植物叶功能性状间的权衡研究进展
2019-07-07闫淑君黄柳菁马雯雯王云霄
靳 莎,闫淑君,黄柳菁,陈 莹,马雯雯,王云霄,王 喆
(福建农林大学 园林学院,福建 福州 350002)
植物功能性状(Plant Functional Trait)是植物在漫长进化过程中长期适应外界环境后所呈现出来的能够客观反映植物对外界环境适应性的特征,对植物个体生长、生存和繁殖有重要意义[1],是将植物与环境和生态系统连接起来的桥梁[2]。叶功能性状是植物功能性状的重要组成部分,与植物的生物量和植物对光照、水分、养分的吸收利用及利用程度密切相关,能够反映植物为了最大限度减小环境带来的不利影响而形成的生态适应策略[3~5]。同时,植物叶性状的相对稳定性、对碳收获的重要性以及各因子间相互关系在各植物种群和群落中所具有的相似格局,对于将叶片微尺度上的研究结果扩展到群落、区域乃至全球尺度中有着重要的桥梁作用[6~7]。因此叶功能性状逐渐成为生态学研究的热点之一。
对叶性状的研究最早可追溯到约20世纪70年代,Azizi等[8]研究了不同温度和光照时长对紫花苜蓿(Medicagosativa)叶斑性状的影响。而植物功能性状第一次出现在植物学期刊中大约是20世纪末,其被认为是能够反映植被响应环境变化的核心属性,其中包含了叶性状,而后开始被学者们用于生态学研究[9~11]。其中关于叶功能性状最著名的研究是由澳大利亚科学家Wright等[6]提出的叶经济谱(Leaf Economics Spectrum)的概念,第一次在全球尺度上对植物叶的结构、生理等性状进行定量分析,并探讨各性状之间的关系,此后大量研究陆续涌现。近年来,国内外有关植物叶片性状的研究涉及个体、群落、区域乃至全球尺度,主要集中在通过对大量植物的对比研究,分析叶性状的生态功能,揭示叶性状的分异规律、不同叶性状之间、叶性状与环境因子之间的相关关系[12]。本文旨在总结叶功能性状的生态作用及各功能性状间的权衡关系,探讨当前研究中的不足并对未来研究方向做出展望,以期为今后研究提供参考。
1 叶功能性状的测定指标及其作用
叶片功能性状主要包括叶片的形态、结构和生理等一系列可测量的指标[13]。叶片形态性状一般指叶片表型特征,叶片结构性状是植物叶片的生物化学结构特性,而叶片生理性状主要表现植物的生长代谢情况[13~16](见表1)。从已有研究成果来看,叶寿命、叶面积、叶厚度、比叶面积、叶干物质含量、比叶质量、叶脉性状以及叶片养分含量等指标因为容易测定且能比较好的反映植物的生长状况,常被应用于植物叶功能性状的研究中[2,6,13,16]。
表1叶功能性状常用指标及其作用
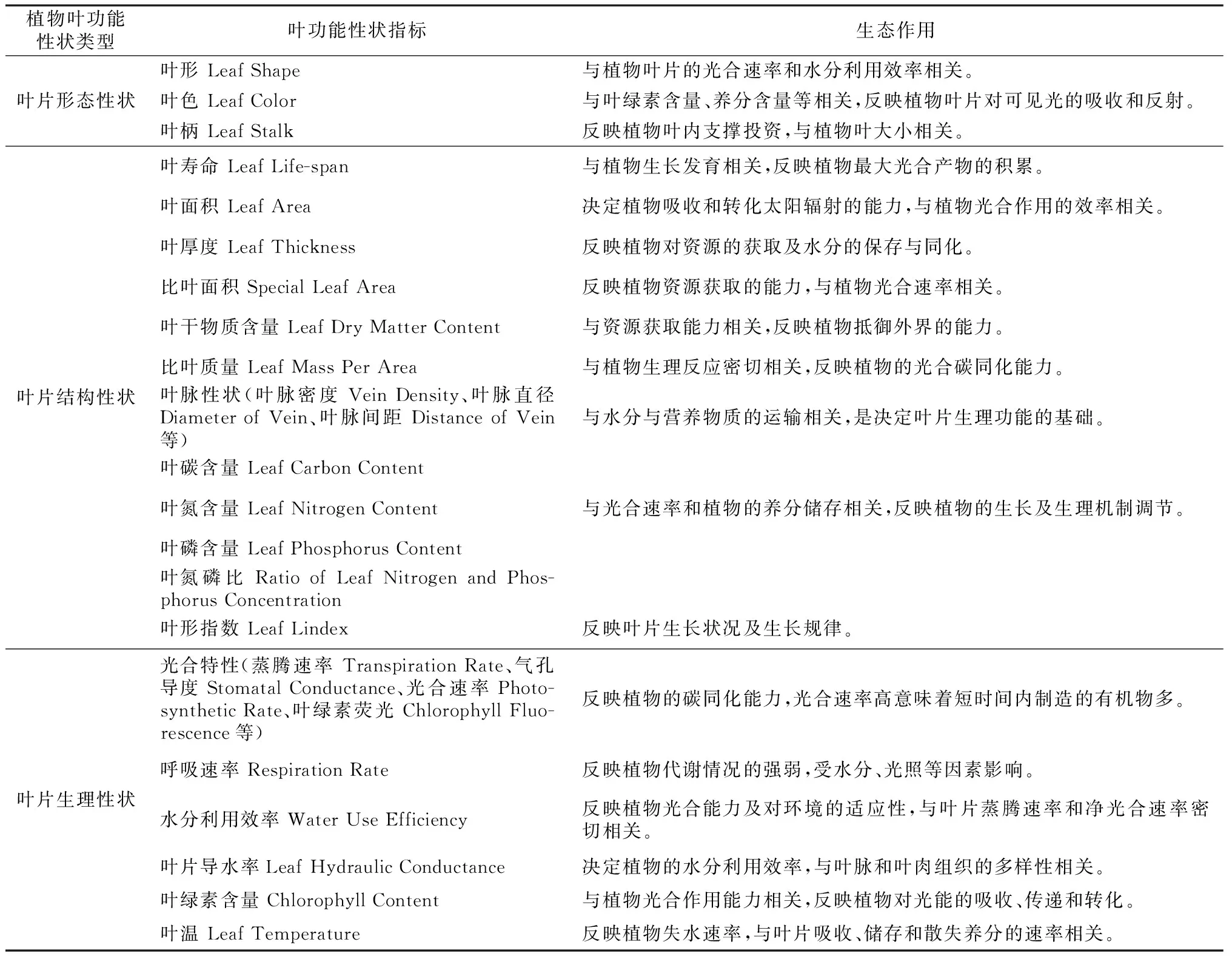
Tab.1 Common indicators of leaf functional traits and their effects
1.1 叶寿命
叶寿命是植物在长期适应自然选择的过程中,为获得最大光合产物而形成的一种综合性指标,能反映植物的生长发育状况和功能[17~18]。
叶寿命的长短直接影响植物干物质的积累,进而影响群落的功能和生产,甚至是生态系统中的物质循环和能量流动[19]。不同植物叶寿命存在差异,而生境改变时同种植物的叶寿命也不尽相同[20]。王希华等[20]研究了浙江天童主要阔叶树种的叶寿命,揭示了不同生活型植物的叶寿命表现为小乔木>灌木>中乔木,而王磊[21]对黄土高原常见落叶木本植物的研究却表明,灌木寿命通常比乔木更长,总体来说符合前人研究中光照强度与叶寿命负相关的研究结果[22],但也有一定差异,造成这种差异的原因可能与植物的选择有关,前者研究对象为常绿阔叶树种,而后者为落叶植物,其叶片更新的周期有差异。同时,叶寿命会受到环境因素的影响,如青海云杉(Piceaasperata)叶寿命在不同海拔之间存在极显著差异性,并且除在海拔 2 550 m处之外,其寿命均为随海拔增加而增加,这种变异是一种非遗传的气候适应特征[23];Hemminga等[24]研究表明海草叶寿命从热带到温带逐渐增加,因为气温越低的地方水分有效性越差,植物叶寿命越长[23]。
1.2 叶面积
叶片是植物固碳的主要部位,叶片大小直接影响着植物对光的截取和对碳的获取能力,与植物的光合作用和蒸腾作用密切相关[25~26]。
叶面积的大小受叶柄限制,通常叶柄干重相同时,常绿树种的叶片小于落叶树种[26],而在给定植物叶面积的情况下,常绿物种比落叶物种需要更高的叶柄生物量投资[27]。同时,叶面积的大小与叶片干重相关,如祝介东等[26]研究表明在相同叶片干重下,灌木较乔木、落叶植物较常绿植物的叶面积更大。此外,气候和地形因子也是引起植物叶面积变化的原因。许多大尺度的研究表明叶片面积与年均温、年降水量呈正相关[28~30];Lebrija-Trejos等[31]认为随海拔升高,大部分植物叶片面积会减小,但也有增大或先增大后减小的现象。通常海拔升高会导致气温降低,而低温使叶的生长季相对较短,细胞生长缓慢,因此叶片会普遍变小;但温度不是决定叶片性状的唯一限制因子,海拔的跨度在叶性状的构成中也起着关键作用,如辽东栎(Quercuswutaishanica)的叶面积在海拔约 1 400 m以下时会随着海拔的升高而增大,海拔高于 1 400 m时随海拔升高而减小,而在海拔约 1 400 m处为最大值[32],可见只有海拔到达一定高度时温度才能成为植物生长的限制因子。同时,在高温干旱条件下,植物也会通过缩小叶面积的方式来降低叶温以适应干旱环境[33],因为单位叶面积减小可以增加叶片保水性,减缓叶温升高的速率,从而提高植物的水分利用效率[34]。
1.3 叶厚度
叶厚度与植物资源获取、水分保存及同化有关。叶片厚度的增加,能降低植物内部水分散失,提高植物保水率;而叶片厚度减少能加强植物在微弱光线下的光合作用能力[35~37]。
研究表明厚叶的单位叶面积叶绿素含量与氮含量相对较高,有利于植物对光的吸收和转化[38~39];雷蕾等[40]研究发现生长在高密度湿地和低密度湿地的黄帚橐吾(Ligulariavirgaurea),自身分别采取了叶绿素含量高的厚叶片和叶绿素含量少的薄叶片两种适应模式;而在土壤水分饱和时,芦苇(Phragmitesaustralis)叶片较薄,反之,叶片较厚[41]。以上研究反映了植物对光照、水分等异质性环境因子的资源利用策略。
1.4 比叶面积、叶干物质含量、比叶质量
比叶面积能反映植物的碳收获策略[42],叶干物质含量与植物抵御外界的能力呈正相关,两种指标都能指示植物利用资源的能力,是植物适应外界环境变化的关键性状[43]。
比叶面积受光照影响较大,Meziane & Shipley[44]研究表明通常光线越弱的地方形成的叶子比叶面积越大,覃鑫浩[45]的研究结果与其一致。此外,比叶面积还与演替、海拔等因素相关,如胡耀升等[46]研究发现长白山森林植被比叶面积随演替的进行呈现明显增加的趋势,并且比叶面积与海拔、坡位和土壤氮含量均呈显著正相关。余华等[47]认为天然刨花楠(Machiluspauhoi)林下幼苗叶干物质含量会随着经度的增加而增加,而李宏伟[48]则发现黄土高原森林群落主要物种的叶干物质含量主要受坡向影响。
比叶质量与比叶面积互为倒数,目前研究表明各生活型植物及植物在不同生长时期的比叶质量均存在差异[49~51],并且其大小与遗传物质和环境因子相关[52~54]。
1.5 叶脉性状
叶脉在植物生长过程中承担了水分与营养物质的运输,是决定叶片生理功能的基础[55]。目前研究中大多用叶脉密度、叶脉直径、叶脉间距及叶脉闭合度等一系列性状指标来表征叶脉性状特点[56]。
关于叶脉的研究主要是将其与植物水分利用及光合作用相结合,探究对植物生理功能的影响和对环境的响应。相关研究表明叶脉密度与叶片的光合速率、水分运输与传导等紧密相关,体现了叶片蒸腾成本与光合收益之间的权衡[57~58]。韩玲等[59]发现芨芨草(Achnatherumsplendens)在土壤水分充足和水分胁迫两种条件下,分别选择少量粗叶脉和大量细叶脉的构建模式,体现了其在资源异质性分布的生境中根据自身需求对资源进行优化配置。谢兆森等[60]研究显示葡萄(Vitisvinifera)叶片末端叶脉会随着葡萄叶片的生长而生长,从而使叶脉密度逐渐增加,提高树体的水分利用效率,表明植物在不同生长阶段对资源需求程度具有差异。而在干旱或半干旱区,年平均降雨量与叶脉密度呈显著负相关[61],体现了植物在干旱环境下的适应策略。同时,游文娟等[62]解剖了不同光强下14种绿化植物的叶片,发现叶脉系统在全光照下比较发达,而在遮阴条件下发育较差,体现了植物在不同光环境下对自身结构的调节。
1.6 叶片养分含量
植物叶片中碳、氮、磷元素的化学计量特征对于解读陆地生态系统空间格局变化规律、未来变化趋势的预测和全球变化的响应具有重要作用[63],因此近年来被频繁应用于植物叶功能性状的研究中。三种元素均对植物的生长及生理机制调节有重要作用,而三种元素两两之间的比值往往能反映植物的生态适应策略[64]。
植物在不同生长时期养分含量不同,且各生活型之间养分含量也有差异。如培植于温室大棚内的铁皮石斛(Dendrobiumofficinale)叶片碳含量在3月最高,随后逐渐降低,8月以后则保持相对稳定,氮含量分别在1月和12月最高,而磷含量则在8月、12月最高[65];林恬等[66]发现福建万木林12种优势植物中,叶片碳、氮含量均表现为春夏高于秋冬,且各生活型间叶碳含量大小顺序为乔木层>灌木层>草本层,氮含量为灌木层>乔木层>草本层。同时,叶片养分含量会影响植物的生理功能,相关研究表明叶氮含量是决定光合能力强弱的重要指标[67],而苗艳明等分析了不同功能型植物叶氮含量与光合特性的关系,指出单位质量植物叶片氮含量与光合速率显著正相关[68]。
植物叶片养分含量通常与其生长环境及遗传特性相关。王利平等[69]认为在低盖度条件下芦苇叶片全碳、全氮、全磷含量最高,方怡然等[70]发现不同阔叶树种叶片养分浓度差异明显,且造成不同树种叶片氮、磷含量差异的主要原因是植物遗传特性。
叶片氮磷比作为描述群落水平上植被结构、功能和养分限制的重要指标[64],在不同地域有所差异。Reich & Oleksyn[71]分析了全球范围内 5 078份植物样品氮和磷的化学计量关系,发现氮磷比随纬度的上升呈减小趋势,因为一般热带土表现为磷缺乏,而温带土表现为氮缺乏[72],使植物可吸收的养分受限。但国内研究者统计了我国52个采样区内649个湿地植物不同器官和全株样本氮、磷含量数据,表明氮磷比的几何平均值在热带地区最大,温带次之,亚热带最小,产生差异的原因可能与湿地复杂的生态系统有关[73]。同时,植物叶片或生物量中的氮磷比也可作为判断环境对植物生长的养分供应情况以及植物生长速率的重要指标[74]。研究表明植物生长受氮磷比限制的阈值为14和16,当植物叶片氮磷比高于16时,植物生长主要受磷限制;当植物叶片氮磷比低于14时,植物生长主要受氮限制;处于两者之间时表示受两者共同限制[75]。而学者们在研究养分供应与维管植物生长速率关系时发现,桦树幼苗氮磷比在磷限制下与生长速率负相关,在氮限制下与生长速率正相关[76]。
2 叶功能性状间的权衡关系
2.1 叶片形态性状之间的权衡
对所有生长的植物而言,植物生长过程中对环境的适应不是通过单一性状的改变来完成的,而是多种性状共同协调,所以植物叶功能性状之间总是存在某种关联。
有学者在已建立的木本植物叶片形态、叶面积等与降雨量有关的模型研究中,表明叶片形态可应用于植物适应能力大小的判断[77]。郭琪等[78]通过对山西刺槐(Robiniapseudoacacia)种质资源叶片表型多样性的分析,指出复叶叶柄长度与其叶长和叶宽均呈极显著正相关关系。杨冬梅等[79]发现叶大小与叶数量之间存在某种权衡,资源的限制通常使植物很难同时增大叶大小与叶数量,在给定某一个叶片生物量时,植物可能会有较多的小叶片或较少的大叶片。
2.2 叶片形态性状与生理性状之间的权衡
植物叶片形态的改变,能在一定程度上能引起其生理性状的改变。如张玉洁等[80]以海岛棉(Gossypiumbarbadense)叶片为材料,对叶片主脉两侧堆成部分的光照日辐射、净光合速率、叶绿素含量以及叶面积进行测试比较,结果表明海岛棉叶片卷翘引起不同方位叶片主脉两侧对称部分所截获的光能异质性,导致叶面积和叶绿素含量不同,进而共同导致了光合能力的差异,由此可见叶片形状与叶片光合作用之间存在某种关联;李永华等[81]认为叶片形态变化与叶片表面温度之间存在某种联系,在干旱区植物叶片变小能有利于降低植物叶片表面温度,而在高温低风速情况下叶片宽度减小也会使叶片温度降低,体现了植物对特殊生境的适应模式。
2.3 叶片结构性状之间的权衡
全球叶片经济型谱的建立明确了比叶质量与叶片单位干物质的最大光合能力、叶龄以及单位干物质的氮、磷含量等密切相关。比叶质量低的植物,其叶寿命较短、单位干物质营养元素含量较高,光合能力强、单位叶面积的经济投入较低,使植物资源的利用效率高而生长速度快[82~83]。Arredondo & Schnyder[84]认为禾本科植物的比叶面积与叶干物质含量呈负相关,这与张晶等[85]对科尔沁沙地典型草地植物的研究结果一致。洪陈洁等[86]研究了不同品系福建山樱花(Cerasuscampanulata)叶功能性状,发现比叶面积与叶干物质含量存在极显著负相关关系,而与叶绿素含量呈极显著正相关。余华等[47]指出天然刨花楠林下幼苗叶比叶面积与叶组织密度、叶面积均呈显著负相关,而叶组织密度与叶干物质含量呈显著正相关。Wright等[6]通过对175个采样点2548种植物叶性状的研究,发现叶片的比叶面积、光合速率、暗呼吸速率、氮和磷含量彼此间呈正相关,而Reich等[87]认为叶寿命与比叶面积、光合速率、以及氮和磷含量均显著负相关。
叶脉作为影响植物水分供应和利用的重要结构,与其他叶性状之间关系紧密。韩玲等[88]研究发现张掖湿地芨芨草叶脉密度和叶脉直径在冠盖区、过渡区及空旷区分别呈现不同程度的负相关关系,而不同土壤水分条件下芨芨草叶片厚度与叶脉密度显著正相关,与叶脉直径显著负相关[59]。徐婷等[89]的研究揭示了不同坡向刺槐一级叶脉密度和二级叶脉密度与叶面积分别呈不同程度的负相关性。而龚容等[61]认为叶脉密度与比叶质量具有显著正相关关系。
在养分含量方面,已有研究结果显示在大多数植物种群和群落中,植物叶片的氮含量都会随比叶面积的增加而增加[87,90]。张继光等[91]研究了南亚热带16种木本植物,认为成熟叶片的氮和磷含量之间存在极显著正相关关系,氮磷比与氮有弱的正相关关系,与磷呈显著负相关。彭阿辉[92]研究了贡嘎山5种常绿树种的叶功能性状,发现叶片的碳含量会随比叶面积的增加而降低,且叶片碳含量与氮、磷含量均呈显著负相关,而张晶等[85]对科尔沁沙地长期封育草地19种植物的研究表明,叶片碳含量与氮含量呈极显著正相关。其中贡嘎山位于青藏高原东南部,地处中国东部湿润季风区与青藏高原寒冷气候的过渡带,而科尔沁沙地位于内蒙古东部,地处我国温带半干旱草原区,两地在海拔、降水、气温以及土壤类别等方面差异较大,再加上两地植被类型不同,因此可能造成叶功能性状上的差异性,也侧面反映了植物对环境的适应策略
2.4 叶片生理性状与结构性状之间的权衡
国外学者通过对不同生态系统的大量植物叶片性状研究结果综合分析,表明光合速率、暗呼吸速率、叶片的氮和磷含量彼此之间成正相关性,而叶寿命与光合速率、叶片氮和磷含量呈显著的负相关关系[6,87,93]。徐婷等[94]认为植物叶脉密度与水分利用效率的关联性影响着植物水分的供需平衡。韩玲等[88]发现芨芨草的水分利用效率、光合有效辐射、蒸腾速率、净光合速率均与叶脉密度和叶脉直径呈显著的相关性。
3 问题与展望
植物叶功能性状的研究已经取得了比较多的成果,使得学者们重新审视植物应对环境变化的适应机制。目前关于叶功能性状的研究已涵盖个体尺度、群落尺度和全球尺度,但主要集中在自然生境下,且对叶片结构性状的研究偏多,对全球尺度上的研究较少。虽然目前的研究已在一定程度上推动了生态学的发展,但仍存在一些问题有待进一步深入研究:
(1)研究对象 鉴于植物叶功能性状的表达是植物长期适应环境变化的结果,目前关于叶功能性状的研究主要是针对成年树木,虽然张曦等[95]通过盆栽试验探索了不同生育时期紫花苜蓿叶性状对不同程度干旱的响应,余华等[47]分析了不同种源刨花楠林下幼苗对地理环境变化的适应机制,但总体来说对幼苗的研究相对较少。通过对成年树木叶片性状指标的测定可以看出植物在适应环境的过程中哪一些性状发生改变,但无法估计叶片性状从哪一阶段发生变化及变化的过程,因此对于幼苗或自幼苗生长至成年树木阶段叶功能性状的研究或许可以充实前人的相关研究结果。
(2)研究区域 国内关于叶功能性状的研究大多针对野外植物,少有涉及对人工林或城市绿化植物的研究。虽有朱济友等[96]研究表明在城市热环境中叶性状关系与全球尺度基本一致,但个别研究不足以概括整体情况,且南北方城市环境存在差异,因此对于城市环境中植物叶功能性状的研究还有待深入。
(3)性状选择 在叶功能性状指标的选择上,大多数研究偏向于选择容易测定的叶片形态和结构性状,对生理性状和解剖形态等研究较少,导致某些研究结果相悖而无法解释其原因,由此可见关于叶片生理性状功能和解剖形态特征的研究亟待补充以弥补当前的不足。
