高羊茅硅结合蛋白的初步提取及鉴定
2019-06-18新布任王涛马燕妮宋涵王明明
新布任 王涛 马燕妮 宋涵 王明明

摘 要:选取禾本科植物高羊茅的叶片,采用低温捣碎分级分离方法,分离出天然硅体,进一步分离硅体和硅质化细胞壁,去除硅质化细胞壁组分中的杂质蛋白,并利用硅体的专一性染色法进行验证后,再进一步提取出粗的硅结合蛋白,通过SDS-PAGE鉴定出大小为117kDa的硅结合蛋白,最终通过切胶回收透析的方法获取纯净的硅结合蛋白,得出了一套有效的硅体分离提取的方法。该研究首次从高羊茅叶片中分离提取硅体,为高羊茅的抗逆性研究及快速生长研究提供了蛋白获取基础。
关键词:硅结合蛋白;高羊茅;低温捣碎分级分离法;SDS-PAGE
中图分类号 S143 文献标识码 A 文章编号 1007-7731(2019)09-0011-3
硅在自然界中分布广泛,岩石、水流及动植物体内等均有其存在,主要以二氧化硅和硅酸盐等形式存在,陆地生物系统和海洋生物系统大部分都存在硅的生物循环[1]。研究表明,硅是在植物生长的过程中,即使过量也不会对其产生危害作用的有益元素。如今硅肥已经是世界各地普遍使用的无任何毒副作用的新型肥料,是具有增产和抗逆的作用的天然农药[2],硅的主动吸收机制在很多植物内都存在[3],参与生物体很多的新陈代谢过程,其在动物以及单细胞生物(如硅藻)中的必需性已有研究证明[4]。硅对于植物生长有益,硅元素的缺乏会导致非典型环境胁迫[5]。
硅以硅结合蛋白的形式存在于植物体,史新慧等研究表明,禾本科植物中含有大量的硅结合蛋白,并分离出水稻SBP117硅结合蛋白[6]。硅结合蛋白可以提高植物的光合作用和根系活性,增强植物的抗倒伏性、抗病能力、抗逆能力[7-9],降低植物的蒸腾作用,提高产量和品质,促进养分的吸收。细胞壁、硅化细胞和胞间隙或角质层是硅在植物体内的主要沉积位置[10]。Yoshida等[11]研究表明,在水稻叶片中,硅元素含量最高的部位是叶尖,硅元素主要的沉积部位为叶片的上下表皮、维管束鞘以及相连的厚壁细胞。
高羊茅(Festuycaelata)属禾本科、羊茅属多年生草本植物,因其耐热,耐践踏,草坪成型快,颜色浓绿,抗逆性强,管理方便以及成本低等优点,一直是足球场、高尔夫场、公园等的绿化优选草种。刘慧霞等[12]研究表明,在高盐环境下,硅能提高高羊茅的适应能力[13]。宋锐研究表明,在高盐的胁迫下,硅通过直接参与高羊茅的生理生化过程,提高了其在高盐环境下幼苗的适应能力[14]。本研究利用低温捣碎提取法对在高盐环境中生存的高羊茅进行了硅结合蛋白的提取并鉴定,为高羊茅的抗逆性及快速生长研究提供蛋白获取基础。
1 材料和方法
1.1 实验材料 大小适中、均匀一致的高羊茅种子。
1.2 实验方法
1.2.1 种子发芽试验 将购买的高羊茅种子装入纸袋放置暗处室温下贮藏。开始实验时,选择成熟、饱满、大小适中且一致的种子,先进行消毒,用蒸馏水反复冲洗,将种子表面水分用滤纸吸干后,整齐地放在铺有滤纸的培养皿中,每皿放50粒种子,选择不同浓度外源硅离子(硅酸钾+氯化钾)来培育。外源硅离子的浓度分别为:2mmol/L的硅酸钾溶液(K2SiO3)和4mmol/L的氯化钾溶液(KCl)的Si+组,以及加入4mmol/L的硅酸钾溶液(K2SiO3)和8mmol/L的氯化钾溶液(KCl)的Si++组。当K2SiO3溶于水后,加入的Cl-后会阻止弱酸根离子生成H2SiO3,使得实验所需Si2+误差变小。KCL中的K+是植物生长的营养元素,文献报道低浓度的Cl-个对植物基本上没有影响。每组进行3个重复实验组,放在光照培养箱内(20℃,16/8h,15000l)培养,至种子萌发长出四叶后备用。
1.2.2 天然硅体的分离 采用低温捣碎分级分离法从发芽好的高羊茅中分离天然硅体,具体步骤如下:
1.2.2.1 分离出硅体和硅质化细胞壁 取高羊茅叶片剪碎,放于-20℃冰箱,冷冻过夜后取出放于室溫,融化后与1%TrionX-100按照体积比1∶3进行混合,之后用JJ-2型组织捣碎匀浆机捣碎、1mm筛网过滤,得到滤液。静置,自然沉降3min直至沉降物不再增多,弃掉上清。用0.1%SDS溶液重悬沉降物,静置弃上清,反复至上清液透明为止,去除大部分非硅质化细胞壁碎片及细胞质物质,获得粗制硅体及硅质化细胞壁。
1.2.2.2 去除硅质化细胞壁组分中的杂质蛋白 将以上得到浅绿色沉降物,参照史新慧[15]的方法,将沉降物与2%SDS漂洗液按照体积比1∶4进行混合,放于90℃1h,期间间歇式混匀后水洗,重复以上操作。最后将上清弃掉,保留沉降物,再用丙酮进行漂洗,将沉降物与丙酮按照体积比1∶2进行混合在室温下搅拌直至无黄色物质析出。最后用高盐溶液漂洗,将沉降物与6mol/LNaCl按照体积比1∶4进行混合,室温下搅拌30min后,弃上清并水洗,留沉降物。
1.2.2.3 获得纯粹的硅体 将以上得到的沉降物用40um的筛网过滤,除去大的硅质化细胞壁碎片,获得纯粹的硅体。
1.2.3 硅体的专一性染色法 参照Dyanaakan等[16]的染色方法,依次进行脱色、酸处理、脱水、染色,最后在光学显微镜下进行观察硅体形态。
1.2.4 提取硅结合蛋白 参照Harrison[17]的方法,先用NH4F进行处理,按每1g湿硅质化细胞壁加入4mL 10mol/LNH4F在数显恒温磁力搅拌器搅拌,直至在显微镜观察硅体溶解完全,再用HF处理将收集的细胞壁与8%HF溶液同体积混合,搅拌3h,收集滤液,将滤液进行透析,使用冷冻干燥处理浓缩,收集干燥物即为硅结合蛋白。
1.2.5 通过SDS-PAGE电泳鉴定硅结合蛋白的大小 参照Hermannn和Gebhard[18]的方法进行跑电泳,跑完电泳后将凝胶浸泡于考马斯亮蓝染色液中缓慢摇动30~60min进行染色处理,最后将凝胶浸泡于洗脱液中缓慢摇动1~2h,其间更换洗脱液1~2次,洗脱后凝胶照像。
1.2.6 纯化硅结合蛋白 根据Mashairo S等[19]的方法,用透析袋电洗脱法洗脱纯化目的蛋白,将所需目的条带从SDS-PAGE凝胶切下,放入透析袋后进行电泳回收,将透析袋放进盛有预冷的Tris甘氨酸电泳缓冲液直至考马斯亮蓝完全从凝胶条带上跑出,再进行4℃透析,最后用冷丙酮沉淀法处理透析袋内的液体进行蛋白沉淀。采用Brandford法,测定纯化的蛋白质含量。
2 结果与分析
2.1 不同硅离子浓度对高羊茅种子发芽生长的影响 本次研究表明,Si+组比Si++组高羊茅叶片长势不同,Si+组高羊茅叶片丰富,生长密度高,平均长度为7~8cm,同一周期内生长比较快,而Si++组高羊茅叶片比较稀疏,生长密度较Si+组低,平均长度为6~7cm,同一周期内生长比较缓慢。
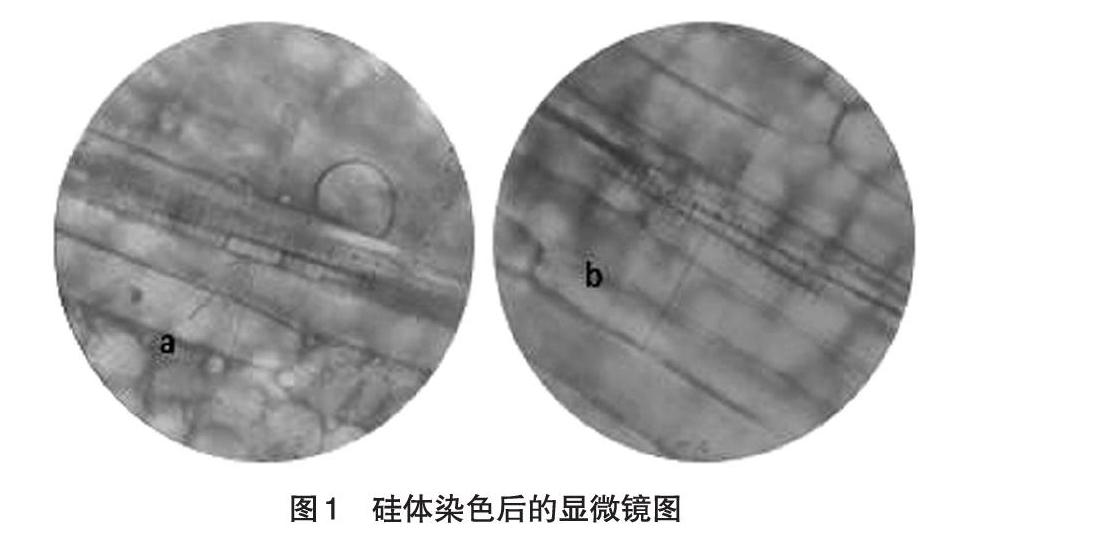
2.2 不同硅离子浓度对细胞形态和数量的影响 如图1所示,a箭头所指为在光学显微镜下油镜(10×100倍镜)观察到Si+组的硅细胞,硅细胞形态短而粗,在叶表皮细胞之间的夹缝产生,不同视野硅细胞数量较少。b箭头所指为在光学显微镜下油镜(10×100倍镜)观察到Si++组的硅细胞,硅细胞形态长而细,在叶表皮细胞之间的夹缝产生,不同视野硅细胞数量较多。
2.3 硅结合蛋白分子的大小 由图2可知:泳道1:Marker:泳道2~4:从加入了2mmol/L硅酸钾溶液(K2SiO3)和4mmol/L氯化钾溶液(KCl)的Si+组发芽的高羊茅提取的硅结合蛋白,泳道5~7:从加入了4mmol/L硅酸钾溶液(K2SiO3)和8mmol/L氯化钾溶液(KCl)的Si++组发芽的高羊茅提取的硅结合蛋白。通过SDS-PAGE鉴定,硅结合蛋白的大小为117kDa。
3 讨论
提取的硅体在显微镜下观察,Si++组较Si+组的的硅细胞形态长而细,不同视野硅细胞数量较多,且从SDS-PAGE电泳的结果图来看,Si++组的高羊茅提取的硅结合蛋白含量较高,并鉴定出了硅结合蛋白的大小。硅促进植物生长主要通过3种方式,即影响叶片光合作用、减少倒伏和增强根系活性等。在维管组织和表皮细胞中硅的沉积增强组织的机械性能,从而增大了叶片受光面积、提升了群体光合效率。硅细胞长而细的形态可能更有利于植物进行光合作用,增强抗逆性。在植物体内硅元素主要存在于细胞壁、硅化细胞和胞间隙或角质层。在高羊茅草的叶鞘中,硅体主要存在于硅化细胞、泡状细胞、硅质化表皮细胞以及硅质化表皮毛等。大部分硅质化表皮细胞形态为长形。
植物体中,硅的存在形态为水化无定形二氧化硅(SiO2·nH20)、二氧化硅(SiO2)、硅酸和胶状硅酸,其中水化无定形二氧化硅(SiO2·nH20)、二氧化硅(SiO2)为主要存在形态[21]。不同植物对硅的吸收能力不同,与植物本身的基因型和环境有关,在pH小于9的条件下,硅以单硅酸Si(OH)4的形式被植物吸收。目前认为高等植物由于硅与不同植物种类的关系不同,硅的吸收形式有主动吸收、被动吸收和拒绝吸收3种[22]。硅对于高羊茅的抗逆性及生长促进作用机制有待今后作进一步的研究。
参考文献
[1]刘鸣达,张玉龙,陈温福.土壤供硅能力评价方法研究的历史回顾与展望[J].土壤,2006(1):11-16.
[2]MaJ.F.,Y.Miyake,E.Takahashi.Silicon as a beneficial element for crop plants.In:Datonoff L.,G Korndoref,G.Synder,eds.Silicon in Agriculutre[J].New York:Elsevier Science,2001:17-39.
[3]TamaiK.,J.F.Ma.CharaeteriZation of silicon uptake by rice roots[J].New Phytol.,2003,158:431-436
[4]wernerD.,R.Roth.Silica metabolism[J].In:Inorganic Plnat Nutrition.New York:New Series,1983.
[5]EPsteinE.The anomaly silicon in Plant biology[J].Proc,Natl.Acad.Sci.USA,1994,91:11-17.
[6]史新慧,覃闯登,宋建兰.水稻及其他禾本科植物体内硅结合蛋白的免疫印迹检测[J].生物化学与生物物理进展,2005,32(4):371-376.
[7]EPsteinE.Silicon.Annu.Rev.Plant Physiol[J].Plant Mol. Biol.,1999,
50:641-664.
[8]NeumnanD.,U.Zurnieden.Silicon and heavy metal tolernace of higher Plnats[J].Phytochemistry,2001,56:685-692.
[9]Linag.YC.,J.W.C.Wong,L.Wei.Silicon-mediated enhancement of cadmium tolerance in maize(ZeamyasL.)grown in cadmium contmainated soil[J].Chemosphere,2005,58(4):475-484.
[10]李文彬.水稻體内硅的生理功能及沉积机理的研究[D].北京:中国农业大学,2004.
[11]Yoshida S.,Y.Ohnishi,K.Kigatishi.Histochmseltry of silicon in rice Plant.II.Localization of silicon within rice tissues[J].Soil Sci.Plant Nutr.,1962,8(l):36-41.
