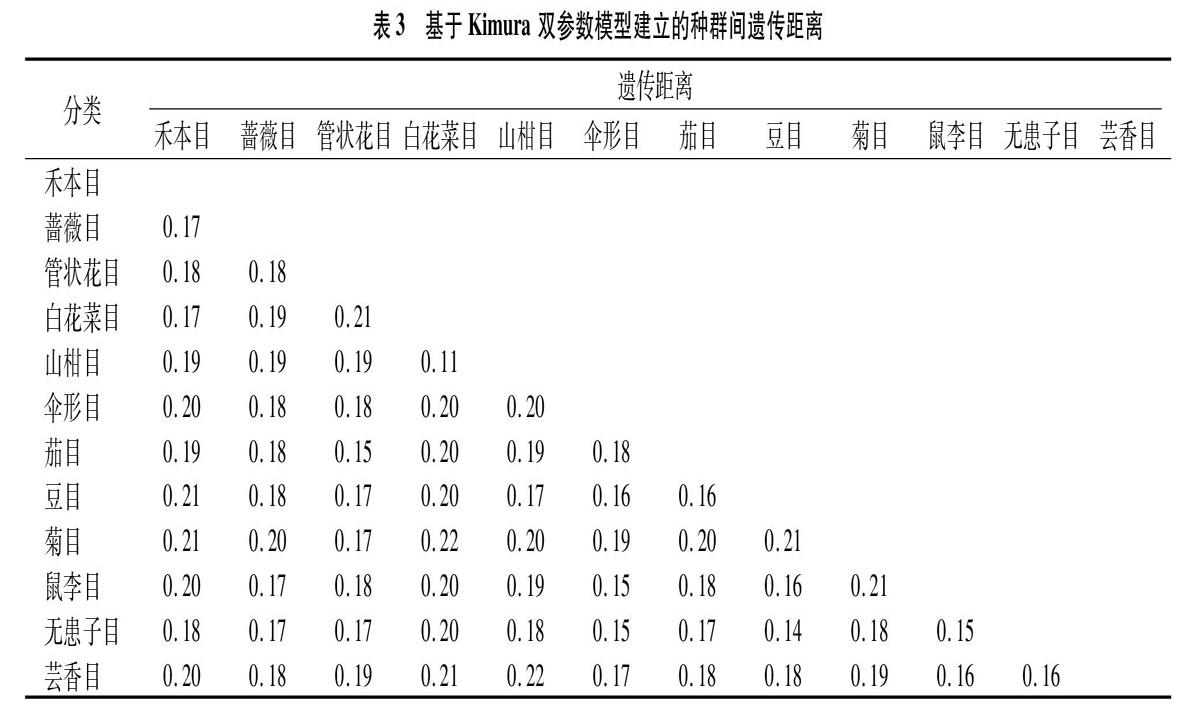
不同植物种群间Kimura双参数距离如表3所示,各分类群体间遗传距离变异区间在0.11~0.22之间波动,其中山柑目和白花菜目之间的遗传距离最近,双参数距离值为011,其次是豆目和无患子目。白花菜目与菊目、山柑目与芸香目之间的遗传距离最远,双参数距离值均为0.22,其次是豆目与菊目、白花菜目与管状花目、白花菜目与芸香目。
2.4 不同植物CMA1基因的聚类分析
如图1所示,43种植物明显分为5类:禾本目(小麦、玉米、二穗短柄草、大麦、甘蔗)和蔷薇目(节节麦、小米、花生、芸豆、绿豆)间的亲缘关系较近;蔷薇目(蒺藜状苜蓿、豌豆、木豆)与豆目(大豆、狭叶羽扇豆、鹰嘴豆)和无患子目(冬青)间的亲缘关系较近;蔷薇目(苹果、白梨),伞形目(人参、西香芹菜),鼠李目(葡萄、枣)和芸香目(克莱门柚)间的亲缘关系较近;管状花目(番茄、马铃薯、矮牵牛)和茄目(簇生椒、烟草)间的亲缘关系较近;白花菜目(小盐芥、萝卜、芜菁、醉蝶花)和山柑目(拟南芥、亚麻荠)间的亲缘关系较近,且同一目下的各个植物间的亲缘关系更近。
3 讨论与结论
密码子偏好性的研究为了解物种起源于进化、目的基因的改造、推测未知基因表达水平及预测目的基因最佳宿主奠定了理论基础[16-17],同时为转基因研究中的甲基化能否引起基因表达量下降或基因沉默提供参考[18]。
本研究通过分析43种不同植物的密码子使用特征,发现其有效密码子数(29.15≤ENc≤61.00)和密码子偏爱指数这2种指标表明不同植物CaM1基因对密码子的使用具有极强的偏好性,通过GC含量、密码子第3位的GC含量和同义密码子相对使用度这3种指标证实不同植物CaM1基因对G、C碱基的使用有极强的偏好性,尤其是对终止密码子UGA,以及CUC、AUC、UCU、CCA、ACU密码子的使用。禾本目与其他分类植物相比,更倾向于使用G/C结尾的碱基,引起其GC3s值远远高于其他植物,且GUC、UCC、UCG、AGC、ACC、UAC、CAG、GAG、CGC、GGC为其最主要的偏爱密码子。分析影响CaM1基因密码子使用偏性的原因发现,G、C碱基的含量和位置对CaM1基因密码子偏性具有重要影响。关于影响密码子偏好性的原因,有研究报道,反转录形成的cDNA序列长度、转运RNA丰度、突变和自然选择等都可造成基因对某些密码子的使用具有极强的偏好性[19]。GC碱基含量会造成同义密码子在使用上具有偏好性的特点,且其偏好性越强,发现该基因以G/C碱基结尾的可能性越大,而且GC3s对该基因密码子偏好性有主导作用[20]。每种生物都有可能面临非自然选择压力,这种非自然选择压力会促使物种的密码子第3位碱基种类发生变化[21]。苏丽艳等发现苹果CaM基因有响应损伤、高温、低氧等非生物胁迫的能力, 在适应胁迫环境初期过程中不可或缺[22]。在体温胁迫下,茶树CaM蛋白也可介导钙信号从而提高适应低温环境的能力[23]。赵存鹏等发现棉花在胁迫处理后,转入CaM基因能提高棉花抵御低温的能力[24]。
基因DNA水平上的分析更有利于基因功能信息的获取,本研究分析12种不同分类植物CaM1基因的遗传距离发现,山柑目与白花菜目之间的遗传距离值最小,其次是豆目和无患子目,这与基于邻近法构建的聚类图一致,表明遗传距离和聚类分析都能描述物种间的亲缘关系。CaM1基因密码子使用特征的确定为研究影响植物抗逆因素及开展抗逆品质改良奠定基础,并为深入建立转基因表达系统提供最优密码子的理论依据。
本研究分析了43种植物CaM1基因密码子使用特征,发现不同类植物CaM1基因对密码子的使用具有较强的偏好性,尤其偏好使用G/C碱基,CUC、AUC、UCU、CCA、ACU和UGA为最优密码子,禾本目植物更偏向使用G/C结尾的碱基。G、C碱基的含量和位置是影响CaM1基因密码子偏性的主要原因。遗传距离分析得出的亲缘关系与系统发育结果一致。
参考文献:
[1]Kudla J,Batistic O,Hashimoto K. Calcium signals:the lead currency of plant information processing[J]. The Plant Cell,2010,22(3):541-563.
[2]Sanders D,Brownlee C,Harper J F. Communicating with calcium[J]. The Plant Cell,1999,11(4):691-706.
[3]张俊红. CaM对增强UV-B辐射小麦幼苗抗氧化系统调控的研究[D]. 临汾:山西师范大学,2010.
[4]Yang X E,Wang S S,Wang M,et al. Arabidopsis thaliana calmodulin-like protein CML24 regulates pollen tube growth by modulating the actin cytoskeleton and controlling the cytosolic Ca2+ concentration[J]. Plant Molecular Biology,2014,86(3):225-236.
[5]赵 芊,张 超,刘 方,等. 钙调素参与细菌信号3-羰基辛酰基高丝氨酸内酯对拟南芥主根生长的调控[J]. 植物生理学报,2016,52(6):933-940.
[6]刘 伟. 小麦与叶锈菌互作过程中CaM的亚细胞定位和CaM亚型的抗体制备[D]. 保定:河北农业大学,2010.
[7]刘庆坡,薛庆中. 遗传密码子及其应用[J]. 中国生物化学与分子生物学报,2006,22(11):851-855.
[8]周 丹,薛仁余,张晓峰,等. 鲤和斑马鱼HOX基因家族同义密码子使用偏性的分析[J]. 水产学杂志,2013,26(2):19-25.
[9]朱孝轩,朱英杰,宋经元,等. 基于全基因组和转录组分析的赤芝密码子使用偏好性比较研究[J]. 药学学报,2014,49(9):1340-1345.
[10]Wei L,He J,Jia X,et al. Analysis of codon usage bias of mitochondrial genome in Bombyx mori and its relation to evolution[J]. BMC Evolutionary Biology,2014,14:262.
[11]刘新颖,王晓杰,薛 杰,等. 小麦钙调素新亚型TaCaM5的克隆及表达分析[J]. 作物学报,2010,36(6):953-960.
[12]程玉豆,葛文雅,闫洪波,等. ‘鸭梨钙调素基因的克隆与表达分析[J]. 河北农业大学学报,2016,39(2):58-63.
[13]郭丽君. 美国山核桃CaM基因的克隆及在盐胁迫下的表达[D]. 南京:南京林业大学,2013.
[14]薛 敏,王雪峰,王志林,等. 沙冬青AmCaM1基因的克隆及其功能初步分析[J]. 植物遗传资源学报,2015,16(6):1315-1320.
[15]赵 晶,王媛媛,朱晓峰,等. 线虫胁迫下大豆钙调蛋白基因的表达分析[J]. 中国油料作物学报,2017,39(1):91-98.
[16]吴宪明,吴松锋,任大明,等. 密码子偏性的分析方法及相关研究进展[J]. 遗传,2007,29(4):420-426.
[17]宗秋芳,戴开宇,刘 颖,等. 猪氨肽酶N(APN)基因密码子偏好性分析[J]. 扬州大学学报(农业与生命科学版),2016,37(2):45-50,73.
[18]张 乐,金龙国,罗 玲,等. 大豆基因组和转录组的核基因密码子使用偏好性分析[J]. 作物学报,2011,37(6):965-974.
[19]Liu G,Wu J,Yang H,et al. Codon usage patterns in Corynebacterium glutamicum:mutational bias,natural selection and amino acid conservation[J]. Comparative and Functional Genomics,2010:343569.
[20]Carlini D B,Chen Y,Stephan W. The relationship between third-codon position nucleotide content,codon bias,mRNA secondary structure and gene expression in the drosophilid alcohol dehydrogenase genes Adh and Adhr[J]. Genetics,2001,159(2):623-633.
[21]時 慧,王 玉,杨路成,等. 茶树抗寒调控转录因子ICE1密码子偏性分析[J]. 园艺学报,2012,39(7):1341-1352.
[22]苏丽艳,田爱梅,陶贵荣,等. 苹果MdCaM的克隆及其对果实釆后非生物胁迫的响应[J]. 华北农学报,2017,32(1):47-52.
[23]Wang X C,Zhao Q Y,Ma C L,et al. Global transcriptome profiles of Camellia sinensis during cold acclimation[J]. BMC Genomics,2013,14:415.
[24]赵存鹏,郭宝生,王凯辉,等. 通过转CaM基因提高了棉花抗寒性[J]. 棉花学报,2016,28(3):234-241.陆秀娟,潘 虹,李祥栋,等. 薏苡种质资源ISSR分子标记筛选及亲缘关系分析[J]. 江苏农业科学,2019,47(5):32-36.
